
Morphology, Phenotype, and Molecular Identification of Clinical and Environmental Fusarium solani Species Complex Isolates from Malaysia

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates
2.2. Morphological Examination
2.3. DNA Extraction and PCR Amplification
2.4. DNA Sequencing and Analysis of TEF1-α and RPB2 Sequences
2.5. Phylogenetic Analyses
2.6. Fusarium Growth Characteristics
3. Results
3.1. Molecular Identification
3.2. Morphological Characteristics
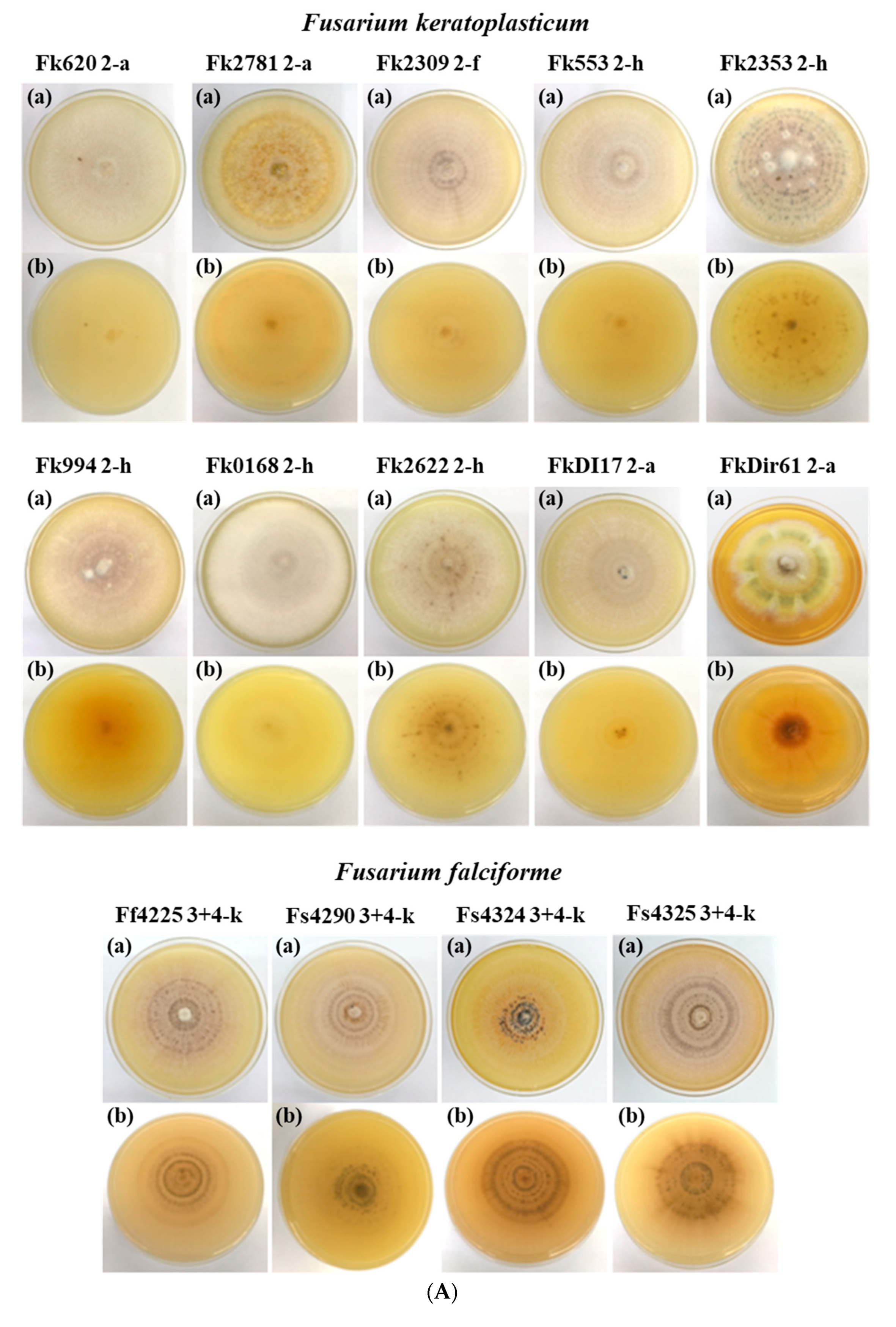
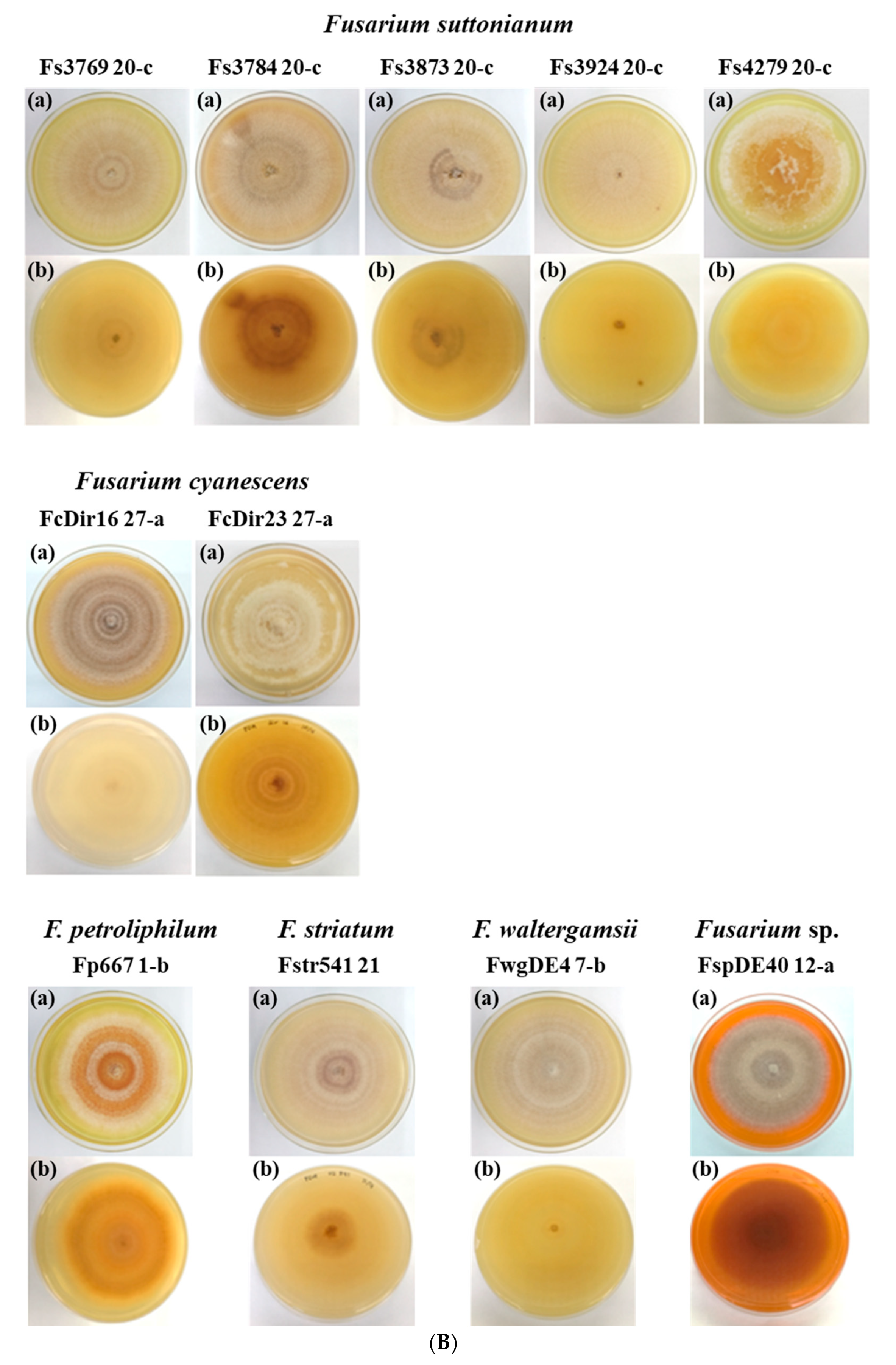
3.2.1. Colony Morphology
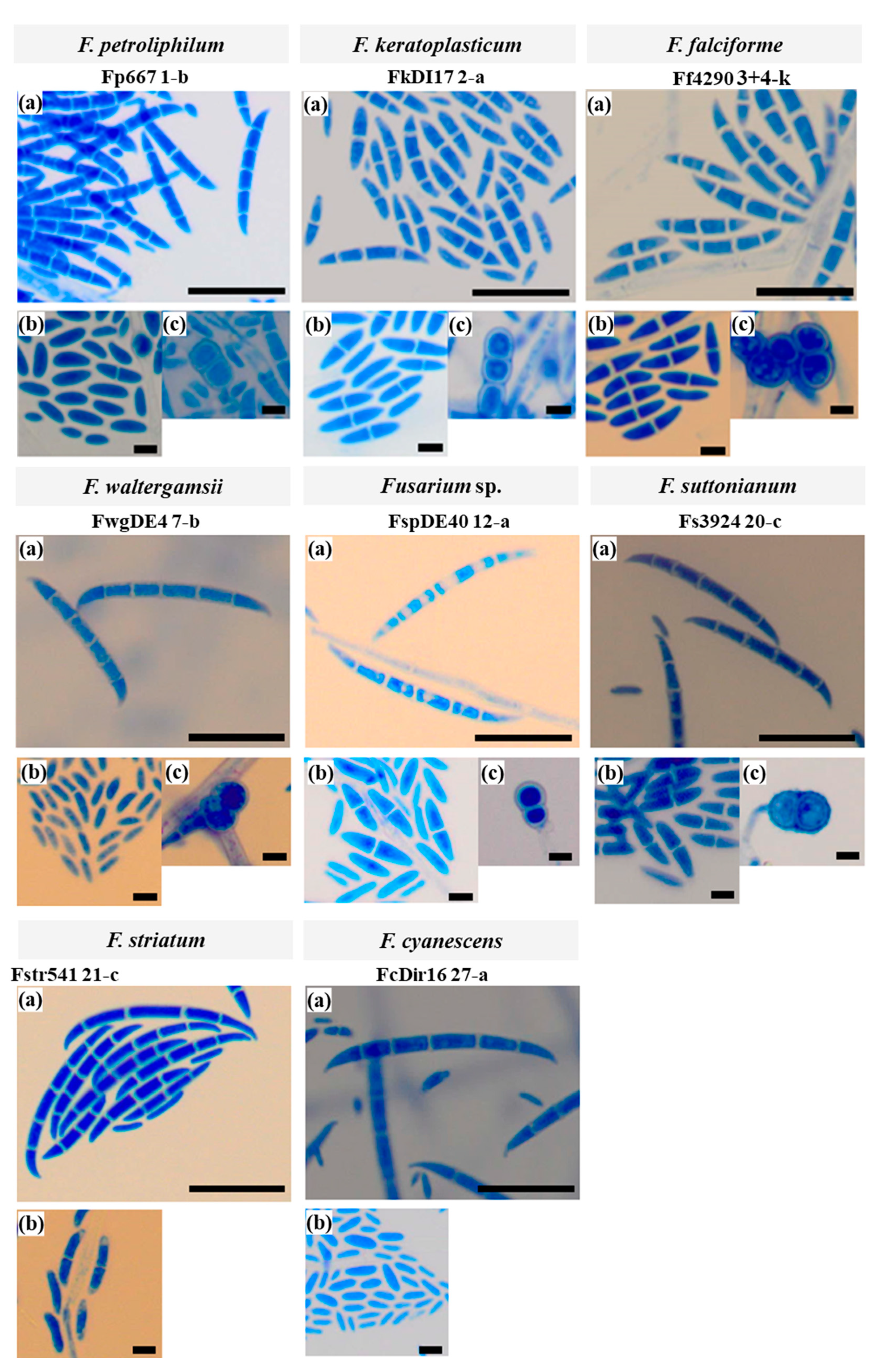
3.2.2. Microconidia and Chlamydospores
3.2.3. Macroconidia
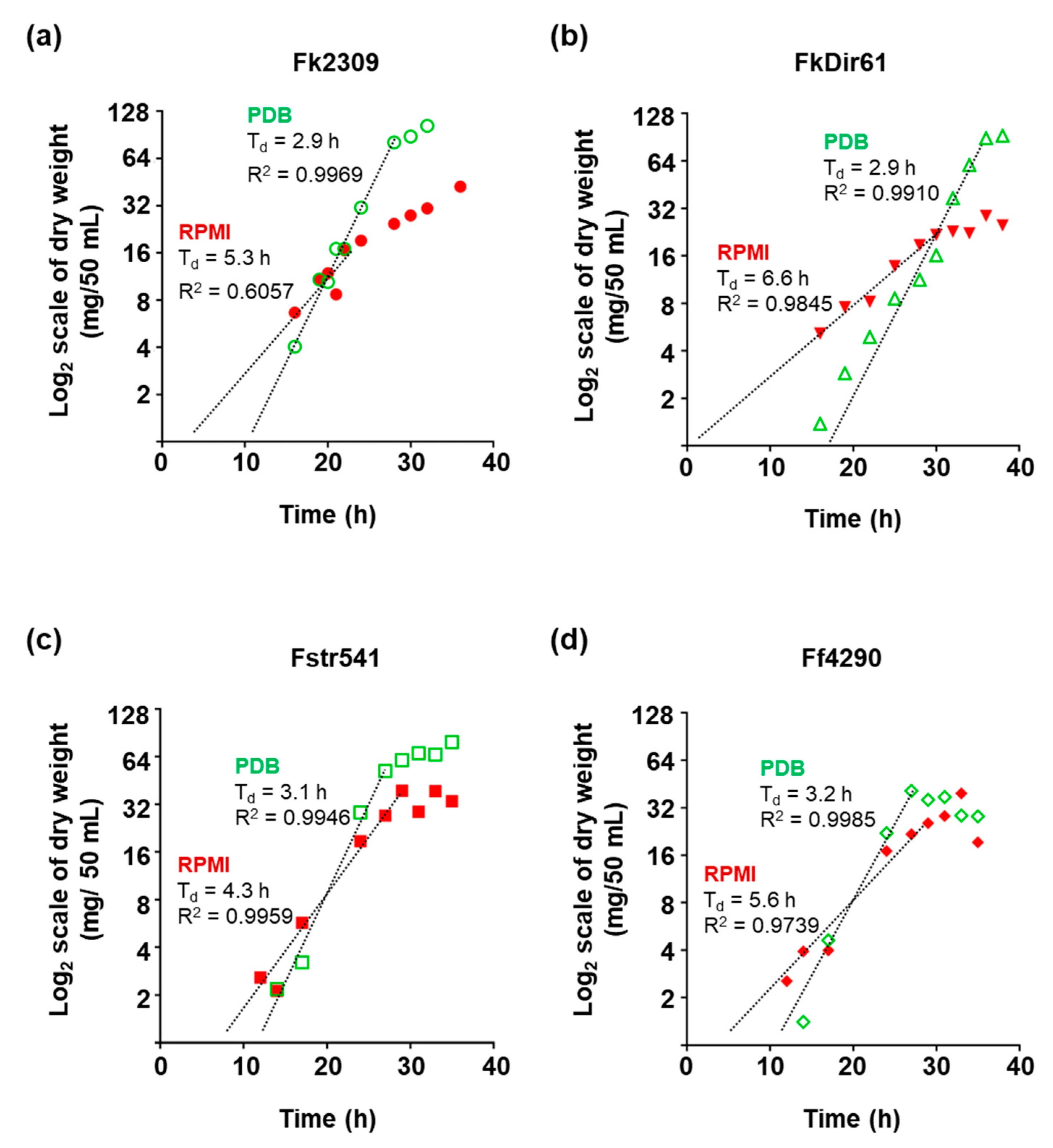
3.3. Growth of F. keratoplasticum, F. striatum and F. falciforme in PDB and RPMI
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coleman, J.J. The Fusarium solani species complex: Ubiquitous pathogens of agricultural importance. Mol. Plant Pathol. 2016, 17, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K.; Gardiner, D.M. Transcriptomics of cereal—Fusarium graminearum interactions: What we have learned so far. Mol. Plant Pathol. 2018, 19, 764–778. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak, C.; Massa, G.; Hummerick, M.; Khodadad, C.; Schuerger, A.; Venkateswaran, K. Draft genome sequences of two Fusarium oxysporum isolates cultured from infected Zinnia hybrida plants grown on the International Space Station. Genome Announc. 2018, 6, e00326-18. [Google Scholar] [CrossRef] [PubMed]
- Dweba, C.; Figlan, S.; Shimelis, H.; Motaung, T.; Sydenham, S.; Mwadzingeni, L.; Tsilo, T. Fusarium head blight of wheat: Pathogenesis and control strategies. Crop Prot. 2017, 91, 114–122. [Google Scholar] [CrossRef]
- Maryani, N.; Lombard, L.; Poerba, Y.; Subandiyah, S.; Crous, P.; Kema, G. Phylogeny and genetic diversity of the banana Fusarium wilt pathogen Fusarium oxysporum f. sp. cubense in the Indonesian centre of origin. Stud. Mycol. 2019, 92, 155–194. [Google Scholar] [CrossRef]
- Stover, R. Fusarium wilt of banana: Some history and current status of the disease. In Proceedings of the First International Conference on Fusarial Wilt of Banana, Miami, FL, USA, 27–30 August 1989; pp. 1–7. [Google Scholar]
- Windels, C.E. Economic and social impacts of Fusarium head blight: Changing farms and rural communities in the Northern Great Plains. Phytopathology 2000, 90, 17–21. [Google Scholar] [CrossRef]
- Ordonez, N.; Seidl, M.F.; Waalwijk, C.; Drenth, A.; Kilian, A.; Thomma, B.P.; Ploetz, R.C.; Kema, G.H. Worse comes to worst: Bananas and Panama disease-when plant and pathogen clones meet. PLoS Pathog. 2015, 11, e1005197. [Google Scholar] [CrossRef]
- Cook, D.C.; Taylor, A.S.; Meldrum, R.A.; Drenth, A. Potential economic impact of Panama disease (tropical race 4) on the Australian banana industry. J. Plant Dis. Prot. 2015, 122, 229–237. [Google Scholar] [CrossRef]
- Ploetz, R.C. Panama disease: An old nemesis rears its ugly head: Part 1. the beginnings of the banana export trades. Plant Health Prog. 2005, 6, 18. [Google Scholar] [CrossRef]
- Nucci, M.; Anaissie, E. Fusarium infections in immunocompromised patients. Clin. Microbiol. Rev. 2007, 20, 695–704. [Google Scholar] [CrossRef]
- Rotjanapan, P.; Chen, Y.C.; Chakrabarti, A.; Li, R.Y.; Rudramurthy, S.M.; Yu, J.; Kung, H.C.; Watcharananan, S.; Tan, A.L.; Saffari, S.E.; et al. Epidemiology and clinical characteristics of invasive mould infections: A multicenter, retrospective analysis in five Asian countries. Med. Mycol. 2018, 56, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Lass-Flörl, C.; Cuenca-Estrella, M. Changes in the epidemiological landscape of invasive mould infections and disease. J. Antimicrob. Chemother. 2017, 72, i5–i11. [Google Scholar] [CrossRef] [PubMed]
- Horn, D.L.; Freifeld, A.G.; Schuster, M.G.; Azie, N.E.; Franks, B.; Kauffman, C.A. Treatment and outcomes of invasive fusariosis: Review of 65 cases from the PATH Alliance((R)) registry. Mycoses 2014, 57, 652–658. [Google Scholar] [CrossRef]
- Nucci, M.; Anaissie, E. Cutaneous infection by Fusarium species in healthy and immunocompromised hosts: Implications for diagnosis and management. Clin. Infect. Dis. 2002, 35, 909–920. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Rooney, A.P.; Proctor, R.H.; Brown, D.W.; McCormick, S.P.; Ward, T.J.; Frandsen, R.J.; Lysøe, E.; Rehner, S.A.; Aoki, T.; et al. Phylogenetic analyses of RPB1 and RPB2 support a middle Cretaceous origin for a clade comprising all agriculturally and medically important fusaria. Fungal Genet. Biol. 2013, 52, 20–31. [Google Scholar] [CrossRef]
- O’Donnell, K.; Sutton, D.A.; Rinaldi, M.G.; Sarver, B.A.; Balajee, S.A.; Schroers, H.J.; Summerbell, R.C.; Robert, V.A.; Crous, P.W.; Zhang, N.; et al. Internet-accessible DNA sequence database for identifying fusaria from human and animal infections. J. Clin. Microbiol. 2010, 48, 3708–3718. [Google Scholar] [CrossRef]
- Schroers, H.J.; Samuels, G.J.; Zhang, N.; Short, D.P.; Juba, J.; Geiser, D.M. Epitypification of Fusisporium (Fusarium) solani and its assignment to a common phylogenetic species in the Fusarium solani species complex. Mycologia 2016, 108, 806–819. [Google Scholar] [CrossRef]
- Al-Hatmi, A.M.S.; Bonifaz, A.; Ranque, S.; Sybren de Hoog, G.; Verweij, P.E.; Meis, J.F. Current antifungal treatment of fusariosis. Int. J. Antimicrob. Agents 2018, 51, 326–332. [Google Scholar] [CrossRef]
- Matuo, T.; Snyder, W.C. Use of morphology and mating populations in the identification of formae speciales in Fusarium solani. J. Phytopathol. 1973, 63, 562–565. [Google Scholar] [CrossRef]
- Chang, D.C.; Grant, G.B.; O’Donnell, K.; Wannemuehler, K.A.; Noble-Wang, J.; Rao, C.Y.; Jacobson, L.M.; Crowell, C.S.; Sneed, R.S.; Lewis, F.M.; et al. Multistate outbreak of Fusarium keratitis associated with use of a contact lens solution. J. Am. Med. Assoc. 2006, 296, 953–963. [Google Scholar] [CrossRef]
- O’Donnell, K.; Sutton, D.A.; Fothergill, A.; McCarthy, D.; Rinaldi, M.G.; Brandt, M.E.; Zhang, N.; Geiser, D.M. Molecular phylogenetic diversity, multilocus haplotype nomenclature, and in vitro antifungal resistance within the Fusarium solani species complex. J. Clin. Microbiol. 2008, 46, 2477–2490. [Google Scholar] [CrossRef]
- O’Donnell, K. Molecular phylogeny of the Nectria haematococca-Fusarium solani species complex. Mycologia 2000, 92, 919–938. [Google Scholar] [CrossRef]
- Aoki, T.; O’Donnell, K.; Homma, Y.; Lattanzi, A.R. Sudden-death syndrome of soybean is caused by two morphologically and phylogenetically distinct species within the Fusarium solani species complex-F. virguliforme in North America and F. tucumaniae in South America. Mycologia 2003, 95, 660–684. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Denis, M.; Crous, P. Removing chaos from confusion: Assigning names to common human and animal pathogens in Neocosmospora. Persoonia 2018, 41, 109–129. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Lombard, L.; Crous, P.W. Back to the roots: A reappraisal of Neocosmospora. Persoonia 2019, 43, 90–185. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Al-Hatmi, A.M.S.; Aoki, T.; Brankovics, B.; Cano-Lira, J.F.; Coleman, J.J.; de Hoog, G.S.; Di Pietro, A.; Frandsen, R.J.N.; Geiser, D.M.; et al. No to Neocosmospora: Phylogenomic and practical reasons for continued inclusion of the Fusarium solani species complex in the genus Fusarium. mSphere 2020, 5, e00810-20. [Google Scholar] [CrossRef]
- Crous, P.W.; Lombard, L.; Sandoval-Denis, M.; Seifert, K.A.; Schroers, H.J.; Chaverri, P.; Gene, J.; Guarro, J.; Hirooka, Y.; Bensch, K.; et al. Fusarium: More than a node or a foot-shaped basal cell. Stud. Mycol. 2021, 98, 100116. [Google Scholar] [CrossRef]
- Espinel-Ingroff, A.; Colombo, A.L.; Cordoba, S.; Dufresne, P.J.; Fuller, J.; Ghannoum, M.; Gonzalez, G.M.; Guarro, J.; Kidd, S.E.; Meis, J.F.; et al. International evaluation of MIC distributions and epidemiological cutoff value (ECV) definitions for Fusarium species identified by molecular methods for the CLSI broth microdilution method. Antimicrob. Agents Chemother. 2016, 60, 1079–1084. [Google Scholar] [CrossRef]
- Blaize, M.; Normand, A.C.; Imbert, S.; Al-Hatmi, A.M.S.; Chryssanthou, E.; Cassaing, S.; Schuttler, C.; Hasseine, L.; Mahinc, C.; Costa, D.; et al. Antifungal susceptibility of 182 Fusarium species isolates from 20 European centers: Comparison between EUCAST and gradient concentration strip methods. Antimicrob. Agents Chemother. 2021, 65, e0149521. [Google Scholar] [CrossRef]
- Stempel, J.M.; Hammond, S.P.; Sutton, D.A.; Weiser, L.M.; Marty, F.M. Invasive fusariosis in the voriconazole era: Single-center 13-year experience. Open Forum Infect. Dis. 2015, 2, ofv099. [Google Scholar] [CrossRef]
- Lortholary, O.; Obenga, G.; Biswas, P.; Caillot, D.; Chachaty, E.; Bienvenu, A.L.; Cornet, M.; Greene, J.; Herbrecht, R.; Lacroix, C.; et al. International retrospective analysis of 73 cases of invasive fusariosis treated with voriconazole. Antimicrob. Agents Chemother. 2010, 54, 4446–4450. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Endo, T.; Hashimoto, D.; Saga, T.; Ara, T.; Ogasawara, R.; Yasumoto, A.; Ibata, M.; Takahata, M.; Shigematsu, A.; et al. Disseminated fusariosis emerged from prolonged local genital infection after cord blood transplantation. J. Infect. Chemother. 2018, 24, 660–663. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.M.; Silveira, A.R.; Betania, C.A.; Macedo, D.P.; Neves, R.P. Disseminated fusariosis secondary to neuroblastoma with fatal outcome. Mycopathologia 2013, 176, 233–236. [Google Scholar] [CrossRef]
- Esnakula, A.K.; Summers, I.; Naab, T.J. Fatal disseminated Fusarium infection in a human immunodeficiency virus positive patient. Case Rep. Infect. Dis. 2013, 2013, 379320. [Google Scholar] [CrossRef] [PubMed]
- Nucci, M.; Anaissie, E.J.; Queiroz-Telles, F.; Martins, C.A.; Trabasso, P.; Solza, C.; Mangini, C.; Simoes, B.P.; Colombo, A.L.; Vaz, J. Outcome predictors of 84 patients with hematologic malignancies and Fusarium infection. Cancer 2003, 98, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Bruhl, C.A.; Imfeld, G.; Knabel, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmuller, A.; et al. Fungicides: An Overlooked Pesticide Class? Environ. Sci. Technol. 2019, 53, 3347–3365. [Google Scholar] [CrossRef] [PubMed]
- Meis, J.F.; Chowdhary, A.; Rhodes, J.L.; Fisher, M.C.; Verweij, P.E. Clinical implications of globally emerging azole resistance in Aspergillus fumigatus. Philos. Trans. R Soc. Lond. B Biol. Sci. 2016, 371, 20150460. [Google Scholar] [CrossRef]
- Chowdhary, A.; Kathuria, S.; Xu, J.; Meis, J.F. Emergence of azole-resistant Aspergillus fumigatus strains due to agricultural azole use creates an increasing threat to human health. PLoS Pathog. 2013, 9, e1003633. [Google Scholar] [CrossRef]
- Cao, D.; Wang, F.; Yu, S.; Dong, S.; Wu, R.; Cui, N.; Ren, J.; Xu, T.; Wang, S.; Wang, M.; et al. Prevalence of azole-resistant Aspergillus fumigatus is highly associated with azole fungicide residues in the fields. Environ. Sci. Technol. 2021, 55, 3041–3049. [Google Scholar] [CrossRef]
- Zhang, J.; van den Heuvel, J.; Debets, A.J.M.; Verweij, P.E.; Melchers, W.J.G.; Zwaan, B.J.; Schoustra, S.E. Evolution of cross-resistance to medical triazoles in Aspergillus fumigatus through selection pressure of environmental fungicides. Proc. Biol. Sci. 2017, 284, 20170635. [Google Scholar] [CrossRef]
- Kang, S.E.; Sumabat, L.G.; Melie, T.; Mangum, B.; Momany, M.; Brewer, M.T. Evidence for the agricultural origin of resistance to multiple antimicrobials in Aspergillus fumigatus, a fungal pathogen of humans. G3 2022, 12, jkab427. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.; Jain, K.; Wang, Y.; Pawar, K.; Kaur, H.; Sharma, K.K.; Tripathy, V.; Singh, A.; Xu, J.; Chowdhary, A. Candida auris on apples: Diversity and clinical significance. mBio 2022, 13, e0051822. [Google Scholar] [CrossRef] [PubMed]
- Bastos, R.W.; Rossato, L.; Goldman, G.H.; Santos, D.A. Fungicide effects on human fungal pathogens: Cross-resistance to medical drugs and beyond. PLoS Pathog. 2021, 17, e1010073. [Google Scholar] [CrossRef] [PubMed]
- Mellado, E.; Diaz-Guerra, T.M.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. Identification of two different 14-alpha sterol demethylase-related genes (CYP51A and CYP51B) in Aspergillus fumigatus and other Aspergillus species. J. Clin. Microbiol. 2001, 39, 2431–2438. [Google Scholar] [CrossRef] [PubMed]
- James, J.E.; Lamping, E.; Santhanam, J.; Milne, T.J.; Abd Razak, M.F.; Zakaria, L.; Cannon, R.D. A 23 bp cyp51A promoter deletion associated with voriconazole resistance in clinical and environmental isolates of Neocosmospora keratoplastica. Front. Microbiol. 2020, 11, 272. [Google Scholar] [CrossRef]
- Becher, R.; Hettwer, U.; Karlovsky, P.; Deising, H.B.; Wirsel, S.G. Adaptation of Fusarium graminearum to tebuconazole yielded descendants diverging for levels of fitness, fungicide resistance, virulence, and mycotoxin production. Phytopathology 2010, 100, 444–453. [Google Scholar] [CrossRef]
- Chen, J.; Wei, J.; Fu, L.; Wang, S.; Liu, J.; Guo, Q.; Jiang, J.; Tian, Y.; Che, Z.; Chen, G.; et al. Tebuconazole resistance of Fusarium graminearum field populations from wheat in Henan Province. J. Phytopathol. 2021, 169, 525–532. [Google Scholar] [CrossRef]
- Al-Hatmi, A.M.; van Diepeningen, A.D.; Curfs-Breuker, I.; de Hoog, G.S.; Meis, J.F. Specific antifungal susceptibility profiles of opportunists in the Fusarium fujikuroi complex. J. Antimicrob. Chemother. 2015, 70, 1068–1071. [Google Scholar] [CrossRef]
- Hafizi, R.; Salleh, B.; Latiffah, Z. Morphological and molecular characterization of Fusarium solani and F. oxysporum associated with crown disease of oil palm. Braz. J. Microbiol. 2013, 44, 959–968. [Google Scholar] [CrossRef]
- Ibrahim, N.F.; Mohd, M.H.; Nor, N.M.I.M.; Zakaria, L. Fusarium fujikuroi causing fusariosis of pineapple in peninsular Malaysia. Australas. Plant Dis. Notes 2016, 11, 21. [Google Scholar] [CrossRef]
- Lim, G. Fusarium in paddy soils of West Malaysia. Plant Soil 1972, 36, 47–51. [Google Scholar] [CrossRef]
- Sidique, S.N.M.; Azuddin, N.F.; Joseph, J. First report of Fusarium species at nesting sites of endangered sea turtles in Terengganu and Melaka, Malaysia. Malays. Appl. Biol. 2017, 46, 195–205. [Google Scholar]
- Manshor, N.; Rosli, H.; Ismail, N.A.; Salleh, B.; Zakaria, L. Diversity of Fusarium species from highland areas in Malaysia. Trop. Life Sci. Res. 2012, 23, 1–15. [Google Scholar] [PubMed]
- Singh, H.; Jamal, F.; Marahakim, M.; Song, C. Fusarium solani keratitis. First report from Malaysia. Int. Med. J. Malays. 1981, 36, 89–91. [Google Scholar]
- Mohd-Tahir, F.; Norhayati, A.; Siti-Raihan, I.; Ibrahim, M. A 5-year retrospective review of fungal keratitis at Hospital Universiti Sains Malaysia. Interdiscip. Perspect. Infect. Dis. 2012, 2012, 851653. [Google Scholar] [CrossRef]
- Leslie, J.; Summerell, B. The Fusarium Laboratory Manual, 1st ed.; Blackwell Publishing: Ames, IA, USA, 2006; pp. 113–117. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Page, R.D. Tree View: An application to display phylogenetic trees on personal computers. J. Bioinform. 1996, 12, 357–358. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. J. Bioinform. 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A new and scalable tool for the selection of DNA and protein evolutionary models. Mol. Biol. Evol. 2019, 37, 291–294. [Google Scholar] [CrossRef]
- O’Donnell, K.; Humber, R.A.; Geiser, D.M.; Kang, S.; Park, B.; Robert, V.A.; Crous, P.W.; Johnston, P.R.; Aoki, T.; Rooney, A.P.; et al. Phylogenetic diversity of insecticolous fusaria inferred from multilocus DNA sequence data and their molecular identification via FUSARIUM-ID and Fusarium MLST. Mycologia 2012, 104, 427–445. [Google Scholar] [CrossRef] [PubMed]
- Herkert, P.F.; Al-Hatmi, A.M.S.; de Oliveira Salvador, G.L.; Muro, M.D.; Pinheiro, R.L.; Nucci, M.; Queiroz-Telles, F.; de Hoog, G.S.; Meis, J.F. Molecular characterization and antifungal susceptibility of clinical Fusarium species from Brazil. Front. Microbiol. 2019, 10, 737. [Google Scholar] [CrossRef] [PubMed]
- Chiewchanvit, S.; Chongkae, S.; Mahanupab, P.; Nosanchuk, J.D.; Pornsuwan, S.; Vanittanakom, N.; Youngchim, S. Melanization of Fusarium keratoplasticum (F. solani species complex) during disseminated fusariosis in a patient with acute leukemia. Mycopathologia 2017, 182, 879–885. [Google Scholar] [CrossRef]
- Tupaki-Sreepurna, A.; Al-Hatmi, A.M.; Kindo, A.J.; Sundaram, M.; de Hoog, G.S. Multidrug-resistant Fusarium in keratitis: A clinico-mycological study of keratitis infections in Chennai, India. Mycoses 2017, 60, 230–233. [Google Scholar] [CrossRef]
- Guevara-Suarez, M.; Cano-Lira, J.F.; de Garcia, M.C.; Sopo, L.; De Bedout, C.; Cano, L.E.; Garcia, A.M.; Motta, A.; Amezquita, A.; Cardenas, M.; et al. Genotyping of Fusarium isolates from onychomycoses in Colombia: Detection of two new species within the Fusarium solani species complex and in vitro antifungal susceptibility testing. Mycopathologia 2016, 181, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Al-Hatmi, A.M.; Bonifaz, A.; Tirado-Sanchez, A.; Meis, J.F.; de Hoog, G.S.; Ahmed, S.A. Fusarium species causing eumycetoma: Report of two cases and comprehensive review of the literature. Mycoses 2017, 60, 204–212. [Google Scholar] [CrossRef]
- O’Donnell, K.; Sutton, D.A.; Wiederhold, N.; Robert, V.A.; Crous, P.W.; Geiser, D.M. Veterinary fusarioses within the United States. J. Clin. Microbiol. 2016, 54, 2813–2819. [Google Scholar] [CrossRef]
- Homa, M.; Shobana, C.S.; Singh, Y.R.; Manikandan, P.; Selvam, K.P.; Kredics, L.; Narendran, V.; Vagvolgyi, C.; Galgoczy, L. Fusarium keratitis in South India: Causative agents, their antifungal susceptibilities and a rapid identification method for the Fusarium solani species complex. Mycoses 2013, 56, 501–511. [Google Scholar] [CrossRef]
- Homa, M.; Galgóczy, L.; Manikandan, P.; Narendran, V.; Sinka, R.; Csernetics, Á.; Vágvölgyi, C.; Kredics, L.; Papp, T. South Indian isolates of the Fusarium solani species complex from clinical and environmental samples: Identification, antifungal susceptibilities, and virulence. Front. Microbiol. 2018, 9, 1052. [Google Scholar] [CrossRef]
- Moretti, M.L.; Busso-Lopes, A.F.; Tararam, C.A.; Moraes, R.; Muraosa, Y.; Mikami, Y.; Gonoi, T.; Taguchi, H.; Lyra, L.; Reichert-Lima, F.; et al. Airborne transmission of invasive fusariosis in patients with hematologic malignancies. PLoS ONE 2018, 13, e0196426. [Google Scholar] [CrossRef]
- Ersal, T.; Al-Hatmi, A.S.; Cilo, B.D.; Curfs-Breuker, I.; Meis, J.F.; Ozkalemkas, F.; Ener, B.; van Diepeningen, A.D. Fatal disseminated infection with Fusarium petroliphilum. Mycopathologia 2015, 179, 119–124. [Google Scholar] [CrossRef] [PubMed]
- James, J.E.; Lamping, E.; Santhanam, J.; Cannon, R.D. PDR transporter ABC1 is involved in the innate azole resistance of the human fungal pathogen Fusarium keratoplasticum. Front. Microbiol. 2021, 12, 673206. [Google Scholar] [CrossRef] [PubMed]
- Summerell, B.A.; Salleh, B.; Leslie, J.F. A utilitarian approach to Fusarium identification. Plant Dis. 2003, 87, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Chehri, K.; Salleh, B.; Zakaria, L. Morphological and phylogenetic analysis of Fusarium solani species complex in Malaysia. Microb. Ecol. 2015, 69, 457–471. [Google Scholar] [CrossRef]
- Short, D.P.; O’Donnell, K.; Thrane, U.; Nielsen, K.F.; Zhang, N.; Juba, J.H.; Geiser, D.M. Phylogenetic relationships among members of the Fusarium solani species complex in human infections and the descriptions of F. keratoplasticum sp. nov. and F. petroliphilum stat. nov. Fungal Genet. Biol. 2013, 53, 59–70. [Google Scholar] [CrossRef]
- Nash, S.M.; Snyder, W.C. Quantitative estimations by plate counts of propagules of the bean root rot Fusarium in field soils. Phytopathology 1962, 52, 567–572. [Google Scholar]
- O’Donnell, K.; Sutton, D.A.; Rinaldi, M.G.; Gueidan, C.; Crous, P.W.; Geiser, D.M. Novel multilocus sequence typing scheme reveals high genetic diversity of human pathogenic members of the Fusarium incarnatum-F. equiseti and F. chlamydosporum species complexes within the United States. J. Clin. Microbiol. 2009, 47, 3851–3861. [Google Scholar] [CrossRef]
- Bougnoux, M.E.; Tavanti, A.; Bouchier, C.; Gow, N.A.R.; Magnier, A.; Davidson, A.D.; Maiden, M.C.J.; Enfert, C.; Odds, F.C. Collaborative consensus for optimized multilocus sequence typing of Candida albicans. J. Clin. Microbiol. 2003, 41, 5265–5266. [Google Scholar] [CrossRef]
- Odds, F.C.; Jacobsen, M.D. Multilocus sequence typing of pathogenic Candida species. Eukaryot. Cell 2008, 7, 1075–1084. [Google Scholar] [CrossRef]
- Bain, J.M.; Tavanti, A.; Davidson, A.D.; Jacobsen, M.D.; Shaw, D.; Gow, N.A.R.; Odds, F.C. Multilocus sequence typing of the pathogenic fungus Aspergillus fumigatus. J. Clin. Microbiol. 2007, 45, 1469–1477. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Source 1 | Species | MLST Type 2 | GenBank Accession Number | |
|---|---|---|---|---|---|
| TEF1-α | RPB2 | ||||
| Clinical Isolates | |||||
| Fp667 | Eye | F. petroliphilum | 1-b | MN178239 | MN263125 |
| Fk620 | Skin | F. keratoplasticum | 2-a | MN178238 | MN263124 |
| Fk2781 | Nail | F. keratoplasticum | 2-a | MN178234 | MN263120 |
| Fk2309 | Nail | F. keratoplasticum | 2-f | MN178231 | MN263117 |
| Fk553 | Skin | F. keratoplasticum | 2-h | MN178237 | MN263123 |
| Fk2353 | Nail | F. keratoplasticum | 2-h | MN178232 | MN263118 |
| Fk994 | Nail | F. keratoplasticum | 2-h | MN178240 | MN263126 |
| Fk0168 | Blood | F. keratoplasticum | 2-h | MN178228 | MN263114 |
| Fk2622 | Nail | F. keratoplasticum | 2-h | MN178233 | MN263119 |
| Fk1049 * | Nail | F. keratoplasticum | 2-h | MN178241 | MN263127 |
| Fk1931 * | Nail | F. keratoplasticum | 2-k | MN178230 | MN263116 |
| Fk1930 * | Nail | F. keratoplasticum | 2-k | MN178229 | MN263115 |
| Ff0020 * | Eye | F. falciforme | 3+4-k | MN178227 | MN263113 |
| Fstr541 | Blood | F. striatum | 21 | MN178236 | MN263122 |
| Fs263 * | Eye | F. suttonianum | 20-c | MN178235 | MN263121 |
| Environmental isolates | |||||
| FkDI17 | Grass | F. keratoplasticum | 2-a | MN178221 | MN263107 |
| FkDir61 | Grass | F. keratoplasticum | 2-a | MN178225 | MN263111 |
| Ff4225 | Tobacco | F. falciforme | 3+4-k | MN178212 | MN263098 |
| Ff4290 | Straw compost | F. falciforme | 3+4-k | MN178215 | MN263101 |
| Ff4324 ** | Soil | F. falciforme | 3+4-k | MN178216 | MN263102 |
| Ff4325 | Honeydew | F. falciforme | 3+4-k | MN178217 | MN263103 |
| FwgDE4 ** | Soil | F. waltergamsii | 7-b | MN178218 | MN263104 |
| FspDE40 ** | Soil | Fusarium sp. | 12-a | MN178220 | MN263106 |
| Fs3769 | Coconut tree | F. suttonianum | 20-c | MN178207 | MN263093 |
| Fs3784 | Mangrove | F. suttonianum | 20-c | MN178209 | MN263094 |
| Fs3873 | Grass | F. suttonianum | 20-c | MN178208 | MN263095 |
| Fs3924 | Sugarcane | F. suttonianum | 20-c | MN178210 | MN263096 |
| Fs4279 | Dragon fruit | F. suttonianum | 20-c | MN178214 | MN263100 |
| FcDir16 ** | Soil | F. cyanescens | 27-a | MN178223 | MN263109 |
| FcDir23 ** | Soil | F. cyanescens | 27-a | MN178224 | MN263110 |
| Isolate 1 | Mycelial Growth on PDA at 28 °C (Day-9) | Length and Width of Macroconidia (µm) 2,3,4 | |||||
|---|---|---|---|---|---|---|---|
| Top View | Viewed from Underneath | Colony Diameter (mm) | 2 and 3 Septa | 4 and 5 Septa | |||
| Length | Width | Length | Width | ||||
| CLINICAL ISOLATES (10) | |||||||
| Fp667 | white, brown | brown, yellow | 55 | 27.10 ± 3.65 | 2.79 ± 0.41 | 30.61 ± 2.68 | 2.99 ± 0.39 |
| Fk620 | White | white | 70 | none | none | none | none |
| Fk2781 | white brownish | yellow | 66 | 23.68 ± 2.95 | 3.05 ± 0.52 | none | none |
| Fk2309 | white with purple stripes | white | 55 | 21.37 ± 2.56 | 3.04 ± 0.47 | none | none |
| Fk553 | White | yellow | 66 | none | none | none | none |
| Fk2353 | white with purplish dark spots | white | 60 | 20.85 ± 1.93 | 3.02 ± 0.46 | none | none |
| Fk994 | white, brown | orange | 63 | 21.71 ± 3.22 | 2.91 ± 0.49 | none | none |
| Fk0168 | White | yellow | 68 | none | none | none | none |
| Fk2622 | White | white | 58 | none | none | none | none |
| Fstr541 | dark brown, white | dark brown, yellow | 65 | 26.63 ± 3.47 | 2.44 ± 0.47 | 31.49 ± 9.90 | 2.81 ± 0.49 |
| ENVIRONMENTAL ISOLATES (15) | |||||||
| FkDI17 | White | yellow | 78 | 24.56 ± 1.92 | 2.95 ± 0.50 | none | none |
| FkDir61 | white, dark brown, yellow, green | dark brown, orange, yellow | 46 | none | none | none | none |
| Ff4225 | white, dark brown | dark brown, orange, yellow | 78 | 21.50 ± 1.64 | 3.37 ± 0.35 | none | none |
| Ff4290 | White | Yellow | 76 | 25.51 ± 3.31 | 3.53 ± 0.53 | none | none |
| Ff4324 | white, purple | white, purple | 78 | 25.82 ± 2.00 | 3.27 ± 0.51 | none | none |
| Ff4325 | white, brown | Brown | 70 | 30.09 ± 2.71 | 3.07 ± 0.39 | none | none |
| FwgDE4 | white, brown | brown, yellow | 75 | 30.68 ± 3.51 | 2.95 ± 0.52 | 40.56 ± 2.79 | 2.92 ± 0.45 |
| FspDE40 | white, dark brown | dark brown, orange | 49 | 40.71 ± 2.15 | 2.98 ± 0.49 | 47.65 ± 4.27 | 2.94 ± 0.43 |
| Fs3769 | White | yellow | 80 | 24.83 ± 3.04 | 3.17 ± 0.42 | 41.02 ± 2.91 | 3.29 ± 0.41 |
| Fs3784 | dark brown, white | dark brown, yellow | 85 | none | none | none | none |
| Fs3873 | White | white | 80 | 33.18 ± 2.14 | 3.30 ± 0.54 | 41.32 ± 2.43 | 3.07 ± 0.52 |
| Fs3924 | White | yellow | 85 | 25.29 ± 5.02 | 3.31 ± 0.39 | none | none |
| Fs4279 | White | yellow | 73 | none | none | none | none |
| FcDir16 | white, dark brown | dark brown, orange, yellow | 63 | 41.56 ± 2.83 | 2.86 ± 0.41 | 50.76 ± 3.86 | 2.74 ± 0.39 |
| FcDir23 | White | white | 63 | 29.85 ± 4.89 | 3.02 ± 0.51 | 36.71 ± 1.37 | 2.95 ± 0.36 |
| Species | Macroconidia 1 | |
|---|---|---|
| Length (µm) | Width (µm) | |
| F. keratoplasticum (n = 5) 2 | 22.43 ± 1.60 | 2.99 ± 0.06 |
| F. falciforme (n = 4) | 25.70 ± 3.51 | 3.32 ± 0.19 |
| F. suttonianum (n = 3) 3 | 27.77 ± 4.69 | 3.26 ± 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
James, J.E.; Santhanam, J.; Zakaria, L.; Mamat Rusli, N.; Abu Bakar, M.; Suetrong, S.; Sakayaroj, J.; Abdul Razak, M.F.; Lamping, E.; Cannon, R.D. Morphology, Phenotype, and Molecular Identification of Clinical and Environmental Fusarium solani Species Complex Isolates from Malaysia. J. Fungi 2022, 8, 845. https://doi.org/10.3390/jof8080845
James JE, Santhanam J, Zakaria L, Mamat Rusli N, Abu Bakar M, Suetrong S, Sakayaroj J, Abdul Razak MF, Lamping E, Cannon RD. Morphology, Phenotype, and Molecular Identification of Clinical and Environmental Fusarium solani Species Complex Isolates from Malaysia. Journal of Fungi. 2022; 8(8):845. https://doi.org/10.3390/jof8080845
Chicago/Turabian StyleJames, Jasper E., Jacinta Santhanam, Latiffah Zakaria, Nuraini Mamat Rusli, Mariahyati Abu Bakar, Satinee Suetrong, Jariya Sakayaroj, Mohd Fuat Abdul Razak, Erwin Lamping, and Richard D. Cannon. 2022. "Morphology, Phenotype, and Molecular Identification of Clinical and Environmental Fusarium solani Species Complex Isolates from Malaysia" Journal of Fungi 8, no. 8: 845. https://doi.org/10.3390/jof8080845