
Exploring the Diversity and Aromatic Hydrocarbon Degrading Potential of Epiphytic Fungi on Hornbeams from Chronically Polluted Areas

 , , , and
, , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling of Hornbeam Leaves from Three Differently Polluted Areas in Poland
2.2. Isolation of Biomass from Hornbeam Phyllosphere
2.3. Total gDNA Extraction from Leaf Wash Suspensions
2.4. Community Fingerprint Analyses
2.5. Shotgun Metagenomic Sequencing
2.6. Isolation of Culturable Epiphytic Fungi
2.7. Improving Isolation of Culturable Epiphytic Fungi via Enrichment Cultures
2.8. Genotypic Characterization of the Isolates
2.9. Laccase and Peroxidase Enzyme Assays
2.10. Fungal Hydroxyl Radical Production Assay
2.11. MAH and PAH Tolerance Test
2.12. Naphthalene Degradation Assay
2.13. Analyses of Naphthalene Concentrations Using GC-MS
2.14. Statistical Analyses
2.15. NCBI Accession Numbers
3. Results
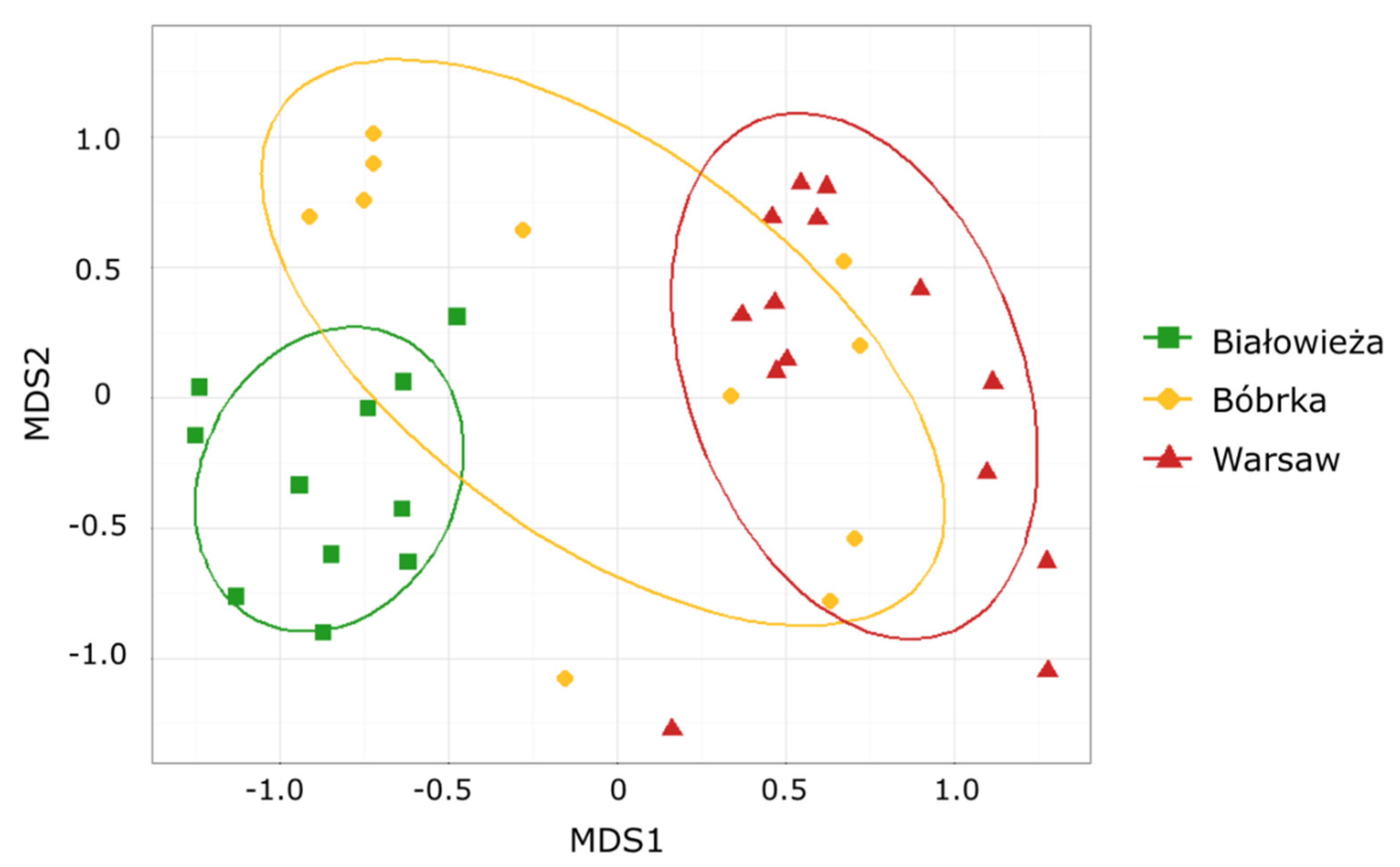
3.1. Comparison of the Hornbeam Epiphytic Fungal Communities via Community Fingerprint
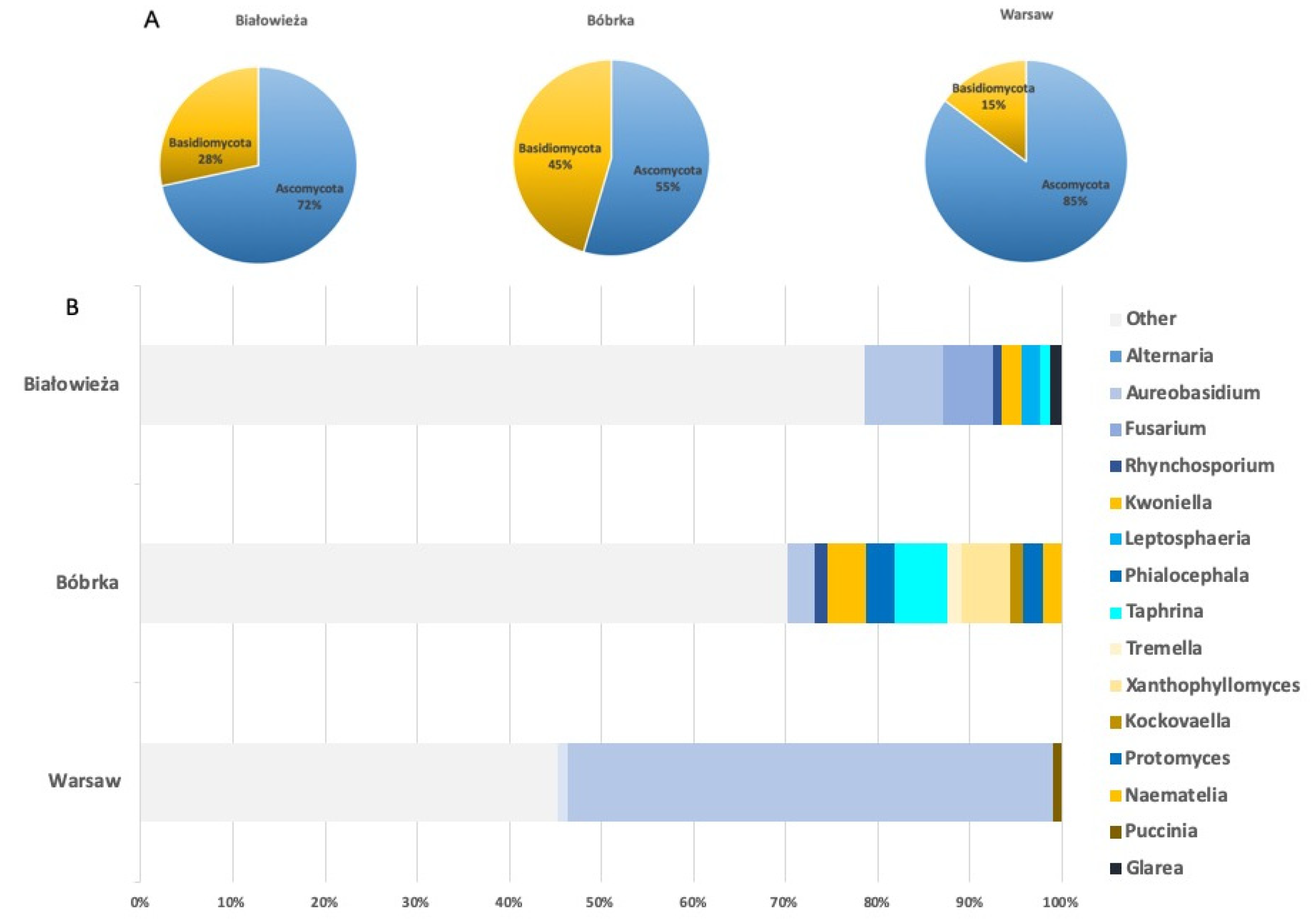
3.2. Shotgun Metagenomic Sequencing of Hornbeam Epiphytic Communities
3.3. Fungal Cultivable Isolates from the Hornbeam Leaves
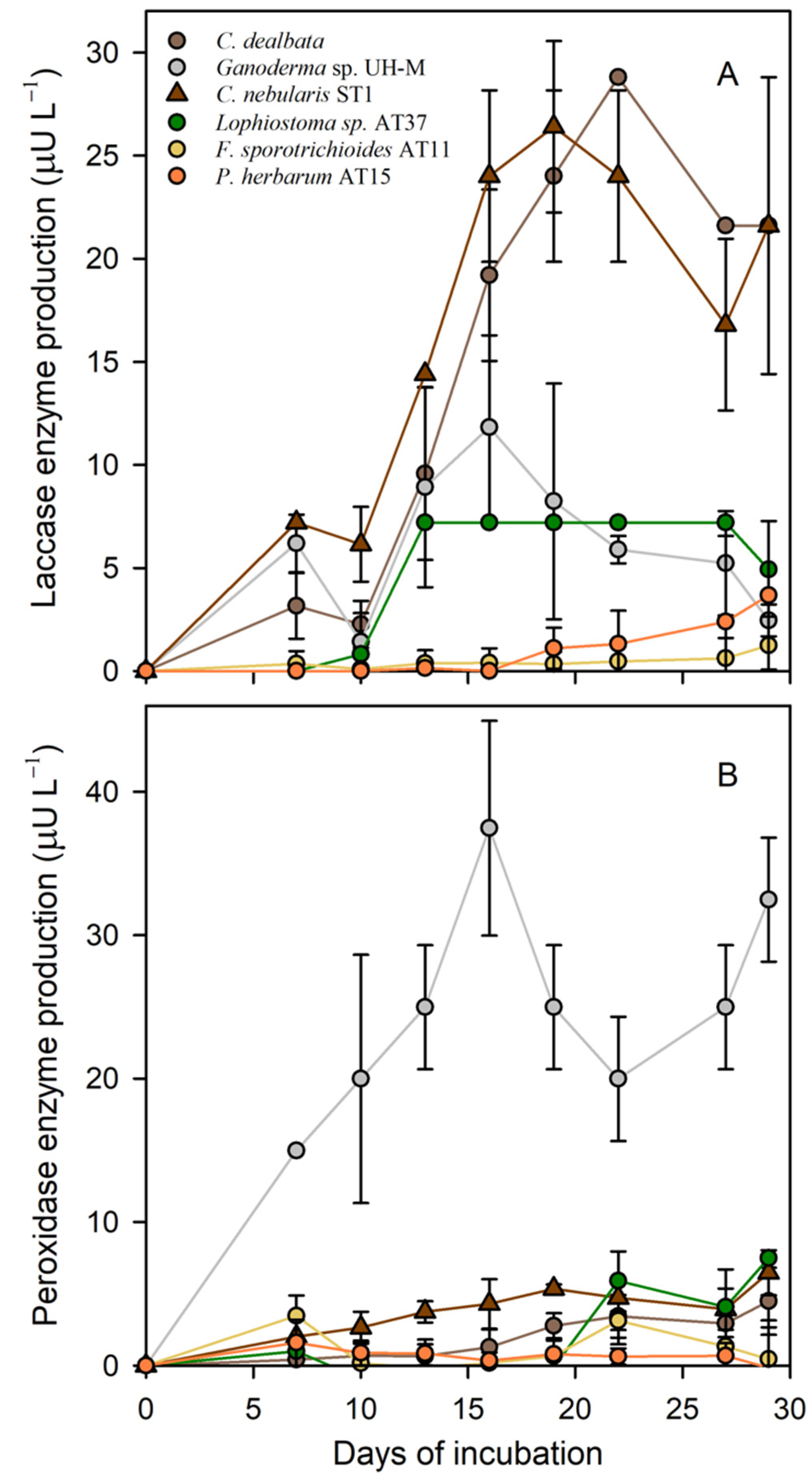
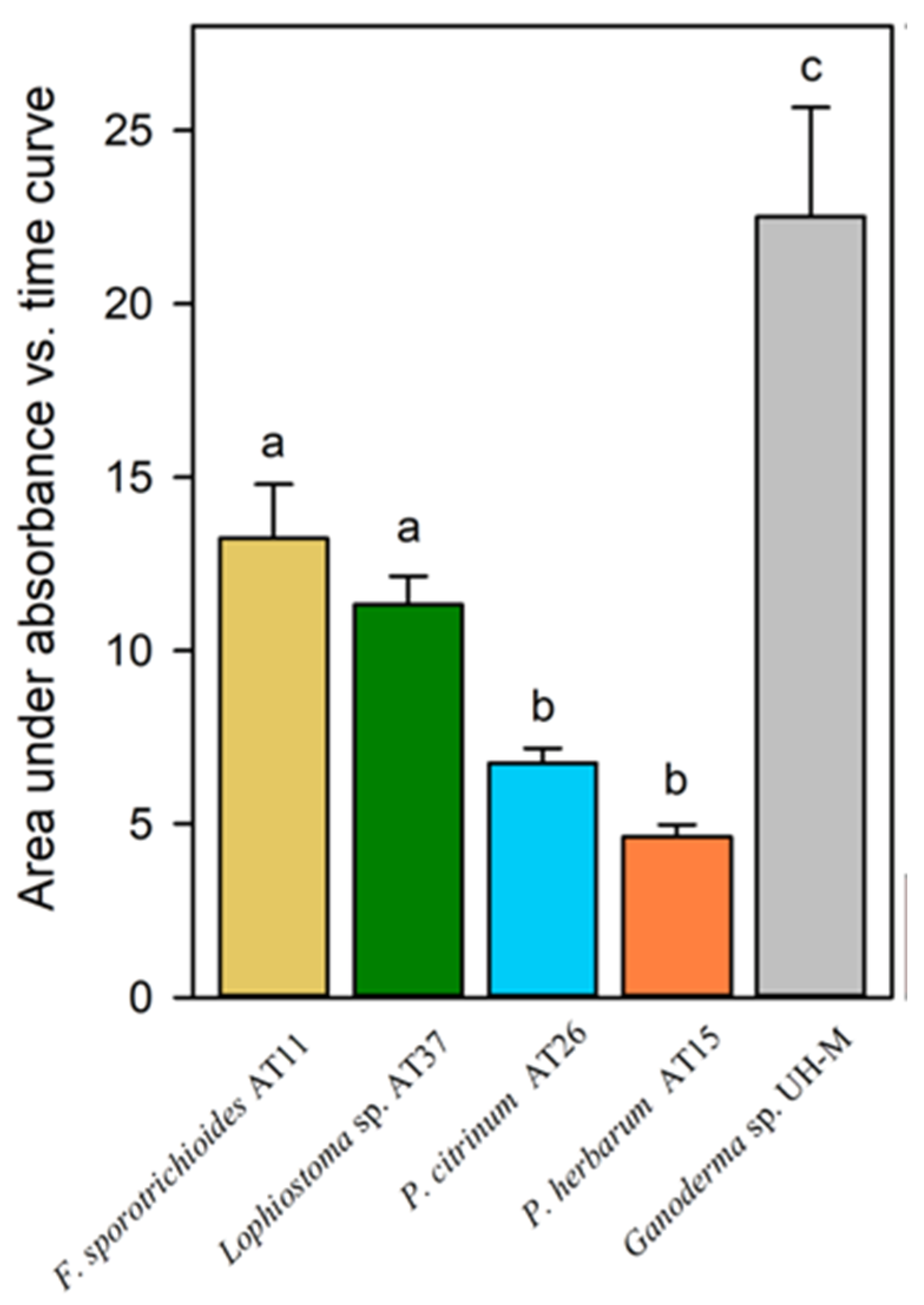
3.4. Laccase and Peroxidase Activities
3.5. Hydroxyl Radical Production
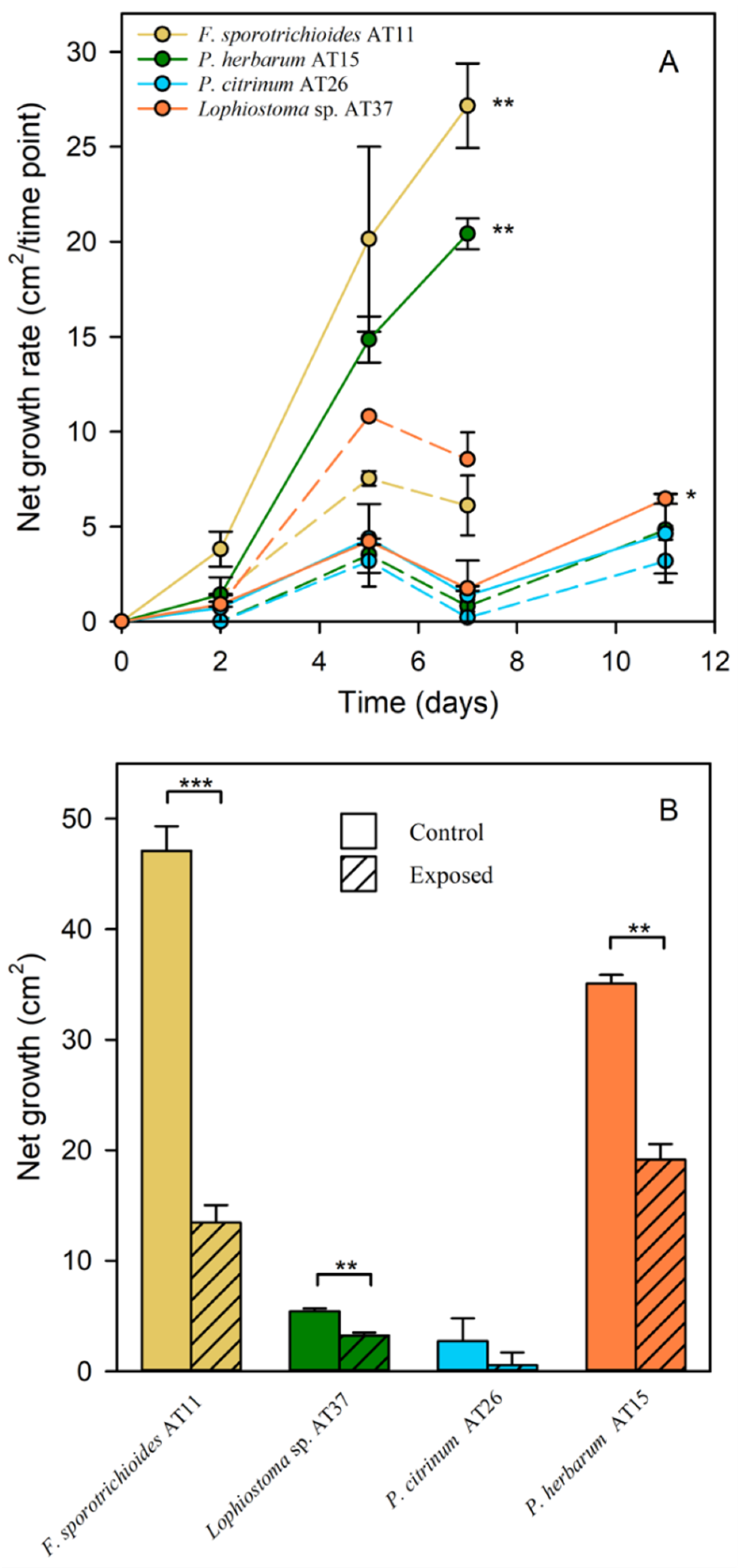
3.6. MAH and PAH Tolerance
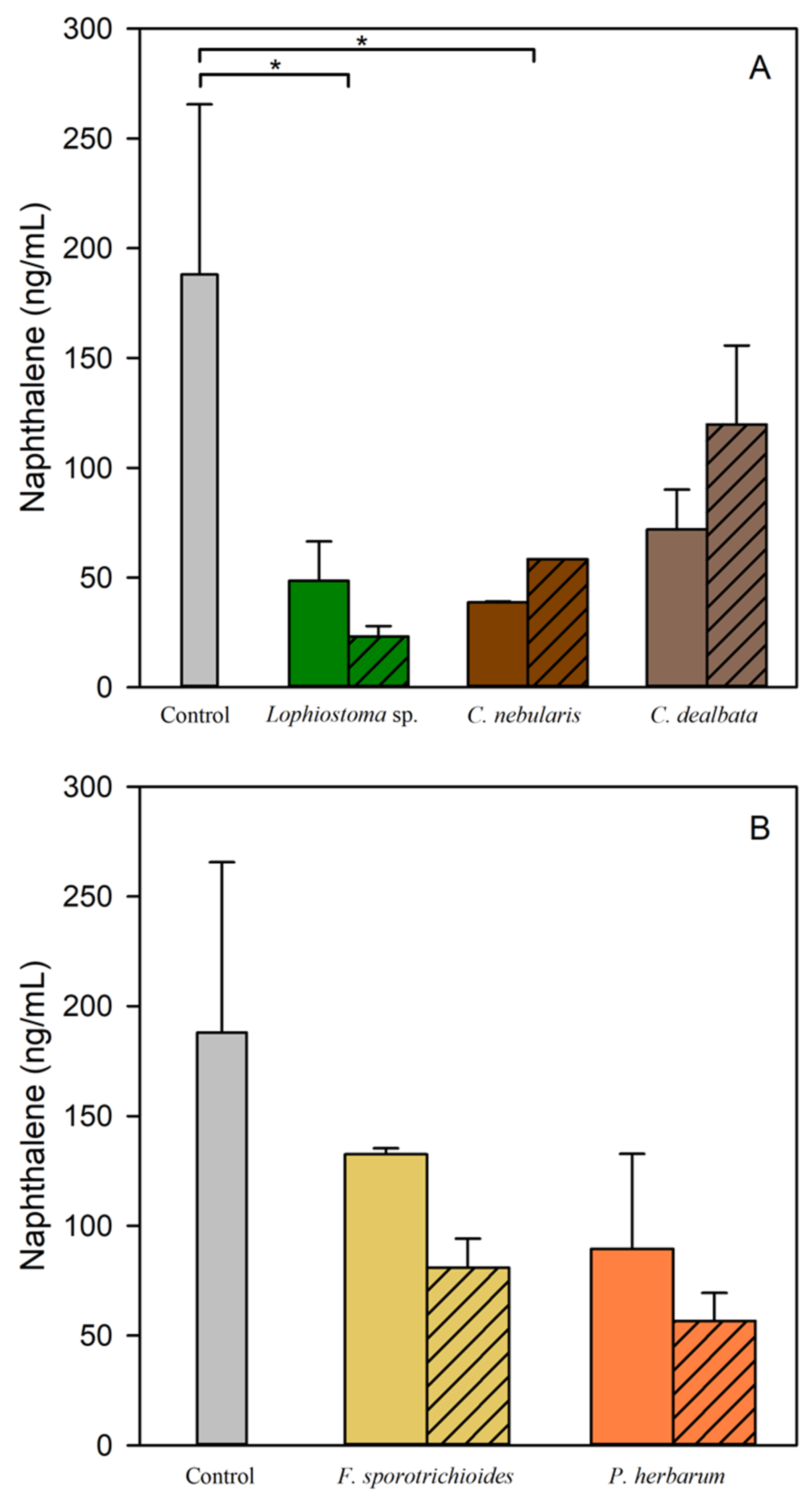
3.7. Naphthalene Degradation
4. Discussion
4.1. Hornbeam Epiphytic Fungal Communities
4.2. Catabolic Enzymes Involved in the Degradation of AHs
4.3. Tolerance to MAH and PAH and Napthalene Degradation Potential
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- WHO. World Health Organization. Available online: https://www.who.int/news-room/detail/02-05-2018-9-out-of-10-people-worldwide-breathe-polluted-air-but-more-countries-are-taking-action (accessed on 1 August 2021).
- Wei, X.; Lyu, S.; Yu, Y.; Wang, Z.; Liu, H.; Pan, D.; Chen, J. Phylloremediation of Air Pollutants: Exploiting the Potential of Plant Leaves and Leaf-Associated Microbes. Front. Plant. Sci. 2017, 8, 1318. [Google Scholar] [CrossRef] [PubMed]
- Dzierzanowski, K.; Popek, R.; Gawronska, H.; Saebo, A.; Gawronski, S.W. Deposition of Particulate Matter of Different Size Fractions on Leaf Surfaces and in Waxes of Urban Forest Species. Int. J. Phytoremediat. 2011, 13, 1037–1046. [Google Scholar] [CrossRef]
- Vorholt, J.A. Microbial life in the phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Weyens, N.; Beckers, B.; Schellingen, K.; Ceulemans, R.; van der Lelie, D.; Newman, L.; Taghavi, S.; Carleer, R.; Vangronsveld, J. The Potential of the Ni-Resistant TCE-Degrading Pseudomonas putida W619-TCE to Reduce Phytotoxicity and Improve Phytoremediation Efficiency of Poplar Cuttings on A Ni-TCE Co-Contamination. Int. J. Phytoremediat. 2015, 17, 40–48. [Google Scholar] [CrossRef]
- Sivakumar, N.; Sathishkumar, R.; Selvakumar, G.; Shyamkumar, R.; Arjunekumar, K. Phyllospheric Microbiomes: Diversity, Ecological Significance, and Biotechnological Applications. In Plant Microbiomes for Sustainable Agriculture; Yadav, A.N., Singh, J., Rastegari, A.A., Yadav, N., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 113–172. [Google Scholar] [CrossRef]
- Leveau, J.H. A brief from the leaf: Latest research to inform our understanding of the phyllosphere microbiome. Curr. Opin. Microbiol. 2019, 49, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Gomes, T.; Pereira, J.A.; Benhadi, J.; Lino-Neto, T.; Baptista, P. Endophytic and Epiphytic Phyllosphere Fungal Communities Are Shaped by Different Environmental Factors in a Mediterranean Ecosystem. Microb. Ecol. 2018, 76, 668–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirschner, R. Fungi on the leaf—A contribution towards a review of phyllosphere microbiology from the mycological perspective. Ecol. Ser. 2018, 34, 433–448. [Google Scholar]
- Davey, M.; Nybakken, L.; Kauserud, H.; Ohlson, M. Fungal biomass associated with the phyllosphere of bryophytes and vascular plants. Mycol. Res. 2009, 113, 1254–1260. [Google Scholar] [CrossRef]
- Jia, T.; Yao, Y.; Guo, T.; Wang, R.; Chai, B. Effects of Plant and Soil Characteristics on Phyllosphere and Rhizosphere Fungal Communities During Plant Development in a Copper Tailings Dam. Front. Microbiol. 2020, 11, 556002. [Google Scholar] [CrossRef]
- Muller, T.; Ruppel, S. Progress in cultivation-independent phyllosphere microbiology. FEMS Microbiol. Ecol. 2014, 87, 2–17. [Google Scholar] [CrossRef]
- De Silva, S.; Aluwihare, S.; Chandimala, J.; Jayasooriya, R.; Undugoda, R. Trends of Floods in Sri Lanka. In Proceedings of the 36th Iahr World Congress, The Hague, The Netherlands, 28 June–3 July 2015; pp. 4985–4991. [Google Scholar]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current State of Knowledge in Microbial Degradation of Polycyclic Aromatic Hydrocarbons (PAHs): A Review. Front. Microbiol. 2016, 7, 1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadri, T.; Rouissi, T.; Brar, S.K.; Cledon, M.; Sarma, S.; Verma, M. Biodegradation of polycyclic aromatic hydrocarbons (PAHs) by fungal enzymes: A review. J. Environ. Sci. 2017, 51, 52–74. [Google Scholar] [CrossRef]
- Irga, P.J.; Pettit, T.J.; Torpy, F.R. The phytoremediation of indoor air pollution: A review on the technology development from the potted plant through to functional green wall biofilters. Rev. Environ. Sci. Bio. 2018, 17, 395–415. [Google Scholar] [CrossRef]
- Molloy, S. Phenol and the phyllosphere. Nat. Rev. Microbiol. 2006, 4, 880–881. [Google Scholar] [CrossRef]
- Bienkowska, M.; Faszcza, L.; Wolyniec, L. Movement to Defend the Bialowieza-The Problem of the Bialowieza Forest Protection as an Example of a Values Conflict. In Borderology: Cross-Disciplinary Insights from the Border Zone; Springer: Cham, Switzerland, 2019; pp. 31–40. [Google Scholar] [CrossRef]
- Olejarz, B. The Museum of the Oil and Gas Industry was owned by Ignacy Lukasiewicz in Bobrce. Przem. Chem. 2013, 92, 15–18. [Google Scholar]
- Imperato, V.; Kowalkowski, L.; Portillo-Estrada, M.; Gawronski, S.W.; Vangronsveld, J.; Thijs, S. Characterisation of the Carpinus betulus L. Phyllomicrobiome in Urban and Forest Areas. Front. Microbiol. 2019, 10, 1110. [Google Scholar] [CrossRef] [Green Version]
- Stevens, V. The Structure of Phyllospheric Microbial Communities and Their Role in Phytoremediation of Air Pollution. UHasselt, Belgium. 2016. Available online: https://www.semanticscholar.org/paper/The-structure-of-phyllospheric-microbial-and-their-Stevens/cb5ed75d9d9c5bfad3e20d823e8ed26c35d4902a (accessed on 1 November 2021).
- Cardinale, M.; Brusetti, L.; Quatrini, P.; Borin, S.; Puglia, A.M.; Rizzi, A.; Zanardini, E.; Sorlini, C.; Corselli, C.; Daffonchio, D. Comparison of different primer sets for use in automated ribosomal intergenic spacer analysis of complex bacterial communities. Appl. Environ. Microbiol. 2004, 70, 6147–6156. [Google Scholar] [CrossRef] [Green Version]
- Ranjard, L.; Poly, F.; Lata, J.C.; Mougel, C.; Thioulouse, J.; Nazaret, S. Characterization of bacterial and fungal soil communities by automated ribosomal intergenic spacer analysis fingerprints: Biological and methodological variability. Appl. Environ. Microb. 2001, 67, 4479–4487. [Google Scholar] [CrossRef] [Green Version]
- Sequerra, J.; Marmeisse, R.; Valla, G.; Normand, P.; Capellano, A.; Moiroud, A. Taxonomic position and intraspecific variability of the nodule forming Penicillium nodositatum inferred from RFLP analysis of the ribosomal intergenic spacer and Random Amplified Polymorphic DNA. Mycol. Res. 1997, 101, 465–472. [Google Scholar] [CrossRef]
- Gastauer, M.; Vera, M.P.O.; de Souza, K.P.; Pires, E.S.; Alves, R.; Caldeira, C.F.; Ramos, S.J.; Oliveira, G. A metagenomic survey of soil microbial communities along a rehabilitation chronosequence after iron ore mining. Sci. Data 2019, 6, 190008. [Google Scholar] [CrossRef]
- Abraham, B.S.; Caglayan, D.; Carrillo, N.V.; Chapman, M.C.; Hagan, C.T.; Hansen, S.T.; Jeanty, R.O.; Klimczak, A.A.; Klingler, M.J.; Kutcher, T.P.; et al. Shotgun metagenomic analysis of microbial communities from the Loxahatchee nature preserve in the Florida Everglades. Environ. Microbiome 2020, 15, 2. [Google Scholar] [CrossRef] [Green Version]
- Salipante, S.J.; Sengupta, D.J.; Rosenthal, C.; Costa, G.; Spangler, J.; Sims, E.H.; Jacobs, M.A.; Miller, S.I.; Hoogestraat, D.R.; Cookson, B.T.; et al. Rapid 16S rRNA Next-Generation Sequencing of Polymicrobial Clinical Samples for Diagnosis of Complex Bacterial Infections. PLoS ONE 2013, 8, e65226. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Iliev, I.D.; Brown, J.; Underhill, D.M.; Funari, V.A. Mycobiome: Approaches to analysis of intestinal fungi. J. Immunol. Methods 2015, 421, 112–121. [Google Scholar] [CrossRef] [Green Version]
- Menzel, P.; Ng, K.L.; Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 2016, 7, 11257. [Google Scholar] [CrossRef] [Green Version]
- Black, W.D. A comparison of several media types and basic techniques used to assess outdoor airborne fungi in Melbourne, Australia. PLoS ONE 2020, 15, e0238901. [Google Scholar] [CrossRef]
- Wu, P.-C.; Su, H.-J.J.; Ho, H.-M. A Comparison of Sampling Media for Environmental Viable Fungi Collected in a Hospital Environment. Environ. Res. 2000, 82, 253–257. [Google Scholar] [CrossRef]
- Abildgren, M.P.; Lund, F.; Thrane, U.; Elmholt, S. Czapek-Dox Agar Containing Iprodione and Dicloran as a Selective Medium for the Isolation of Fusarium Species. Lett. Appl. Microbiol. 1987, 5, 83–86. [Google Scholar] [CrossRef]
- Luria, S.E.; Burrous, J.W. Hybridization between Escherichia-Coli and Shigella. J. Bacteriol. 1957, 74, 461–476. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Bose, C.; Ojha, N.; Das, N.; Das, J.; Pal, M.; Khurana, S. Evolution of bacterial and fungal growth media. Bioinformation 2015, 11, 182–184. [Google Scholar] [CrossRef]
- Bushnell, L.D.; Haas, H.F. The Utilization of Certain Hydrocarbons by Microorganisms. J. Bacteriol. 1941, 41, 653–673. [Google Scholar] [CrossRef] [Green Version]
- Ingestad, T.; Kahr, M. Nutrition and Growth of Coniferous Seedlings at Varied Relative Nitrogen Addition Rate. Physiol. Plantarum. 1985, 65, 109–116. [Google Scholar] [CrossRef]
- Thorn, R.G.; Reddy, C.A.; Harris, D.; Paul, E.A. Isolation of saprophytic basidiomycetes from soil. Appl. Environ. Microb. 1996, 62, 4288–4292. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Ren, X.; Lei, Q.; Wang, L. Screening and Comparison of Lignin Degradation Microbial Consortia from Wooden Antiques. Molecules 2021, 26, 2862. [Google Scholar] [CrossRef]
- Balseiro-Romero, M.; Gkorezis, P.; Kidd, P.S.; Van Hamme, J.; Weyens, N.; Monterroso, C.; Vangronsveld, J. Characterization and degradation potential of diesel-degrading bacterial strains for application in bioremediation. Int. J. Phytoremediat. 2017, 19, 955–963. [Google Scholar] [CrossRef] [Green Version]
- Op De Beeck, M.; Lievens, B.; Busschaert, P.; Declerck, S.; Vangronsveld, J.; Colpaert, J.V. Comparison and validation of some ITS primer pairs useful for fungal metabarcoding studies. PLoS ONE 2014, 9, e97629. [Google Scholar] [CrossRef] [Green Version]
- Orgiazzi, A.; Lumini, E.; Nilsson, R.H.; Girlanda, M.; Vizzini, A.; Bonfante, P.; Bianciotto, V. Unravelling soil fungal communities from different Mediterranean land-use backgrounds. PLoS ONE 2012, 7, e34847. [Google Scholar] [CrossRef]
- VanderMolen, K.M.; Raja, H.A.; El-Elimat, T.; Oberlies, N.H. Evaluation of culture media for the production of secondary metabolites in a natural products screening program. Amb. Express 2013, 3, 71. [Google Scholar] [CrossRef] [Green Version]
- Torres-Farrada, G.; Leon, A.M.M.; Rineau, F.; Alonso, L.L.L.; Sanchez-Lopez, M.I.; Thijs, S.; Colpaert, J.; Ramos-Leal, M.; Guerra, G.; Vangronsveld, J. Diversity of Ligninolytic Enzymes and Their Genes in Strains of the Genus Ganoderma: Applicable for Biodegradation of Xenobiotic Compounds? Front. Microbiol. 2017, 8, 898. [Google Scholar] [CrossRef]
- Teerapatsakul, C.; Abe, N.; Bucke, C.; Kongkathip, N.; Jareonkitmongkol, S.; Chitradon, L. Novel laccases of Ganoderma sp KU-Alk4, regulated by different glucose concentration in alkaline media. World J. Microb. Biot. 2007, 23, 1559–1567. [Google Scholar] [CrossRef]
- Claiborne, A.; Malinowski, D.P.; Fridovich, I. Purification and Characterization of Hydroperoxidase-Ii of Escherichia-Coli-B. J. Biol. Chem. 1979, 254, 1664–1668. [Google Scholar] [CrossRef]
- Page, S.E.; Arnold, W.A.; McNeill, K. Terephthalate as a probe for photochemically generated hydroxyl radical. J. Environ. Monitor. 2010, 12, 1658–1665. [Google Scholar] [CrossRef]
- Jambon, I.; Thijs, S.; Torres-Farradá, G.; Rineau, F.; Weyens, N.; Carleer, R.; Samyn, P.; Vangronsveld, J. Fenton-Mediated Biodegradation of Chlorendic Acid—A Highly Chlorinated Organic Pollutant—By Fungi Isolated From a Polluted Site. Front. Microbiol. 2019, 10, 1892. [Google Scholar] [CrossRef] [Green Version]
- Jia, C.R.; Batterman, S. A Critical Review of Naphthalene Sources and Exposures Relevant to Indoor and Outdoor Air. Int. J. Environ. Res. Public Health 2010, 7, 2903–2939. [Google Scholar] [CrossRef] [Green Version]
- Sekar, A.; Varghese, G.K.; Varma, M.K.R. Analysis of benzene air quality standards, monitoring methods and concentrations in indoor and outdoor environment. Heliyon 2019, 5, e02918. [Google Scholar] [CrossRef] [Green Version]
- Batterman, S.; Chin, J.Y.; Jia, C.; Godwin, C.; Parker, E.; Robins, T.; Max, P.; Lewis, T. Sources, concentrations, and risks of naphthalene in indoor and outdoor air. Indoor Air 2012, 22, 266–278. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Shafy, H.I.; Mansour, M.S.M. A review on polycyclic aromatic hydrocarbons: Source, environmental impact, effect on human health and remediation. Egypt. J. Pet. 2016, 25, 107–123. [Google Scholar] [CrossRef] [Green Version]
- Lawal, A.T. Polycyclic aromatic hydrocarbons. A review. Cogent Environ. Sci. 2017, 3, 1339841. [Google Scholar] [CrossRef]
- Cervantes, C.; Gutierrezcorona, F. Copper Resistance Mechanisms in Bacteria and Fungi. FEMS Microbiol. Rev. 1994, 14, 121–137. [Google Scholar] [CrossRef]
- Tisserand, R.; Young, R. 6—The respiratory system. In Essential Oil Safety, 2nd ed.; Tisserand, R., Young, R., Eds.; Churchill Livingstone: St. Louis, MO, USA, 2014; pp. 99–110. [Google Scholar] [CrossRef]
- Steven, W.K.; Rebecca, C.M. Plant traits and taxonomy drive host associations in tropical phyllosphere fungal communities. Botany 2014, 92, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Kembel, S.W.; O’Connor, T.K.; Arnold, H.K.; Hubbell, S.P.; Wright, S.J.; Green, J.L. Relationships between phyllosphere bacterial communities and plant functional traits in a neotropical forest. Proc. Natl. Acad. Sci. USA 2014, 111, 13715. [Google Scholar] [CrossRef] [Green Version]
- Hawksworth, D.L. Fungal diversity and its implications for genetic resource collections. Stud. Mycol. 2004, 50, 9–17. [Google Scholar]
- Janakiev, T.; Dimkić, I.; Unković, N.; Ljaljević Grbić, M.; Opsenica, D.; Gašić, U.; Stanković, S.; Berić, T. Phyllosphere Fungal Communities of Plum and Antifungal Activity of Indigenous Phenazine-Producing Pseudomonas synxantha Against Monilinia laxa. Front. Microbiol. 2019, 10, 2287. [Google Scholar] [CrossRef] [Green Version]
- Laforest-Lapointe, I.; Messier, C.; Kembel, S.W.; Brodie, E.L. Tree Leaf Bacterial Community Structure and Diversity Differ along a Gradient of Urban Intensity. mSystems 2017, 2, e00087-17. [Google Scholar] [CrossRef] [Green Version]
- Nock, C.A.; Paquette, A.; Follett, M.; Nowak, D.J.; Messier, C. Effects of Urbanization on Tree Species Functional Diversity in Eastern North America. Ecosystems 2013, 16, 1487–1497. [Google Scholar] [CrossRef]
- Oke, T.R. City size and the urban heat island. Atmos. Environ. (1967) 1973, 7, 769–779. [Google Scholar] [CrossRef]
- Jumpponen, A.; Jones, K.L.; David Mattox, J.; Yaege, C. Massively parallel 454-sequencing of fungal communities in Quercus spp. ectomycorrhizas indicates seasonal dynamics in urban and rural sites. Mol. Ecol. 2010, 19 (Suppl. 1), 41–53. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, V.L.; Monteiro, A.D.; Braga, D.T.; Santoro, M.M. Phenol degradation by Aureobasidium pullulans FE13 isolated from industrial effluents. J. Hazard. Mater. 2009, 161, 1413–1420. [Google Scholar] [CrossRef]
- Schoeman, M.W.; Dickinson, D.J. Aureobasidium pullulans can utilize simple aromatic compounds as a sole source of carbon in liquid culture. Lett. Appl. Microbiol. 1996, 22, 129–131. [Google Scholar] [CrossRef]
- Penselin, D.; Munsterkotter, M.; Kirsten, S.; Felder, M.; Taudien, S.; Platzer, M.; Ashelford, K.; Paskiewicz, K.H.; Harrison, R.J.; Hughes, D.J.; et al. Comparative genomics to explore phylogenetic relationship, cryptic sexual potential and host specificity of Rhynchosporium species on grasses. BMC Genom. 2016, 17, 953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brighigna, L.; Gori, A.; Gonnelli, S.; Favilli, F. The influence of air pollution on the phyllosphere microflora composition of Tillandsia leaves (Bromeliaceae). Rev. Biol. Trop. 2000, 48, 511–517. [Google Scholar]
- Ljs, U.; Kannangara, S.; Sirisena, D. Aromatic Hydrocarbon Degrading Fungi Inhabiting the Phyllosphere of Ornamental Plants on Roadsides of Urban Areas in Sri Lanka. J. Bioremediat. Biodegrad. 2016, 7, 2155–6199. [Google Scholar] [CrossRef]
- Hong, J.W.; Park, J.Y.; Gadd, G.M. Pyrene degradation and copper and zinc uptake by Fusarium solani and Hypocrea lixii isolated from petrol station soil. J. Appl. Microbiol. 2010, 108, 2030–2040. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.W.; Dyer, B.J.; Noé, J. Hydrocarbon Utilization by Higher Marine Fungi from Diverse Habitats and Localities. Mycologia 1991, 83, 227–230. [Google Scholar] [CrossRef]
- Barnes, N.M.; Khodse, V.B.; Lotlikar, N.P.; Meena, R.M.; Damare, S.R. Bioremediation potential of hydrocarbon-utilizing fungi from select marine niches of India. 3 Biotech. 2018, 8, 21. [Google Scholar] [CrossRef]
- Hughes, K.A.; Bridge, P.; Clark, M.S. Tolerance of Antarctic soil fungi to hydrocarbons. Sci. Total Environ. 2007, 372, 539–548. [Google Scholar] [CrossRef]
- Janusz, G.; Pawlik, A.; Sulej, J.; Swiderska-Burek, U.; Jarosz-Wilkolazka, A.; Paszczynski, A. Lignin degradation: Microorganisms, enzymes involved, genomes analysis and evolution. FEMS Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef] [Green Version]
- Tuomela, M.; Hatakka, A. Oxidative Fungal Enzymes for Bioremediation. Compr. Biotechnol. 2011, 6, 183–196. [Google Scholar]
- Krueger, M.C.; Bergmann, M.; Schlosser, D. Widespread ability of fungi to drive quinone redox cycling for biodegradation. FEMS Microbiol. Lett. 2016, 363, fnw105. [Google Scholar] [CrossRef] [Green Version]
- Rineau, F.; Roth, D.; Shah, F.; Smits, M.; Johansson, T.; Canback, B.; Olsen, P.B.; Persson, P.; Grell, M.N.; Lindquist, E.; et al. The ectomycorrhizal fungus Paxillus involutus converts organic matter in plant litter using a trimmed brown-rot mechanism involving Fenton chemistry. Environ. Microbiol. 2012, 14, 1477–1487. [Google Scholar] [CrossRef] [Green Version]
- De Menezes, T.A.; Bispo, A.S.; Koblitz, M.G.; Vandenberghe, L.P.; Kamida, H.M.; Goes-Neto, A. Production of Basidiomata and Ligninolytic Enzymes by the Lingzhi or Reishi Medicinal Mushroom, Ganoderma lucidum (Agaricomycetes), in Licuri (Syagrus coronata) Wastes in Brazil. Int. J. Med. Mushrooms 2016, 18, 1141–1149. [Google Scholar] [CrossRef]
- Weyens, N.; Thijs, S.; Popek, R.; Witters, N.; Przybysz, A.; Espenshade, J.; Gawronska, H.; Vangronsveld, J.; Gawronski, S.W. The Role of Plant-Microbe Interactions and Their Exploitation for Phytoremediation of Air Pollutants. Int. J. Mol. Sci. 2015, 16, 25576–25604. [Google Scholar] [CrossRef] [Green Version]
- Lysenko, S.V.; Liakh, S.P. The protective role of pigments against UV rays in fungi isolated from the mesosphere. Mikrobiologiia 1977, 46, 867–877. [Google Scholar] [PubMed]
- Pagano, M.C.; Dhar, P.P. Fungal pigments: An overview. In Fungal Biomolecules: Sources, Applications and Recent Developments; Wiley: Hoboken, NJ, USA, 2015; Chapter 13; pp. 173–181. [Google Scholar] [CrossRef]
- Dsouza, G.C.; Sheriff, R.S.; Ullanat, V.; Shrikrishna, A.; Joshi, A.V.; Hiremath, L.; Entoori, K. Fungal biodegradation of low-density polyethylene using consortium of Aspergillus species under controlled conditions. Heliyon 2021, 7, e07008. [Google Scholar] [CrossRef] [PubMed]
- Kennes, C.; Veiga, M.C. Fungal biocatalysts in the biofiltration of VOC-polluted air. J. Biotechnol. 2004, 113, 305–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.; Veiga, M.; Kennes, C. Performance optimization of the fungal biodegradation of pinene in gas-phase biofilter. Process. Biochem. 2006, 41, 1722–1728. [Google Scholar] [CrossRef] [Green Version]
- Zeinali, M.; Vossoughi, M.; Ardestani, S.K. Naphthalene metabolism in Nocardia otitidiscaviarum strain TSH1, a moderately thermophilic microorganism. Chemosphere 2008, 72, 905–909. [Google Scholar] [CrossRef]
- Hadibarata, T.; Yusoff, A.R.M.; Aris, A.; Kristanti, R.A. Identification of naphthalene metabolism by white rot fungus Armillaria sp. F022. J. Environ. Sci. 2012, 24, 728–732. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungi | Bacteria [20] | |||
|---|---|---|---|---|
| Mean | SD | Mean | SD | |
| Bóbrka | 14,106.00 | 7097.73 | 340,110.50 | 64,229.54 |
| Białowieża | 4449.00 | 1011.79 | 426,475.33 | 77,314.96 |
| Warsaw | 5704.17 | 4734.63 | 336,557.33 | 62,557.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imperato, V.; Portillo-Estrada, M.; Saran, A.; Thoonen, A.; Kowalkowski, Ł.; Gawronski, S.W.; Rineau, F.; Vangronsveld, J.; Thijs, S. Exploring the Diversity and Aromatic Hydrocarbon Degrading Potential of Epiphytic Fungi on Hornbeams from Chronically Polluted Areas. J. Fungi 2021, 7, 972. https://doi.org/10.3390/jof7110972
Imperato V, Portillo-Estrada M, Saran A, Thoonen A, Kowalkowski Ł, Gawronski SW, Rineau F, Vangronsveld J, Thijs S. Exploring the Diversity and Aromatic Hydrocarbon Degrading Potential of Epiphytic Fungi on Hornbeams from Chronically Polluted Areas. Journal of Fungi. 2021; 7(11):972. https://doi.org/10.3390/jof7110972
Chicago/Turabian StyleImperato, Valeria, Miguel Portillo-Estrada, Anabel Saran, Anneleen Thoonen, Łukasz Kowalkowski, Stanislaw W. Gawronski, Francois Rineau, Jaco Vangronsveld, and Sofie Thijs. 2021. "Exploring the Diversity and Aromatic Hydrocarbon Degrading Potential of Epiphytic Fungi on Hornbeams from Chronically Polluted Areas" Journal of Fungi 7, no. 11: 972. https://doi.org/10.3390/jof7110972