
Impact of Iron Mining Activity on the Endophytic Fungal Community of Aspilia grazielae

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Plant Collection and Soil Samples
2.3. Pre-Treatment of the Plant Samples
2.4. Extraction of Genomic DNA, PCR Amplification and ITS rDNA Sequencing
2.5. Statistical Analysis
3. Results
3.1. Soil Analysis
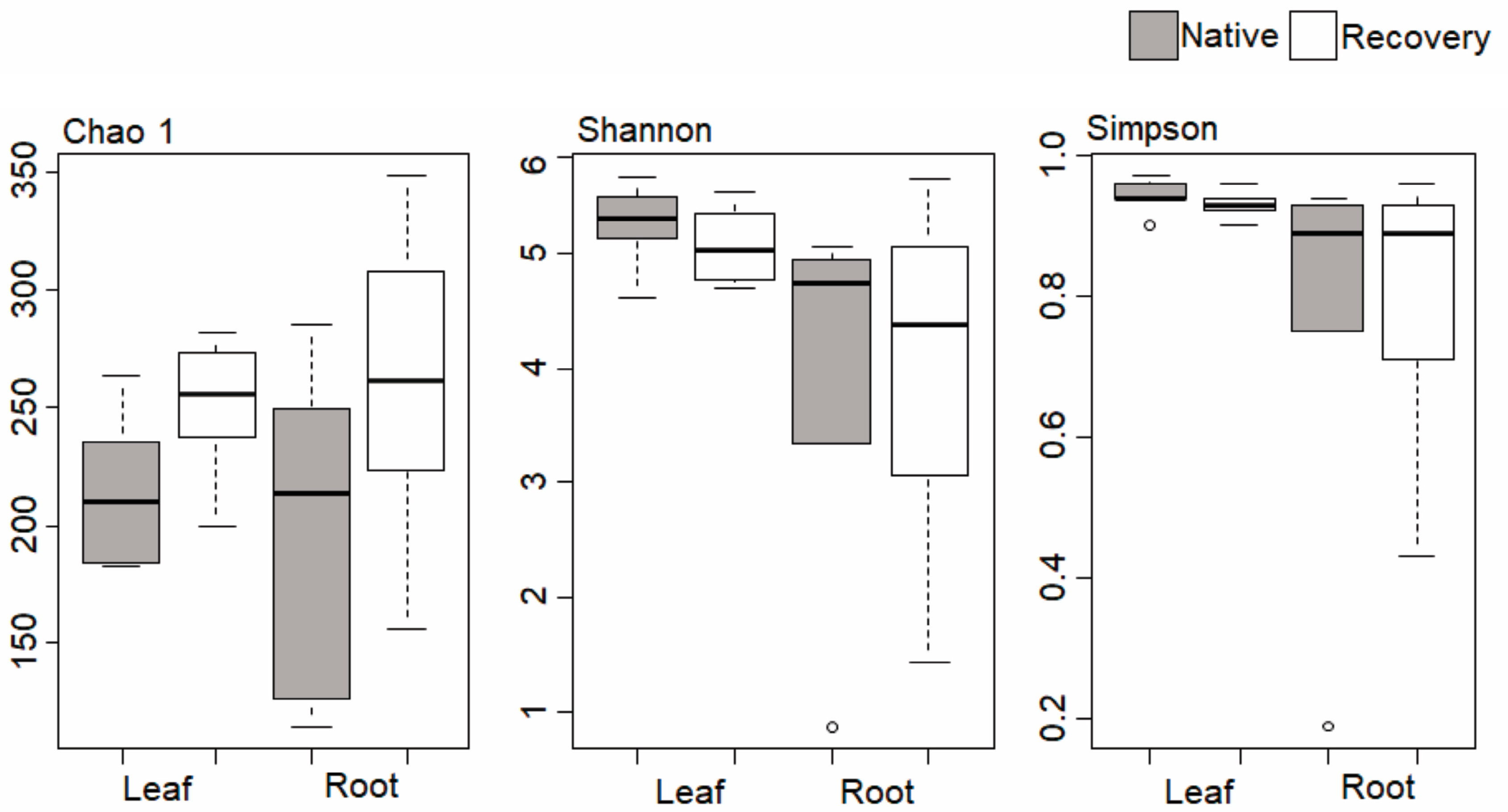
3.2. Community Diversity
4. Discussion
4.1. Revealing the Diversity of Endophytic Fungi in Different Plant Tissues
4.2. Diversity of Endophytic Fungi in A. grazielae Considering Land Use, Soil and Vegetation
5. Limitations, Implications and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marengo, J.A.; Cunha, A.P.; Cuartas, L.A.; Deusdará Leal, K.R.; Broedel, E.; Seluchi, M.E.; Michelin, C.M.; De Praga Baião, C.F.; Ângulo, E.C.; Almeida, E.K.; et al. Extreme drought in the Brazilian Pantanal in 2019–2020: Characterization, causes, and impacts. Front. Water 2021, 3, 639204. [Google Scholar] [CrossRef]
- Tomas, W.M.; de Oliveira Roque, F.; Morato, R.G.; Medici, P.E.; Chiaravalloti, R.M.; Tortato, F.R.; Lourival, R.F.F. Sustainability Agenda for the Pantanal Wetland: Perspectives on a Collaborative Interface for Science, Policy, and Decision-Making. Trop. Conserv. Sci. 2019, 12, 194008291987263. [Google Scholar] [CrossRef]
- Roque, F.O.; Ochoa-Quintero, J.; Ribeiro, D.B.; Sugai, L.S.M.; Costa-Pereira, R.; Lourival, R.; Bino, G. Upland habitat loss as a threat to Pantanal wetlands. Conserv. Biol. 2016, 30, 1131–1134. [Google Scholar] [CrossRef] [PubMed]
- Libonati, R.; Dacamara, C.; Peres, L.; Sander de Carvalho, L.; Garcia, L. Rescue Brazil’s burning Pantanal wetlands. Nature 2020, 588, 217–219. [Google Scholar] [CrossRef]
- Noriler, A.S.; Savi, D.C.; Aluizio, R.; Palácio-Cortez, A.M.; Possiede, Y.M.; Glienke, C. Bioprospecting and structure of fungal endophyte communities found in the Brazilian biomes, Pantanal and Cerrado. Front. Microbiol. 2018, 9, 1526. [Google Scholar] [CrossRef]
- Assad, B.M.; Savi, D.C.; Biscaia, S.M.; Mayrhofer, B.F.; Iantas, J.; Mews, M.; de Oliveira, J.C.; Trindade, E.S.; Glienke, C. Endophytic actinobacteria of Hymenachne amplexicaulis from the Brazilian Pantanal wetland produce compounds with antibacterial and antitumor activities. Microbiol. Res. 2021, 248, 126768. [Google Scholar] [CrossRef]
- Godinho, B.T.V.; Santos, I.A.F.M.; Gomes, E.A.; Picooli, R.H.; Cardoso, P.G. Endophytic fungi community in Eremanthus erythropappus tree from anthropogenic and natural areas of Minas Gerais. CERNE 2019, 25, 283–293. [Google Scholar] [CrossRef]
- Nicoletti, R. Occurrence and Functions of Endophytic Fungi in Crop Species. Agriculture 2020, 11, 18. [Google Scholar] [CrossRef]
- Fontana, D.C.; De Paula, S.; Torres, A.G.; De Souza, V.H.M.; Pascholati, S.F.; Schmidt, D.; Dourado Neto, D. Endophytic Fungi: Biological Control and Induced Resistance to Phytopathogens and Abiotic Stresses. Pathogens 2021, 10, 570. [Google Scholar] [CrossRef]
- Martin, F.M.; Uroz, S.; Barker, D.G. Ancestral alliances: Plant mutualistic symbioses with fungi and bacteria. Science 2017, 356, eaad4501. [Google Scholar] [CrossRef]
- Dos Santos, E.A.; De Lima, D.P.; Silva, D.B.; Marques, M.R.; Rodrigues, A. Dal’O. Plant–Microbe Interactions for Bioremediation of Pesticides. In Rhizobiont in Bioremediation of Hazardous Waste; Kumar, V., Prasad, R., Kumar, M., Eds.; Springer: Singapore, 2021; pp. 1–24. [Google Scholar] [CrossRef]
- Thapa, S.; Prasanna, R. Prospecting the characteristics and significance of the phyllosphere microbiome. Ann. Microbiol. 2018, 68, 229–245. [Google Scholar] [CrossRef]
- Rodrigues, A.D.O.; Montanholi, A.S.; Shimabukuro, A.A.; Yonekawa, M.K.A.; Cassemiro, N.S.; Silva, D.B.; Marchetti, C.R.; Weirich, C.E.; Beatriz, A.; Zanoelo, F.F.; et al. N-acetylation of Toxic Aromatic Amines by Fungi: Strain Screening, Cytotoxicity and Genotoxicity Evaluation, and Application in Bioremediation of 3,4-dichloroaniline. J. Hazard. Mater. 2023, 441, 129887. [Google Scholar] [CrossRef] [PubMed]
- Peay, K.G.; Kennedy, P.G.; Talbot, J.M. Dimensions of biodiversity in the Earth mycobiome. Nat. Rev. Microbiol. 2016, 14, 434–447. [Google Scholar] [CrossRef] [PubMed]
- Wężowicz, K.; Rozpądek, P.; Turnau, K. Interactions of arbuscular mycorrhizal and endophytic fungi improve seedling survival and growth in post-mining waste. Mycorrhiza 2017, 27, 499–511. [Google Scholar] [CrossRef]
- Wang, F. Occurrence of arbuscular mycorrhizal fungi in mining-impacted sites and their contribution to ecological restoration: Mechanisms and applications. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1901–1957. [Google Scholar] [CrossRef]
- Nirola, R.; Megharaj, M.; Beecham, S.; Aryal, R.; Thavamani, P.; Vankateswarlu, K.; Saint, C. Remediation of metalliferous mines, revegetation challenges and emerging prospects in semi-arid and arid conditions. Environ. Sci. Pollut. Res. 2016, 23, 20131–20150. [Google Scholar] [CrossRef]
- Boldt-Burisch, K.; Naeth, M.A. Early colonization of root associated fungal communities on reclamation substrates at a diamond mine in the Canadian Sub-Arctic. Appl. Soil Ecol. 2017, 110, 118–126. [Google Scholar] [CrossRef]
- Khalid, M.; Ur-Rahman, S.; Hassani, D.; Hayat, K.; Zhou, P.; Hui, N. Advances in fungal-assisted phytoremediation of heavy metals: A review. Pedosphere 2021, 31, 475–495. [Google Scholar] [CrossRef]
- Xia, Y.; Amna, A.; Opiyo, S.O. The culturable endophytic fungal communities of switchgrass grown on a coal-mining site and their effects on plant growth. PLoS ONE 2018, 13, e0198994. [Google Scholar] [CrossRef]
- Colin, Y.; Tétrault, M.-P.; Fauchery, L.; Buée, M.; Uroz, S. Forest plant cover and mineral type determine the diversity and composition of mineral-colonizing fungal communities. Eur. J. Soil Biol. 2021, 105, 103334. [Google Scholar] [CrossRef]
- Liu, B.; Yao, J.; Ma, B.; Li, S.; Duran, R. Disentangling biogeographic and underlying assembly patterns of fungal communities in metalliferous mining and smelting soils. Sci. Total Environ. 2022, 845, 157151. [Google Scholar] [CrossRef] [PubMed]
- Kalu, C.M.; Ogola, H.J.O.; Selvarajan, R.; Memory Tekere, M.; Ntushelo, K. Fungal and metabolome diversity of the rhizosphere and endosphere of Phragmites australis in an AMD-polluted environment. Heliyon 2021, 7, e06399. [Google Scholar] [CrossRef] [PubMed]
- Hind, D.J.N.; Miranda, E.B. Lista Preliminar da Família Compositae na Região Nordeste do Brasil: Preliminary List of the Compositae in Northeastern Brazil; Royal Botanic Gardens: Kew, Australia, 2008; pp. 20–55.
- Dos Santos, J.U.M. O Gênero Aspilia Thou. (Asteraceae–Heliantheae) No Brasil; Museu Paraense Emílio Goeldi-Funtec: Belém, Brazil, 2001; pp. 40–45.
- Centro Nacional de Conservação da Flor. Projeto Lista Vermelha. 2013. Available online: http://cncflora.jbrj.gov.br/portal/pt-br/profile/Aspilia%20grazielae (accessed on 14 September 2022).
- BRASIL. Lista Oficial de Espécies da Flora Ameaçadas de Extinção. Portaria n. 148, de 7 de Junho 2022. Available online: https://www.icmbio.gov.br/cepsul/images/stories/legislacao/Portaria/2020/P_mma_148_2022_altera_anexos_P_mma_443_444_445_2014_atualiza_especies_ameacadas_extincao.pdf (accessed on 14 October 2022).
- Skirycz, A.; Castilho, A.; Chaparro, C.; Carvalho, N.; Tzotzos, G.; Siqueira, J.O. Canga biodiversity, a matter of mining. Front. Plant Sci. 2014, 5, 653. [Google Scholar] [CrossRef] [PubMed]
- Vieira, C.K.; Marascalchi, M.N.; Rodrigues, A.V.; de Armas, R.D.; Stürmer, S.L. Morphological and molecular diversity of arbuscular mycorrhizal fungi in revegetated iron-mining site has the same magnitude of adjacent pristine ecosystems. J. Environ. Sci. 2018, 67, 330–343. [Google Scholar] [CrossRef]
- Graciolli, G.; Roque, F.O.; Farinaccio, M.A.; de Souza, P.R.; Pinto, J.O.P. Biota-MS: Montando o quebra-cabeça da biodiversidade de Mato Grosso do Sul. Iheringia Sér. Zool. 2017, 107, e2017100. [Google Scholar] [CrossRef]
- Pott, A.; Oliveira, A.; Damasceno-Junior, G.; Silva, J. Plant diversity of the Pantanal wetland. Braz. J. Biol. 2011, 71, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Haralyi, N.L.E.; Walde, D.H.G. Os minerais de ferro e manganês da região de Urucum, Corumbá, Mato Grosso do Sul. In Principais Depósitos Minerais do Brasil, II; Schobbenhaus, C., Coelho, C.E.S., Eds.; DNPM: Brasília, Brazil, 1986; pp. 127–144. [Google Scholar]
- Raij, B.; Van Andrade, J.C.; Cantarella, H.; Quaggio, J.Á. Análise Química para Avaliação da Fertilidade de Solos Tropicais; Campinas Instituto Agronômico: São Paulo, Brazil, 2001; pp. 5–277.
- Teixeira, P.C.; Donagemma, G.K.; Fontana, A.; Teixeira, W.G. Manual de Métodos de Análise de Solo, 3rd ed.; Embrapa Informação Tecnológica: Brasília, Brazil, 2017; pp. 20–426. [Google Scholar]
- Hawkins, C.L.; Davies, M.J. Hypochlorite-Induced Damage to DNA, RNA, and Polynucleotides: Formation of Chloramines and Nitrogen-Centered Radicals. Chem. Res. Toxicol. 2002, 15, 83–92. [Google Scholar] [CrossRef]
- Osinnikova, D.N.; Moroshkina, E.B.; Mokronosova, E.S. Effect of sodium hypochlorite on nucleic acids of different primary and secondary structures. J. Phys. Conf. Ser. 2019, 1400, 033001. [Google Scholar] [CrossRef]
- Araujo, W.L.; Maccheroni, W., Jr.; Agualar-Wildoso, C.L.; Barroso, P.A.V.; Saridakis, H.O.; Azevedo, J.L. Variability and interaction between endophytic bacteria and fungi isolated from leaf tissues of citrus rootstocks. Can. J. Microbiol. 2001, 47, 229–236. [Google Scholar] [CrossRef]
- Pereira, J.O.; Azevedo, J.L.; Petrini, O. Endophytic fungi of Stylosanthes: A First Report. Mycologia 1993, 85, 362–364. [Google Scholar] [CrossRef]
- Al-Bulushi, I.M.; Bani-Uraba, M.S.; Guizani, N.S.; Al-Khusaibi, M.K.; Al-Sadi, A.M. Illumina MiSeq sequencing analysis of fungal diversity in stored dates. BMC Microbiol. 2017, 17, 72. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedarsoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Beiko, R.G. Identifying biologically relevant differences between metagenomic communities. Bioinformatics 2010, 26, 715–721. [Google Scholar] [CrossRef]
- Boraks, A.; Amend, A.S. Fungi in soil and understory have coupled distribution pattern. PeerJ 2021, 9, e11915. [Google Scholar] [CrossRef] [PubMed]
- Nisa, H.; Kamili, A.N.; Nawchoo, I.A.; Shafi, S.; Shameem, N.; Bandh, A.S. Fungal endophytes as prolific source of phytochemicals and other bioactive natural products: A review. Microb. Pathog. 2015, 82, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G.; Daisy, B. Bioprospecting for microbial endophytes and their natural products. Microbiol. Mol. Biol. Rev. 2003, 67, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Murali, M.; Naziya, B.; Ansari, M.A.; Alomary, M.N.; AlYahya, S.; Almatroudi, A.; Thriveni, M.C.; Gowtham, H.G.; Singh, S.B.; Aiyaz, M.; et al. Bioprospecting of Rhizosphere-Resident Fungi: Their Role and Importance in Sustainable Agriculture. J. Fungi 2021, 7, 314. [Google Scholar] [CrossRef]
- Khan, A.R.; Ullah, I.; Waqas, M.; Park, G.S.; Khan, A.L.; Hong, S.J.; Ullah, R.; Jung, B.K.; Park, C.E.; Ur-Rehman, S.; et al. Host plant growth promotion and cadmium detoxification in Solanum nigrum, mediated by endophytic fungi. Ecotoxicol. Environ. Saf. 2017, 136, 180–188. [Google Scholar] [CrossRef]
- Zhao, Y.; Xiong, Z.; Wu, G.; Bai, W.; Zhu, Z.; Gao, Y.; Parmar, S.; Sharma, V.K.; Li, H. Fungal endophytic communities of two wild rose varieties with different powdery mildew susceptibilities. Front. Microb. 2018, 9, 2462. [Google Scholar] [CrossRef]
- Wehner, J.; Powell, J.R.; Muller, L.A.H.; Caruso, T.; Veresoglou, S.D.; Hempel, S.; Rillig, M.C. Determinants of root-associated fungal communities within Asteraceae in a semi-arid grassland. J. Ecol. 2014, 102, 425–436. [Google Scholar] [CrossRef]
- Wang, J.; Chen, C.; Ye, Z.; Li, J.; Feng, Y.; Lu, Q. Relationships between fungal and plant communities differ between desert and grassland in a typical dryland region of northwest China. Front. Microbiol. 2018, 9, 2327. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Cao, L. Fungal endophytes and their interactions with plants in phytoremediation: A review. Chemosphere 2017, 168, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, B.; Dong, Y.; Lin, H. Endophyte colonization enhanced cadmium phytoremediation by improving endosphere and rhizosphere microecology characteristics. J. Hazard. Mater. 2022, 434, 128829. [Google Scholar] [CrossRef] [PubMed]
- Nandy, S.; Das, T.; Tudu, C.K.; Pandey, D.K.; Dey, A.; Ray, P. Fungal endophytes: Futuristic tool in recent research area of phytoremediation. S. Afr. J. Bot. 2020, 134, 285–295. [Google Scholar] [CrossRef]
- He, W.; Megharaj, M.; Wu, C.Y.; Subashchandrabose, S.R.; Dai, C.C. Endophyte-assisted phytoremediation: Mechanisms and current application strategies for soil mixed pollutants. Crit. Rev. Biotechnol. 2020, 40, 31–45. [Google Scholar] [CrossRef]
- Shadmani, L.; Jamali, S.; Fatemi, A. Isolation, identification, and characterization of cadmium-tolerant endophytic fungi isolated from barley (Hordeum vulgare L.) roots and their role in enhancing phytoremediation. Braz. J. Microbiol. 2021, 52, 1097–1106. [Google Scholar] [CrossRef]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J. Trichoderma: The current status of its application in agriculture for the biocontrol of fungal phytopathogens and stimulation of plant growth. Int. J. Mol. Sci. 2022, 23, 2329. [Google Scholar] [CrossRef]
- Govarthanan, M.; Mythili, R.; Selvankumar, T.; Kamala-Kannan, S.; Kim, H. Myco-phytoremediation of arsenic-and lead-contaminated soils by Helianthus annuus and wood rot fungi, Trichoderma sp. isolated from decayed wood. Ecotoxicol. Environ. Saf. 2018, 151, 279–284. [Google Scholar] [CrossRef]
- Chomcheon, P.; Wiyakrutta, S.; Sriubolmas, N.; Ngamrojanavanich, N.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Metabolites from the endophytic mitosporic Dothideomycete sp. LRUB20. Phytochemistry 2009, 70, 121–127. [Google Scholar] [CrossRef]
- Sharma, D.; Pramanik, A.; Agrawal, P.K. Evaluation of bioactive secondary metabolites from endophytic fungus Pestalotiopsis neglecta BAB-5510 isolated from leaves of Cupressus torulosa D. Don. 3Biotech 2016, 6, 210. [Google Scholar] [CrossRef]
- Ali, A.; Guo, D.; Mahar, A.; Ma, F.; Li, R.; Shen, F.; Wang, P.; Zhang, Z. Streptomyces pactum assisted phytoremediation in Zn/Pb smelter contaminated soil of Feng County and its impact on enzymatic activities. Sci. Rep. 2017, 7, 46087. [Google Scholar] [CrossRef] [PubMed]
- Sim, C.S.F.; Cheow, Y.L.; Ng, S.L.; Ting, A.S.Y. Discovering metal-tolerant endophytic fungi from the phytoremediator plant Phragmites. Water Air Soil Pollut. 2018, 229, 68. [Google Scholar] [CrossRef]
- Ali, A.; Bilal, S.; Khan, A.L.; Mabood, F.; Al-Harrasi, A.; Lee, I.J. Endophytic Aureobasidium pullulans BSS6 assisted developments in phytoremediation potentials of Cucumis sativus under Cd and Pb stress. J. Plant Interact. 2019, 14, 303–313. [Google Scholar] [CrossRef]
- Lu, D.; Jin, H.; Yang, X.; Zhang, D.; Yan, Z.; Li, X.; Zhao, Y.; Han, R.; Qin, B. Characterization of rhizosphere and endophytic fungal communities from roots of Stipa purpurea in alpine steppe around Qinghai Lake. Can. J. Microbiol. 2016, 62, 643–656. [Google Scholar] [CrossRef] [PubMed]
- Tisthammer, K.; Cobian, G.M.; Amend, A.S. Global biogeography of marine fungi is shaped by the environment. Fungal Ecol. 2016, 19, 39–46. [Google Scholar] [CrossRef]
- Andrew, D.R.; Fitak, R.R.; Munguia-Veja, A.; Racolta, A.; Martinson, V.G.; Dontsova, K. Abiotic factors shape microbial diversity in Sonoran desert soils. Appl. Environ. Microbiol. 2012, 78, 7527–7537. [Google Scholar] [CrossRef]
- Aguilar-Trigueros, C.A.; Rillig, M.C. Effect of different root endophytic fungi on plant community structure in experimental microcosms. Ecol. Evol. 2016, 6, 8149–8158. [Google Scholar] [CrossRef]
- Liu, H.; Cheng, J.; Jin, H.; Xu, Z.; Yang, X.; Min, D.; Xu, X.; Shao, X.; Lu, D.; Qin, B. Characterization of Rhizosphere and Endophytic Microbial Communities Associated with Stipa purpurea and Their Correlation with Soil Environmental Factors. Plants 2022, 11, 363. [Google Scholar] [CrossRef]
- Egidi, E.; Delgado-Baquerizo, M.; Plett, J.M.; Wang, J.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K. A few Ascomycota taxa dominate soil fungal communities worldwide. Nat. Commun. 2019, 10, 2369. [Google Scholar] [CrossRef]
- Barnett, S.E.; Youngblut, N.D.; Buckley, D.H. Soil characteristics and land-use drive bacterial community assembly patterns. FEMS Microbiol. Ecol. 2019, 96, fiz194. [Google Scholar] [CrossRef]
- Sadet-Bourgeteau, S.; Houot, S.; Karimi, B.; Mathieu, O.; Mercier, V.; Montenache, D.; Morvanf, T.; Sappin-Didierg, V.; Watteauh, F.; Nowaka, V.; et al. Microbial communities from different soil types respond differently to organic waste input. Appl. Soil Ecol. 2019, 143, 70–79. [Google Scholar] [CrossRef]
- Leef, J.W.; Jones, S.E.; Prober, S.M.; Barberán, A.; Borer, E.T.; Firn, J.L.; Harpole, W.S.; Hobbie, S.E.; Hofmockel, K.S.; Knops, J.M.H.; et al. Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proc. Natl. Acad. Sci. USA 2015, 112, 10967–10972. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.D.; Terry, J.C.D.; Rossberg, A.G. Intrinsic ecological dynamics drive biodiversity turnover in model metacommunities. Nat. Commun. 2021, 12, 3627. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weirich, C.E.; Marques, M.R.; de Castro, A.P.; Assumpção Benitez, B.; Roque, F.d.O.; Marchetti, C.R.; Rodrigues, A.D.; de Lima, D.P.; dos Santos, E.d.A. Impact of Iron Mining Activity on the Endophytic Fungal Community of Aspilia grazielae. J. Fungi 2023, 9, 632. https://doi.org/10.3390/jof9060632
Weirich CE, Marques MR, de Castro AP, Assumpção Benitez B, Roque FdO, Marchetti CR, Rodrigues AD, de Lima DP, dos Santos EdA. Impact of Iron Mining Activity on the Endophytic Fungal Community of Aspilia grazielae. Journal of Fungi. 2023; 9(6):632. https://doi.org/10.3390/jof9060632
Chicago/Turabian StyleWeirich, Carlos Eduardo, Maria Rita Marques, Alinne Pereira de Castro, Beatriz Assumpção Benitez, Fabio de Oliveira Roque, Clarice Rossato Marchetti, Amanda Dal’Ongaro Rodrigues, Dênis Pires de Lima, and Edson dos Anjos dos Santos. 2023. "Impact of Iron Mining Activity on the Endophytic Fungal Community of Aspilia grazielae" Journal of Fungi 9, no. 6: 632. https://doi.org/10.3390/jof9060632