
Antagonistic Effect of Trichoderma longibrachiatum (TL6 and TL13) on Fusarium solani and Fusarium avenaceum Causing Root Rot on Snow Pea Plants
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates
2.1.1. Medium and Culture
2.1.2. Dual-Culture Assay of Antagonistic Fungi and Pathogens
2.1.3. Fungal Inoculum Preparation
2.2. Microscopic Observations of the Mycoparasitic Effects of TL6 and TL13 on F. solani and F. avenaceum
2.3. Effects of TL6 and TL13 on Snow Pea Seeds Germination under F. solani and F. avenaceum Infections
2.4. Growth Parameters
2.5. Disease Assessment
2.5.1. Determination of Chlorophyll Content and Carotenoid
2.5.2. Determination of MDA and H2O2 in Leaves
2.5.3. Antioxidant Enzyme Activities in Snow Pea Leaves
2.6. Statistical Analysis
3. Results
3.1. In Vitro Colony Growth Inhibition of TL6 and TL13 on F. avenaceum and F. solani
Microscopic Observations of the Mycoparasitic Effects of TL6 and TL13 on F. solani and F. avenaceum
3.2. Effects of TL6 and TL13 on Snow Pea Seeds Germination under F. solani and F. avenaceum Infections
3.3. Growth Parameters
3.4. Disease Assessment
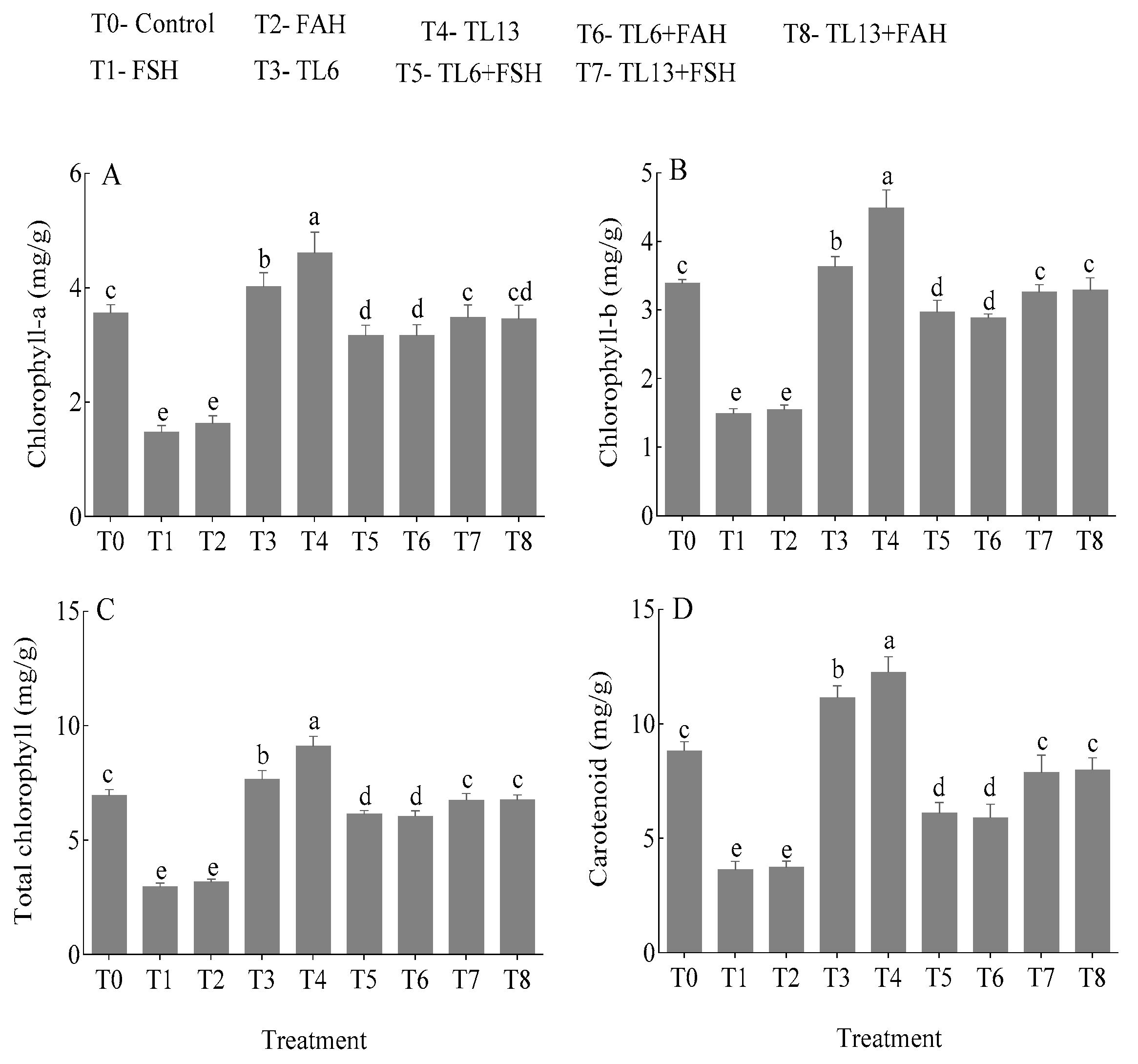
3.4.1. Chlorophyll and Carotenoid Contents
3.4.2. MDA and H2O2 Accumulation
3.5. Antioxidant Enzymes Activities
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, J.-p.; Qian, Y.-f.; Zhao, L.-y.; Chen, G.-t. Antidiabetic activities of glycoprotein from pea (Pisum sativum L.) in STZ-induced diabetic mice. Food Funct. 2021, 12, 5087–5095. [Google Scholar] [CrossRef]
- Tulbek, M.; Lam, R.; Asavajaru, P.; Lam, A. Pea: A sustainable vegetable protein crop. In Sustainable Protein Sources; Elsevier: Amsterdam, The Netherlands, 2017; pp. 145–164. [Google Scholar]
- Chaudhary, S.; Sagar, S.; Lal, M.; Tomar, A.; Kumar, V.; Kumar, M. Biocontrol and growth enhancement potential of Trichoderma spp. against Rhizoctonia solani causing sheath blight disease in rice. J. Environ. Biol. 2020, 41, 1034–1045. [Google Scholar] [CrossRef]
- Hegde, N.P. Evaluating Chemical Seed Treatments for Fusarium Root Rot Control in Dry Beans and Field Peas. Master’s Thesis, North Dakota State University, Fargo, ND, USA, 2014. [Google Scholar]
- El-Kazzaz, M.K.; Ghoneim, K.E.; Agha, M.K.M.; Helmy, A.; Behiry, S.I.; Abdelkhalek, A.; Saleem, M.H.; Al-Askar, A.A.; Arishi, A.A.; Elsharkawy, M.M. Suppression of Pepper Root Rot and Wilt Diseases Caused by Rhizoctonia solani and Fusarium oxysporum. Life 2022, 12, 587. [Google Scholar] [CrossRef]
- Safarieskandari, S. Towards Development of a Disease Risk Model for Pea Root Rot Disease. Ph.D. Thesis, University of Alberta, Edmonton, AB, Canada, 2019. [Google Scholar]
- Kamfwa, K.; Mwala, M.; Okori, P.; Gibson, P.; Mukankusi, C. Identification of QTL for Fusarium root rot resistance in common bean. J. Crop Improv. 2013, 27, 406–418. [Google Scholar] [CrossRef]
- Abeysinghe, S. Biological control of Fusarium solani f. sp. phaseoli the causal agent of root rot of bean using Bacillus subtilis CA32 and Trichoderma harzianum RU01. Ruhuna J. Sci. 2012, 2, 82–88. [Google Scholar]
- Eke, P.; Chatue, G.C.; Wakam, L.N.; Kouipou, R.M.T.; Fokou, P.V.T.; Boyom, F.F. Mycorrhiza consortia suppress the Fusarium root rot (Fusarium solani f. sp. Phaseoli) in common bean (Phaseolus vulgaris L.). Biol. Control 2016, 103, 240–250. [Google Scholar] [CrossRef]
- Saghafi, D.; Ghorbanpour, M.; Lajayer, B.A. Efficiency of Rhizobium strains as plant growth promoting rhizobacteria on morpho-physiological properties of Brassica napus L. under salinity stress. J. Soil Sci. Plant Nutr. 2018, 18, 253–268. [Google Scholar] [CrossRef] [Green Version]
- Boamah, S.; Zhang, S.; Xu, B.; Li, T.; Calderón-Urrea, A. Trichoderma longibrachiatum (TG1) Enhances wheat seedlings tolerance to salt stress and resistance to Fusarium pseudograminearum. Front. Plant Sci. 2021, 12, 741231. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, T.; Liu, J.; Liu, X.; Ou, Y.; Zhang, H.; Li, M.; Sonnewald, U.; Song, B.; Xie, C. Subtle regulation of potato acid invertase activity by a protein complex of invertase, invertase inhibitor, and sucrose nonfermenting1-related protein kinase. Plant Physiol. 2015, 168, 1807–1819. [Google Scholar] [CrossRef] [Green Version]
- Ramegowda, V.; Senthil-Kumar, M. The interactive effects of simultaneous biotic and abiotic stresses on plants: Mechanistic understanding from drought and pathogen combination. J. Plant Physiol. 2015, 176, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Wakeel, A.; Xu, M.; Gan, Y. Chromium-induced reactive oxygen species accumulation by altering the enzymatic antioxidant system and associated cytotoxic, genotoxic, ultrastructural, and photosynthetic changes in plants. Int. J. Mol. Sci. 2020, 21, 728. [Google Scholar] [CrossRef]
- Yaman, S.O.; Ayhanci, A. Lipid peroxidation. In Accenting Lipid Peroxidation; IntechOpen: London, UK, 2021; pp. 1–11. [Google Scholar]
- Zhao, X.-F.; Lin, C.; Rehmani, M.I.; Wang, Q.-S.; WANG, S.-H.; HOU, P.-F.; LI, G.-H.; DING, Y.-F. Effect of nitric oxide on alleviating cadmium toxicity in rice (Oryza sativa L.). J. Integr. Agric. 2013, 12, 1540–1550. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, K.K.; Chaudhary, N.; Agrawal, S.; Agrawal, M. Reactive oxygen species: Generation, damage, and quenching in plants during stress. In Reactive Oxygen Species in Plants: Boon or Bane-Revisiting the Role of ROS; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 89–115. [Google Scholar]
- Ahanger, M.A.; Morad-Talab, N.; Abd-Allah, E.F.; Ahmad, P.; Hajiboland, R. Plant growth under drought stress: Significance of mineral nutrients. Water Stress Crop Plants Sustain. Approach 2016, 2, 649–668. [Google Scholar]
- Hossain, M.A.; Bhattacharjee, S.; Armin, S.-M.; Qian, P.; Xin, W.; Li, H.-Y.; Burritt, D.J.; Fujita, M.; Tran, L.-S.P. Hydrogen peroxide priming modulates abiotic oxidative stress tolerance: Insights from ROS detoxification and scavenging. Front. Plant Sci. 2015, 6, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afzal, I.; Sabir, A.; Sikandar, S. Trichoderma: Biodiversity, Abundances, and Biotechnological Applications. In Recent Trends in Mycological Research; Springer: Berlin/Heidelberg, Germany, 2021; pp. 293–315. [Google Scholar]
- Sachdev, S.; Singh, R.P. Trichoderma: A multifaceted fungus for sustainable agriculture. In Ecological and Practical Applications for Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2020; pp. 261–304. [Google Scholar]
- Poveda, J.; Abril-Urias, P.; Escobar, C. Biological control of plant-parasitic nematodes by filamentous fungi inducers of resistance: Trichoderma, mycorrhizal and endophytic fungi. Front. Microbiol. 2020, 11, 992. [Google Scholar] [CrossRef]
- Card, S.; Johnson, L.; Teasdale, S.; Caradus, J. Deciphering endophyte behaviour: The link between endophyte biology and efficacious biological control agents. FEMS Microbiol. Ecol. 2016, 92, fiw114. [Google Scholar] [CrossRef] [Green Version]
- Puyam, A. Advent of Trichoderma as a bio-control agent-a review. J. Appl. Nat. Sci. 2016, 8, 1100–1109. [Google Scholar] [CrossRef]
- Nakkeeran, S.; Renukadevi, P.; Aiyanathan, K. Exploring the potential of Trichoderma for the management of seed and soil-borne diseases of crops. In Integrated Pest Management of Tropical Vegetable Crops; Springer: Berlin/Heidelberg, Germany, 2016; pp. 77–130. [Google Scholar]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]
- Rajani, P.; Rajasekaran, C.; Vasanthakumari, M.; Olsson, S.B.; Ravikanth, G.; Shaanker, R.U. Inhibition of plant pathogenic fungi by endophytic Trichoderma spp. through mycoparasitism and volatile organic compounds. Microbiol. Res. 2021, 242, 126595. [Google Scholar] [CrossRef]
- Sharma, A.K.; Sharma, P. Trichoderma; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Guzmán-Guzmán, P.; Porras-Troncoso, M.D.; Olmedo-Monfil, V.; Herrera-Estrella, A. Trichoderma species: Versatile plant symbionts. Phytopathology 2019, 109, 6–16. [Google Scholar] [CrossRef] [Green Version]
- Cai, F.; Chen, W.; Wei, Z.; Pang, G.; Li, R.; Ran, W.; Shen, Q. Colonization of Trichoderma harzianum strain SQR-T037 on tomato roots and its relationship to plant growth, nutrient availability and soil microflora. Plant Soil 2015, 388, 337–350. [Google Scholar] [CrossRef]
- Tucci, M.; Ruocco, M.; De Masi, L.; De Palma, M.; Lorito, M. The beneficial effect of Trichoderma spp. on tomato is modulated by the plant genotype. Mol. Plant Pathol. 2011, 12, 341–354. [Google Scholar] [CrossRef]
- Tripathi, P.; Singh, P.C.; Mishra, A.; Chauhan, P.S.; Dwivedi, S.; Bais, R.T.; Tripathi, R.D. Trichoderma: A potential bioremediator for environmental clean up. Clean Technol. Environ. Policy 2013, 15, 541–550. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Jambhulkar, P.P.; Raja, M.; Singh, B.; Katoch, S.; Kumar, S.; Sharma, P. Potential native Trichoderma strains against Fusarium verticillioides causing post flowering stalk rot in winter maize. Crop Prot. 2022, 152, 105838. [Google Scholar] [CrossRef]
- Lu, Z.-X.; TU, G.-P.; ZHANG, T.; LI, Y.-Q.; WANG, X.-H.; ZHANG, Q.-G.; Wei, S.; Jie, C. Screening of antagonistic Trichoderma strains and their application for controlling stalk rot in maize. J. Integr. Agric. 2020, 19, 145–152. [Google Scholar] [CrossRef]
- Rahman, M.; Begum, M.; Alam, M. Screening of Trichoderma isolates as a biological control agent against Ceratocystis paradoxa causing pineapple disease of sugarcane. Mycobiology 2009, 37, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Gan, Y.; Xu, B.; Xue, Y. The parasitic and lethal effects of Trichoderma longibrachiatum against Heterodera avenae. Biol. Control 2014, 72, 1–8. [Google Scholar] [CrossRef]
- Yassin, S.M.; Aly, A.; Abdel-Kader, D.A.; Morsy, K.; Atallah, O. Antagonistic potential of rhizospheric biocontrol agents against soybean root rot-wilt disease complex syndrome. Zagazig J. Agric. Res. 2019, 46, 1395–1418. [Google Scholar] [CrossRef]
- Zhang, S.; Gan, Y.; Xu, B. Mechanisms of the IAA and ACC-deaminase producing strain of Trichoderma longibrachiatum T6 in enhancing wheat seedling tolerance to NaCl stress. BMC Plant Biol. 2019, 19, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Oluwaranti, A.; Fakorede, M.; Adeboye, F. Maturity groups and phenology of maize in a rainforest location. Int. J. Agric. Innov. Res. 2015, 4, 124–127. [Google Scholar]
- Niu, X.; Mi, L.; Li, Y.; Wei, A.; Yang, Z.; Wu, J.; Zhang, D.; Song, X. Physiological and biochemical responses of rice seeds to phosphine exposure during germination. Chemosphere 2013, 93, 2239–2244. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Philpot, W.D. Relationship between surface soil water content, evaporation rate, and water absorption band depths in SWIR reflectance spectra. Remote Sens. Environ. 2015, 169, 280–289. [Google Scholar] [CrossRef]
- Zhang, X.-X.; Sun, H.-Y.; Shen, C.-M.; Li, W.; Yu, H.-S.; Chen, H.-G. Survey of Fusarium spp. causing wheat crown rot in major winter wheat growing regions of China. Plant Dis. 2015, 99, 1610–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miazek, K.; Ledakowicz, S. Chlorophyll extraction from leaves, needles and microalgae: A kinetic approach. Int. J. Agric. Biol. Eng. 2013, 6, 107–115. [Google Scholar]
- Monteiro, V.N.; do Nascimento Silva, R.; Steindorff, A.S.; Costa, F.T.; Noronha, E.F.; Ricart, C.A.O.; de Sousa, M.V.; Vainstein, M.H.; Ulhoa, C.J. New insights in Trichoderma harzianum antagonism of fungal plant pathogens by secreted protein analysis. Curr. Microbiol. 2010, 61, 298–305. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- García-Díaz, S.E.; Aldrete, A.; Alvarado-Rosales, D.; Cibrián-Tovar, D.; Mendez-Montiel, J.T.; Valdovinos-Ponce, G.; Equihua-Martinez, A. Effect of Fusarium circinatum on germination and growth of Pinus greggii seedlings in three substrates. Agrociencia 2017, 51, 895–908. [Google Scholar]
- Kaur, N.; Sehgal, S.; Glover, K.; Byamukama, E.; Ali, S. Impact of Fusarium graminearum on seed germination and seedling blight in hard red spring wheat in south Dakota. JPPM 2020, 11, 495. [Google Scholar]
- Worlu, C.; Nwauzoma, A.; Chuku, E.; Ajuru, M. Comparative Effects of Trichoderma species on Growth Parameters and Yield of Zea mays (L.). Int. J. Biol. Med. Sci. 2022, 5, 1–9. [Google Scholar]
- Sánchez-Montesinos, B.; Diánez, F.; Moreno-Gavíra, A.; Gea, F.J.; Santos, M. Role of Trichoderma aggressivum f. europaeum as Plant-Growth Promoter in Horticulture. Agronomy 2020, 10, 1004. [Google Scholar] [CrossRef]
- Erazo, J.; Palacios, S.; Pastor, N.; Giordano, F.; Rovera, M.; Reynoso, M.; Venisse, J.; Torres, A. Biocontrol mechanisms of Trichoderma harzianum ITEM 3636 against peanut brown root rot caused by Fusarium solani RC 386. Biol. Control 2021, 164, 104774. [Google Scholar] [CrossRef]
- Jaiswal, A.K.; Graber, E.R.; Elad, Y.; Frenkel, O. Biochar as a management tool for soilborne diseases affecting early stage nursery seedling production. Crop Prot. 2019, 120, 34–42. [Google Scholar] [CrossRef]
- Mayo, S.; Gutierrez, S.; Malmierca, M.G.; Lorenzana, A.; Campelo, M.P.; Hermosa, R.; Casquero, P.A. Influence of Rhizoctonia solani and Trichoderma spp. in growth of bean (Phaseolus vulgaris L.) and in the induction of plant defense-related genes. Front. Plant Sci. 2015, 6, 685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youssef, S.A.; Tartoura, K.A.; Abdelraouf, G.A. Evaluation of Trichoderma harzianum and Serratia proteamaculans effect on disease suppression, stimulation of ROS-scavenging enzymes and improving tomato growth infected by Rhizoctonia solani. Biol. Control 2016, 100, 79–86. [Google Scholar] [CrossRef]
- Nostar, O.; Ozdemir, F.; Bor, M.; Turkan, I.; Tosun, N. Combined effects of salt stress and cucurbit downy mildew (Pseudoperonospora cubensis Berk. and Curt. Rostov.) infection on growth, physiological traits and antioxidant activity in cucumber (Cucumis sativus L.) seedlings. Physiol. Mol. Plant Pathol. 2013, 83, 84–92. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.; Alqarawi, A.; Al Huqail, A.A.; Egamberdieva, D. Alleviation of abiotic salt stress in Ochradenus baccatus (Del.) by Trichoderma hamatum (Bonord.) Bainier. J. Plant Interact. 2014, 9, 857–868. [Google Scholar] [CrossRef]
- Saidimoradi, D.; Ghaderi, N.; Javadi, T. Salinity stress mitigation by humic acid application in strawberry (Fragaria x ananassa Duch.). Sci. Hortic. 2019, 256, 108594. [Google Scholar] [CrossRef]
- Rawat, L.; Singh, Y.; Shukla, N.; Kumar, J. Seed biopriming with salinity tolerant isolates of Trichoderma harzianum alleviates salt stress in rice: Growth, physiological and biochemical characteristics. J. Plant Pathol. 2012, 94, 353–365. [Google Scholar]
- Zhang, F.; Ge, H.; Zhang, F.; Guo, N.; Wang, Y.; Chen, L.; Ji, X.; Li, C. Biocontrol potential of Trichoderma harzianum isolate T-aloe against Sclerotinia sclerotiorum in soybean. Plant Physiol. Biochem. 2016, 100, 64–74. [Google Scholar] [CrossRef]
- Kumar, V.; Parkhi, V.; Kenerley, C.M.; Rathore, K.S. Defense-related gene expression and enzyme activities in transgenic cotton plants expressing an endochitinase gene from Trichoderma virens in response to interaction with Rhizoctonia solani. Planta 2009, 230, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Burman, U.; Kumar Garg, B.; Kathju, S. Interactive effects of phosphorus, nitrogen, and thiourea on clusterbean (Cyamopsis tetragonoloba L.) under rainfed conditions of the Indian arid zone. J. Plant Nutr. Soil Sci. 2007, 170, 803–810. [Google Scholar] [CrossRef]
- Seckin, B.; Sekmen, A.H.; Türkan, I. An enhancing effect of exogenous mannitol on the antioxidant enzyme activities in roots of wheat under salt stress. J. Plant Growth Regul. 2009, 28, 12–20. [Google Scholar] [CrossRef]
- Singh, A.; Sharma, P.; Meena, M.; Kumar, A.; Mishra, A.; Kumar, P.; Chaudhari, S.; Sharma, D. Effect of salinity on gas exchange parameters and ionic relations in bael (Aegle marmelos Correa). Indian J. Hort 2016, 73, 48–53. [Google Scholar] [CrossRef]
- Mann, A.; Kaur, G.; Kumar, A.; Sanwal, S.K.; Singh, J.; Sharma, P. Physiological response of chickpea (Cicer arietinum L.) at early seedling stage under salt stress conditions. Legume Res. Int. J. 2019, 42, 625–632. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, S.K.; Lata, C.; Devi, R.; Kulshrestha, N.; Krishnamurthy, S.; Singh, K.; Yadav, R.K. Impact of water deficit (salt and drought) stress on physiological, biochemical and yield attributes on wheat (Triticum aestivum) varieties. Indian J. Agric. Sci. 2018, 88, 1624–1632. [Google Scholar]
- Guler, N.S.; Pehlivan, N.; Karaoglu, S.A.; Guzel, S.; Bozdeveci, A. Trichoderma atroviride ID20G inoculation ameliorates drought stress-induced damages by improving antioxidant defence in maize seedlings. Acta Physiol. Plant. 2016, 38, 1–9. [Google Scholar] [CrossRef]
- Shukla, N.; Awasthi, R.; Rawat, L.; Kumar, J. Biochemical and physiological responses of rice (Oryza sativa L.) as influenced by Trichoderma harzianum under drought stress. Plant Physiol. Biochem. 2012, 54, 78–88. [Google Scholar] [CrossRef]
- Bacha, H.; Tekaya, M.; Drine, S.; Guasmi, F.; Touil, L.; Enneb, H.; Triki, T.; Cheour, F.; Ferchichi, A. Impact of salt stress on morpho-physiological and biochemical parameters of Solanum lycopersicum cv. Microtom leaves. S. Afr. J. Bot. 2017, 108, 364–369. [Google Scholar] [CrossRef]
- Kusvuran, S.; Kiran, S.; Ellialtioglu, S.S.J.A.; Advances, B.S.i.P.R.; Perspectives, F. Antioxidant enzyme activities and abiotic stress tolerance relationship in vegetable crops. In Abiotic and Biotic Stress in Plants—Recent Advances and Future Perspectives; IntechOpen: London, UK, 2016; pp. 481–506. [Google Scholar]
- Zhang, S.; Xu, B.; Gan, Y. Seed treatment with Trichoderma longibrachiatum T6 promotes wheat seedling growth under NaCl stress through activating the enzymatic and nonenzymatic antioxidant defense systems. Int. J. Mol. Sci. 2019, 20, 3729. [Google Scholar] [CrossRef] [Green Version]
- Luan, J.; Dong, J.; Song, X.; Jiang, J.; Li, H. Overexpression of tamarix hispida ThTrx5 confers salt tolerance to Arabidopsis by activating stress response signals. Int. J. Mol. Sci. 2020, 21, 1165. [Google Scholar] [CrossRef] [Green Version]
- Chowdappa, P.; Kumar, S.M.; Lakshmi, M.J.; Upreti, K. Growth stimulation and induction of systemic resistance in tomato against early and late blight by Bacillus subtilis OTPB1 or Trichoderma harzianum OTPB3. Biol. Control 2013, 65, 109–117. [Google Scholar] [CrossRef]
- Nawrocka, J.; Małolepsza, U.; Szymczak, K.; Szczech, M. Involvement of metabolic components, volatile compounds, PR proteins, and mechanical strengthening in multilayer protection of cucumber plants against Rhizoctonia solani activated by Trichoderma atroviride TRS25. Protoplasma 2018, 255, 359–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekmen Cetinel, A.H.; Gokce, A.; Erdik, E.; Cetinel, B.; Cetinkaya, N. The Effect of Trichoderma citrinoviride Treatment under Salinity Combined to Rhizoctonia solani Infection in Strawberry (Fragaria x ananassa Duch.). Agronomy 2021, 11, 1589. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Name | Treatment |
|---|---|---|
| T0 | Negative control | Distilled water |
| T1 | Positive control | Fusarium solani (FSH) |
| T2 | Positive control | Fusarium avenaceum (FAH) |
| T3 | TL6 | Trichoderma longibrachiatum (TL6) |
| T4 | TL13 | Trichoderma longibrachiatum TL13 |
| T5 | TL6 + FSH | Inoculation of strain TL6 + FSH |
| T6 | TL6 + FAH | Inoculation of strain TL6 + FAH |
| T7 | TL13 + FSH | Inoculation of strain TL13 + FSH |
| T8 | TL13 + FAH | Inoculation of strain TL13 + FAH |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boakye, T.A.; Li, H.; Osei, R.; Boamah, S.; Min, Z.; Ni, C.; Wu, J.; Shi, M.; Qiao, W. Antagonistic Effect of Trichoderma longibrachiatum (TL6 and TL13) on Fusarium solani and Fusarium avenaceum Causing Root Rot on Snow Pea Plants. J. Fungi 2022, 8, 1148. https://doi.org/10.3390/jof8111148
Boakye TA, Li H, Osei R, Boamah S, Min Z, Ni C, Wu J, Shi M, Qiao W. Antagonistic Effect of Trichoderma longibrachiatum (TL6 and TL13) on Fusarium solani and Fusarium avenaceum Causing Root Rot on Snow Pea Plants. Journal of Fungi. 2022; 8(11):1148. https://doi.org/10.3390/jof8111148
Chicago/Turabian StyleBoakye, Thomas Afriyie, Huixia Li, Richard Osei, Solomon Boamah, Zhang Min, Chunhui Ni, Jin Wu, Mingming Shi, and Wanqiang Qiao. 2022. "Antagonistic Effect of Trichoderma longibrachiatum (TL6 and TL13) on Fusarium solani and Fusarium avenaceum Causing Root Rot on Snow Pea Plants" Journal of Fungi 8, no. 11: 1148. https://doi.org/10.3390/jof8111148