Multicopper Oxidases in Saccharomyces cerevisiae and Human Pathogenic Fungi

1
Department of Microbiology, University of Szeged, Kozep fasor, 6726 Szeged, Hungary
2
Departments of Medicine and Microbiology and Immunology, Albert Einstein College of Medicine, New York, NY 10461, USA
3
MTA-SZTE Lendület Mycobiome Research Group, University of Szeged, 6726 Szeged, Hungary
*
Author to whom correspondence should be addressed.
J. Fungi 2020, 6(2), 56; https://doi.org/10.3390/jof6020056
Submission received: 30 March 2020
/
Revised: 20 April 2020
/
Accepted: 25 April 2020
/
Published: 27 April 2020
(This article belongs to the Special Issue Pathogenesis of Candidiasis)
{kind=link}
Abstract
:Multicopper oxidases (MCOs) are produced by microscopic and macroscopic fungal species and are involved in various physiological processes such as morphogenesis, lignin degradation, and defense mechanisms to stress inducing environmental conditions as well as fungal virulence. This review will summarize our current understanding regarding the functions of MCOs present in Saccharomyces cerevisiae and in different human fungal pathogens. Of the two main MCO groups, the first group of MCOs is involved in iron homoeostasis and the second includes laccases. This review will also discuss their role in the pathogenesis of human fungal pathogens.
1. Introduction
Multicopper oxidases (MCOs) are blue copper containing proteins and they generally have multiple copper atoms (1 to 6) per molecule [1,2]. Diverse enzymes belong to this family of proteins that include laccases, ferroxidases, ascorbate oxidase, bilirubin oxidase and laccase-like MCOs [3]. MCOs are produced by both prokaryotic as well as various eukaryotic organisms. MCOs consist of four enzyme families: laccases (EC 1.10.3.2), ascorbate oxidases (EC 1.10.3.3), ferroxidases (EC 1.16.3.1), and ceruloplasmin (EC 1.16.3.1) [4]. Detailed analyses of 350 different MCOs were used to classify them in 10 super families, which are: (A) basidiomycete laccases, (B) ascomycete laccases, (C) insect laccases, (D) fungal pigment MCOs, (E) fungal ferroxidases, (F) fungal and plant ascorbate oxidases, (G) plant laccase-like MCOs, (H) copper resistance proteins (CopA), (I) bilirubin oxidases, and (J) copper efflux (CueO) proteins [2]. All MCOs can oxidize aromatic compounds, and can be divided into two functional classes [5]. The first group consists of laccases and laccase-like enzymes (e.g., laccase-like polyphenol oxidases in Arabidopsis thaliana encoded by TT10 gene) [6], favoring oxidation of organic substrates over metal ions. The second group oxidizes metal ions (Fe (II), Cu (I) and/or Mn (II)) with high efficiency, and these MCOs are generally referred to as metal oxidases [7]. These enzymes catalyze oxidation of substrates with a concomitant reduction of molecular oxygen to water. Detailed spectroscopic and X-ray crystallographic analyses of electronic and geometric structure of the active site of MCOs reveal that the catalytic center responsible for these reactions is comprised of four or more copper atoms, leading to three T types which are T1 (blue copper), T2 (normal copper), and T3 (binuclear copper center). Due to the role of Cu in a diverse array of biological oxidation–reduction reactions as a cofactor, MCOs also have a high redox potential in a wide variety of reactions [8,9]. The substrate undergoes oxidation in the T1 center having the His-Cys-His (characteristic of MCOs) sequence, where the Cu atom is bound by two histidine residues and one cysteine forming a distorted trigonal pyramid structure. The unique structure of the MCO enzymes allows the binding and reduction of molecular oxygen into water [10]. Most MCOs contain around 500 amino acid residues and display a β-sheet structure, forming a Greek Key motif [11]. The three main steps of MCO catalytic mechanisms include: (1) T1 Cu site reduction through the capture of an electron from the oxidized substrate, (2) electron transfer from the T1 site to the trinuclear Cu cluster, and (3) O2 reduction to two H2O molecules. Structurally, laccases belong to the group of multicopper blue proteins that contain two domains, whereas yeast ferroxidases are six domain-containing multicopper blue proteins, similarly to human ceruloplasmin. An X-ray crystallography study showed that laccases do not undergo drastic conformational changes after losing their Cu atom, contrary to ceruloplasmin [12].
The influence of several laccases and ferroxidases on pathogenesis has been reported in various plant pathogenic fungi such as Magnaporthe grisea, Ophiostoma novo-ulmi, Gaeumannomyces graminis, and Ustilago maydis [13,14,15,16]. Ferroxidases present in the plant pathogen Colletotrichum graminicola complement the growth defect of ferroxidase mutants in Saccharomyces cerevisae [17]. In the white rot fungus, Phanerochaete chrysosporium, a new class of MCOs has been discovered which is structurally different from conventional fungal laccases [18]. A recent study also revealed gene duplication events, the evolutionary history of MCO genes and the importance of horizontal gene transfer in coprophilous and non-coprophilous fungi [19]. The presence of various MCOs and their importance is also reported in diverse human fungal pathogens. Specifically, ferroxidases involved in iron oxidation and laccases involved in pigmentation and their association with virulence have been reported by different research groups. The focus of the current review is to provide an updated knowledge of ferroxidases in S. cerevisiae and ferroxidases as well as laccase in different human pathogenic fungi and their role in iron metabolism and pathogenesis development.
2. Multicopper Oxidases in Saccharomyces cerevisiae
Although not typically pathogenic in humans, studies in S. cerevisiae provide significant insights into MCOs in virulent fungi. Iron uptake is a two-step process in this species: (1) first, reduction of Fe (III) by iron reductases in the plasma membrane, followed by (2) the internalization of the Fe (II) products by either of two Fe (II) uptake systems. The plasma membrane-based Fe (III) reduction is a high-affinity system with a Km = 0.15 µM that requires FET3 and is induced by low iron concentration. Fet3p is an essential component of the high-affinity reductive iron assimilation (RIA) pathway in fungi, which functions in combination with Ftr1p [20]. Fet3p functions similarly to human ceruloplasmin [21,22,23]. The internalization step is characterized by low affinity (Km = 30 µM) and requires Fet4p, which is generally active under iron-replete conditions [23]. Surprisingly, the FET3-FET4 double deletion mutant of this species remains viable; however, it is extremely sensitive to iron limitation. This result led to the discovery of another ferroxidase in S. cerevisiae, namely FET5. Overexpression of this gene suppressed the growth defect of the fet3Δ/Δ- fet4Δ/Δ mutant under iron-limited conditions, confirming that Fet5p also plays a role in iron transport and homeostasis [24]. Fet3p forms a complex with the iron permease Ftr1p, a transmembrane protein. In S. cerevisiae (likely most fungi and plants), the extracellular Fe (III) is mobilized after the reduction to Fe (II) by the surface metalloreductase Fre1p and then oxidized by Fet3p. The oxidized form of Fe (III) is not a substrate of Ftr1p, so the iron uptake is paired strictly with ferroxidation. This permease-oxidase based transport system in S. cerevisiae provides a model that explains the copper-iron connection in biology [25,26]. Structural analysis shows that Fet3p belongs to the family of integral membrane protein (type 1). The N-terminal catalytic domain of the protein is located on the extracellular side of the plasma membrane and the single C-terminus transmembrane domain is tethered to the membrane [27]. The detailed 3D structure of the Fet3p extracellular ferroxidase domain shows a unique characteristic that enables Fe transport in eukaryotes. This structure also reveals structural differences between Fet3p and laccases [28,29]. Figure 1 schematically summarizes the presence of different MCOs in S. cerevisiae and various human pathogenic fungi.
3. Multicopper Oxidases and Their Function in Human Pathogenic Budding Yeast
3.1. Candida albicans
C. albicans is the most frequently isolated species of invasive Candida infections [30]. It is an opportunistic pathogenic fungus that is also a member of the healthy human mycobiota. It is able to cause both superficial infections of the skin or mucosal surfaces and invasive infections, where the fungus can spread to all vital organs through dissemination [31,32]. In order to survive within different host niches with restricted amounts of accessible iron, C. albicans has evolved various iron uptake mechanisms. Iron is an essential micronutrient for both the host and C. albicans, therefore iron uptake during infection also plays a role in the pathogenesis of this fungus [33]. Similar to S. cerevisiae, C. albicans has reductive iron uptake machinery and has several genes encoding ferric reductases (17 putative genes) and five ferroxidases belong to MCO in its genome [34].
The expression of the five ferroxidases, namely FET3 (orf19.4211), FET31 (orf19.4213), FET33 (orf19.943), FET34 (orf19.4215), and FET99 (orf19.4212), varies depending on environmental conditions, including the availability of iron and oxygen and the presence of antifungals [35,36,37]. Functional characterization of these ferroxidases confirmed their role in iron uptake, morphological transition, and pathogenesis. A homozygous deletion mutant of FET31 (referred to as FET3 in the publication) showed a growth defect under low iron conditions; however, the mutant strain was as virulent as the wild type strain in a mouse model of systemic candidiasis [38]. CaFET3 and CaFET34 are important ferroxidases under iron starvation conditions. Specifically, deletion of CaFET33 and CaFET34 decrease the cellular iron content under iron limited conditions, although the mutants do not exhibit a growth defect. Deletion of CaFET34 also significantly reduces filamentous growth and the virulence of C. albicans in a mouse model of systemic candidiasis [39]. Fet31p and Fet34p are localized to the plasma membrane and participate in Fe-uptake by forming a complex with Ftr1p [40]. Interestingly, CaFET3 has also been associated with fungal prostaglandin E2 production [41].
3.2. Candida parapsilosis
C. parapsilosis is also an opportunistic human fungal pathogen, and depending on the country, it is the second or third most frequently isolated Candida species in immunocompromised patients with candidemia [42]. C. parapsilosis accounts for the highest numbers of candidiasis episodes in premature or low birth weight infants and can exist in diverse environments besides the human mycobiome. It typically exists in a yeast form, though it can generate pseudohyphae and it avidly forms multidrug resistant biofilms on abiotic surfaces such as dentures or catheters [42,43]. Although various virulence factors are associated with this fungus’ pathogenicity, the molecular mechanisms of iron metabolism and homoeostasis are poorly understood. Recently, however, three putative ferroxidase encoding genes were identified in its genome, which show high sequence similarity with ScFET3. A study of gene-deletion strains of CPAR2_603600 (79% identity with CaFET3 and 54% identity with ScFET3) demonstrated that this gene is required for fungal growth under low iron conditions. The deletion mutant revealed a marked reduction in pseudohyphae and biofilm formation as well as alterations in the expression of many orthologous genes potentially involved in iron metabolism regulation (in press). Unlike in C. albicans, the deletion mutant had attenuated virulence in a mouse model of systemic candidiasis, suggesting a divergent role in these and possibly other species. Similar to CaFET3, this gene in C. parapsilosis also plays a role in PGE2 production from externally added arachidonic acids [44,45].
3.3. Candida glabrata
C. glabrata is also an opportunistic human fungal pathogen and part of the healthy human mycobiota [46]. C. glabrata is a unicellular budding yeast and phylogenetically closer to S. cerevisiae than to C. albicans. Approximately 12% of total bloodstream Candida infections globally are caused by C. glabrata and the mortality rate can reach up to 30% [46,47,48,49]. In silico analysis has revealed three ferroxidase genes, namely CgFET3 (high affinity iron uptake; 70% similarity with ScFET3), CgFET4 (low-affinity ferrous transporter of the plasma membrane; 58% similarity with ScFET4), and CgFET5 (iron storage and utilization; 64% similarity with ScFET5). Phenotypic profiling of the fet3Δ/Δ deletion mutant in C. glabrata revealed that FET3 is required for growth in the presence of the iron chelator bathophenanthrolinedisulfonic acid (BPS), suggesting that FET3 is a part of the high-affinity iron uptake system in C. glabrata. The mutant strain is sensitive to oxidative stress and to the antifungal drug fluconazole, and also displays attenuated growth on media containing sodium chloride (osmotic stressor), caffeine and congo red (cell wall stressors), and SDS (sodium dodecyl sulfate, membrane stressor). Comparison of the intracellular iron levels reveals that the iron content of the CgFET3 deletion mutant is ∼20%–50% lower than wild type cell. Disruption of this gene also resulted in ∼10%–20% reduction in the mitochondrial aconitase activity, which is a Fe-S containing protein [50]. C. glabrata is able to grow inside macrophages [51], but the deletion of CgFET3 and CgFET5 reduces this ability. Fungal burden analysis of the kidneys of mice revealed lower CFUs in mice infected with the mutant strains compared to the wild type [50]. Hence, FET3 and FET5 regulate the pathogenesis of C. glabrata. The homologue of an mRNA-degrading protein Cth2 has also been shown to regulate the expression FET3 in C. glabrata [52,53].
3.4. Cryptococcus neoformans
Cryptococcosis remains a leading cause of death in the HIV/AIDS population as it is estimated that there are ~180,000 deaths annually, mostly in sub-Saharan Africa [54,55]. Most of these patients are either infected with C. neoformans or, to a lesser extent, C. gattii [54]. The two main components of the C. neoformans high affinity iron uptake system are the iron permease CFT1 and the ferroxidase CFO1. Both CFT1 and CFO1 are present on chromosome 12 and are divergently transcribed. C. neoformans also contains CFT2 and CFO2, which encode an iron permease and a ferroxidase, respectively, and are present on chromosome 3 [56]. Deletion mutants of each revealed that CFO1 is required for high-affinity iron transport, however, the lack of CFO2 led to no visible growth defect under limited iron conditions [57]. Using a mouse inhalation model of cryptococcosis, the virulence of the cfo1Δ/Δ, cfo2Δ/Δ and the cfo1Δ- cfo2 Δ double mutants was analyzed. The cfo1Δ single mutant as well as the cfo1Δ- cfo2Δ double mutant showed a significantly attenuated virulence compared to the wild type strain [58]. Similarly to the equivalent C. glabrata mutants, these mutant strains in C. neoformans were also more susceptible to fluconazole [58]. A localization study using GFP tagged CFO1 revealed that the protein is localized to plasma membrane [59]. Interestingly, it has been suggested that the cAMP pathway regulates the intracellular trafficking of Cfo1p, as a defect in cAMP signaling leads to mis-localization of the Cfo1-GFP fusion protein [59].
C. neoformans also contains homologues of a laccase encoded by LAC1 and LAC2 expressed from the same chromosome. Deletion mutants of both the genes show significant reduction in melanin production [60]. The formation of melanin in the cell wall of this species protects the cell from different environmental stress conditions and host immune attacks [61]. The deletion mutant of these two genes also make the cells more susceptible to killing by alveolar macrophages [62]. The LAC1 gene was also shown to be regulating fungal prostaglandin E2 production [63]. The LAC1 gene also plays an important role in C. neoformans’ virulence as the mutant with the disrupted gene was not lethal in the mouse infection model [64].
4. Multicopper Oxidases and Their Function in Other Human Pathogenic Fungi
4.1. Mucor circinelloides
Mucormycosis is an emerging fungal infection threatening mainly immunocompromised patients suffering from diabetes or cancer or those underwent organ transplantation. The mortality rates of mucormycosis can reach as high as 90% in disseminated infections, which is a consequence of the lack of effective treatments and antifungal drug resistance [65,66,67,68]. M. circinelloides, a frequently used model to study mucormycosis, is a dimorphic fungus that can multiply through the formation of branching coenocytic hyphae under aerobic conditions or spherical multipolar budding yeasts in an oxygen deprived environment [69]. Three putative ferroxidase genes have been identified that show sequence similarity with ScFET3: fet3a, fet3b, and fet3c. Under aerobic conditions, only fet3a is expressed during the yeast phase, while the other two are specifically expressed in mycelia. Gene expression and deletion analysis of these genes revealed their necessity for growth under iron-limited conditions (media with iron chelator phenanthroline). Murine experiments with the fet3aΔ/Δ, fet3bΔ/Δ and fet3cΔ/Δ strains revealed that only fet3c significantly impacts virulence [70].
4.2. Histoplasma capsulatum
Histoplasma capsulatum is an intracellular pathogen that is the main cause of histoplasmosis in both immunocompetent and immunocompromised individuals. Histoplasma is a dimorphic fungus that is mycelial in nature and a yeast during mammalian disease [71]. Ohio and Mississippi river valleys, and the southeastern, central, and mid-Atlantic states report the most cases of histoplasmosis, and the infection rate can reach up to almost 500,000 individuals annually [72]. Although previous in silico analyses of the H. capsulatum G186AR genome identified orthologues of S. cerevisiae FET3 and FTR1 (sequence similarity: 65% for FET3 and 61% for FTR1, respectively), functional studies have not yet been performed. Interestingly, in silico analyses showed that H. capsulatum strain (G217B) lacks the orthologues of either of these genes in the genome [73,74].
4.3. Aspergillus fumigatus
Invasive aspergillosis is the most common cause of airborne fungal invasive infections among immunocompromised patients worldwide and is primarily caused by A. fumigatus [75]. Patients with prolonged neutropenia are particularly susceptible [76]. The mortality rate of invasive aspergillosis is ~50% and increases in the setting of drug resistant strains [77,78,79,80]. Four iron uptake systems have been described in A. fumigatus: the low affinity ferrous iron uptake system (not yet characterized at the molecular level), two siderophore mediated high affinity ferric iron uptake systems, and the reductive iron assimilation system (RIA) [81]. Disruption of sreA, mediating siderophore biosynthesis in A. fumigatus, results in decreased virulence in mice [82,83]. The three major components of reductive iron assimilation are the ferric reductase FreB which helps in reduction of ferric to ferrous iron, then the import of ferrous iron through iron permease FtrA, and finally, oxidation by ferrous to ferric iron by ferroxidase FetC. FetCp is 52% identical to C. albicans Fet3p and FtrAp is 55% identical to C. albicans Ftr1p at the amino acid level [81,84] Although FetC is upregulated under iron-restricted conditions in A. fumigatus, a detailed characterization of this MCO has not been performed yet.
A. fumigatus contains two putative laccase encoding genes, namely Abr1 and Abr2, that are part of a gene cluster participating in melanin synthesis. The expression of A. fumigatus laccases Abr1/2 is dependent on hyphal competency and significantly increased during conidiation [85]. The abr2Δ/Δ mutant strain showed increased sensitivity to reactive oxygen species and reduced laccase activity in sporulating mycelia. However, unlike in C. neoformans, Abr2 deletion in A. fumigatus did not show any reduction in virulence in an intranasal mouse infection model [86]. Previous sequence analyses showed that the Abr1 protein does not contain the four residues that are present in S. cerevisiae Fet3 for Fe(II) binding, only the glutamic acid (E185) [87]. This also suggests that they represent a separate class of MCOs, evolved from an ancient canonical ferroxidase [88]. However, the deletion mutant of the arp1 gene (regulating melanin production in Colletotrichum lagenarium and Magnaporthe grisea) in A. fumigatus produced reddish-pink conidia that are more susceptible to complement attack [89].
5. Conclusions
The family of MCOs is one of the most diverse family of enzymes, having a wide variety of functions. The importance of MCOs and their role in metal homoeostasis is well known in various human pathogens including bacteria, fungi, and parasites, although their detailed roles in virulence is still not fully explored. Multicopper oxidases are ubiquitous in the fungal kingdom which shows their evolutionary importance. In the current review, we aimed to collect all relevant information about different MCOs present in human fungal pathogens belonging to diverse fungal kingdoms, such as Ascomycetes, Basidiomycetes, and Zygomycetes. They have evolved different pathogenic mechanisms to infect human hosts. Our knowledge of the role of iron in fungal pathogenicity has advanced over recent years, however still little information is available about the precise role and inclusion of reductive iron uptake systems—including the Fet/Ftr complex—in pathogenicity mechanisms. For instance, the presence of iron for the human pathogenic Paracoccidioides species also influences virulence, although no detailed study is available about the MCOs of this particular fungus. Only in silico analyses suggest that PAAG_06004 and PADG_05994 could encode functional ferroxidases [90]. Laccases also play an important role in fungal pathogens by generating mainly melanin. However, more research is needed to address why some of the pathogenic fungi evolved a laccase enzyme system with the reductive iron system. Moreover, the presence of this evolutionary conserved system in pathogenic fungal species can be a potential target for selective therapeutic intervention in multiple mycoses.
Author Contributions
T.C. and A.G. created the concept of the review. T.C. prepared the initial manuscript with the figure, which were edited, corrected and carefully reviewed by R.T. and J.D.N. A.G. finalized and approved the concluding study. All authors have read and agreed to the published version of the manuscript.
Funding
T.C. was supported by the European Union Seventh Framework Programme (FP7/2007-2013) under grant agreements FP7-PEOPLE-2013-ITN-606786 “ImResFun”. A.G. were supported by grants 20,391 3/2018/FEKUSTRAT, NKFIH K 123952, and GINOP-2.3.2.-15-2016-00015. A.G. was additionally funded by LP2018-15/2018. J.D.N. was supported by US NIH RO1 AI52733.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rydén, L.G.; Hunt, L.T. Evolution of protein complexity: The blue copper-containing oxidases and related proteins. J. Mol. Evol. 1993, 36, 41–66. [Google Scholar] [CrossRef] [PubMed]
- Hoegger, P.J.; Kilaru, S.; James, T.Y.; Thacker, J.R.; Kues, U. Phylogenetic comparison and classification of laccase and related multicopper oxidase protein sequences. FEBS J. 2006, 273, 2308–2326. [Google Scholar] [CrossRef] [PubMed]
- Reiss, R.; Ihssen, J.; Richter, M.; Eichhorn, E.; Schilling, B.; Thöny-Meyer, L. Laccase versus Laccase-Like Multi-Copper Oxidase: A Comparative Study of Similar Enzymes with Diverse Substrate Spectra. PLoS ONE 2013, 8, e65633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirim, D.; Wagner, F.; Wang, L.; Schmid, R.D.; Pleiss, J. The Laccase Engineering Database: A classification and analysis system for laccases and related multicopper oxidases. Database 2011, 2011, bar006. [Google Scholar] [CrossRef] [Green Version]
- Martins, L.O.; Durao, P.; Brissos, V.; Lindley, P.F. Laccases of prokaryotic origin: Enzymes at the interface of protein science and protein technology. Cell. Mol. Life Sci. 2015, 72, 911–922. [Google Scholar] [CrossRef]
- Pourcel, L.; Routaboul, J.-M.; Kerhoas, L.; Caboche, M.; Lepiniec, L.; Debeaujon, I. TRANSPARENT TESTA10 encodes a laccase-like enzyme involved in oxidative polymerization of flavonoids in Arabidopsis seed coat. Plant Cell 2005, 17, 2966–2980. [Google Scholar] [CrossRef] [Green Version]
- Stoj, C.; Kosman, D.J. Cuprous oxidase activity of yeast Fet3p and human ceruloplasmin: Implication for function. FEBS Lett. 2003, 554, 422–426. [Google Scholar] [CrossRef] [Green Version]
- Solomon, E.I.; Sundaram, U.M.; Machonkin, T.E. Multicopper Oxidases and Oxygenases. Chem. Rev. 1996, 96, 2563–2606. [Google Scholar] [CrossRef]
- Góralczyk-bińkowska, A.; Jasińska, A.; Długoński, J. Characteristics and use of multicopper oxidases enzymes. Post. Mikrobiol. 2019, 58, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, T.; Kataoka, K. Basic and applied features of multicopper oxidases, CueO, bilirubin oxidase, and laccase. Chem. Rec. 2007, 7, 220–229. [Google Scholar] [CrossRef] [Green Version]
- Kosman, D.J. Multicopper oxidases: A workshop on copper coordination chemistry, electron transfer, and metallophysiology. J. Biol. Inorg. Chem. 2010, 15, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; GO, N. Function and molecular evolution of multicopper blue proteins. Cell. Mol. Life Sci. 2005, 62, 2050–2066. [Google Scholar] [CrossRef] [PubMed]
- Iyer, G.; Chattoo, B.B. Purification and characterization of laccase from the rice blast fungus, Magnaporthe grisea. FEMS Microbiol. Lett. 2003, 227, 121–126. [Google Scholar] [CrossRef] [Green Version]
- Binz, T.; Canevascini, G. Purification and Partial Characterization of the Extracellular Laccase from Ophiostoma novo-ulmi. Curr. Microbiol. 1997, 35, 278–281. [Google Scholar] [CrossRef]
- Edens, W.A.; Goins, T.Q.; Dooley, D.; Henson, J.M. Purification and characterization of a secreted laccase of Gaeumannomyces graminis var. tritici. Appl. Environ. Microbiol. 1999, 65, 3071–3074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichhorn, H.; Lessing, F.; Winterberg, B.; Schirawski, J.; Kamper, J.; Muller, P.; Kahmann, R. A ferroxidation/permeation iron uptake system is required for virulence in Ustilago maydis. Plant Cell 2006, 18, 3332–3345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albarouki, E.; Deising, H.B. Infection structure-specific reductive iron assimilation is required for cell wall integrity and full virulence of the maize pathogen Colletotrichum graminicola. Mol. Plant Microbe Interact. 2013, 26, 695–708. [Google Scholar] [CrossRef] [Green Version]
- Larrondo, L.F.; Salas, L.; Melo, F.; Vicuña, R.; Cullen, D. A Novel Extracellular Multicopper Oxidase from Phanerochaete chrysosporium with Ferroxidase Activity. Appl. Environ. Microbiol. 2003, 69, 6257–6263. [Google Scholar] [CrossRef] [Green Version]
- Poggeler, S. Evolution of multicopper oxidase genes in coprophilous and non-coprophilous members of the order sordariales. Curr. Genom. 2011, 12, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Kosman, D.J. Molecular mechanisms of iron uptake in fungi. Mol. Microbiol. 2003, 47, 1185–1197. [Google Scholar] [CrossRef]
- De Silva, D.M.; Askwith, C.C.; Eide, D.; Kaplan, J. The FET3 gene product required for high affinity iron transport in yeast is a cell surface ferroxidase. J. Biol. Chem. 1995, 270, 1098–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askwith, C.; Eide, D.; Van Ho, A.; Bernard, P.S.; Li, L.; Davis-Kaplan, S.; Sipe, D.M.; Kaplan, J. The FET3 gene of S. cerevisiae encodes a multicopper oxidase required for ferrous iron uptake. Cell 1994, 76, 403–410. [Google Scholar] [CrossRef]
- Dix, D.R.; Bridgham, J.T.; Broderius, M.A.; Byersdorfer, C.A.; Eide, D.J. The FET4 gene encodes the low affinity Fe(II) transport protein of Saccharomyces cerevisiae. J. Biol. Chem. 1994, 269, 26092–26099. [Google Scholar] [PubMed]
- Spizzo, T.; Byersdorfer, C.; Duesterhoeft, S.; Eide, D. The yeast FET5 gene encodes a FET3-related multicopper oxidase implicated in iron transport. Mol. Gen. Genet. 1997, 256, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Stearman, R.; Yuan, D.S.; Yamaguchi-Iwai, Y.; Klausner, R.D.; Dancis, A. A permease-oxidase complex involved in high-affinity iron uptake in yeast. Science 1996, 271, 1552–1557. [Google Scholar] [CrossRef] [PubMed]
- De Freitas, J.; Wintz, H.; Kim, J.H.; Poynton, H.; Fox, T.; Vulpe, C. Yeast, a model organism for iron and copper metabolism studies. Biometals 2003, 16, 185–197. [Google Scholar] [CrossRef]
- Yuan, D.S.; Stearman, R.; Dancis, A.; Dunn, T.; Beeler, T.; Klausner, R.D. The Menkes/Wilson disease gene homologue in yeast provides copper to a ceruloplasmin-like oxidase required for iron uptake. Proc. Natl. Acad. Sci. USA 1995, 92, 2632–2636. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.B.; Stoj, C.S.; Ziegler, L.; Kosman, D.J.; Hart, P.J. The copper-iron connection in biology: Structure of the metallo-oxidase Fet3p. Proc. Natl. Acad. Sci. USA 2005, 102, 15459–15464. [Google Scholar] [CrossRef] [Green Version]
- Stoj, C.S.; Augustine, A.J.; Zeigler, L.; Solomon, E.I.; Kosman, D.J. Structural basis of the ferrous iron specificity of the yeast ferroxidase, Fet3p. Biochemistry 2006, 45, 12741–12749. [Google Scholar] [CrossRef]
- Brown, G.D.; Denning, D.W.; Gow, N.A.R.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef] [Green Version]
- Brandt, M.E. Candida and Candidiasis. Emerg. Infect. Dis. 2002, 8, 876. [Google Scholar] [CrossRef]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutak, R.; Lesuisse, E.; Tachezy, J.; Richardson, D.R. Crusade for iron: Iron uptake in unicellular eukaryotes and its significance for virulence. Trends Microbiol. 2008, 16, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.S.; Wilson, D.; Hube, B. Candida albicans iron acquisition within the host. FEMS Yeast Res. 2009, 9, 1000–1012. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.T.; Lee, R.E.B.; Barker, K.S.; Lee, R.E.; Wei, L.; Homayouni, R.; Rogers, P.D. Genome-wide expression profiling of the response to azole, polyene, echinocandin, and pyrimidine antifungal agents in Candida albicans. Antimicrob. Agents Chemother. 2005, 49, 2226–2236. [Google Scholar] [CrossRef] [Green Version]
- Rogers, P.D.; Barker, K.S. Evaluation of differential gene expression in fluconazole-susceptible and -resistant isolates of Candida albicans by cDNA microarray analysis. Antimicrob. Agents Chemother. 2002, 46, 3412–3417. [Google Scholar] [CrossRef] [Green Version]
- Synnott, J.M.; Guida, A.; Mulhern-Haughey, S.; Higgins, D.G.; Butler, G. Regulation of the hypoxic response in Candida albicans. Eukaryot. Cell 2010, 9, 1734–1746. [Google Scholar] [CrossRef] [Green Version]
- Eck, R.; Hundt, S.; Hartl, A.; Roemer, E.; Kunkel, W. A multicopper oxidase gene from Candida albicans: Cloning, characterization and disruption. Microbiology 1999, 145 Pt 9, 2415–2422. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Xu, N.; Yu, Q.; Ding, X.; Qian, K.; Zhao, Q.; Wang, Y.; Zhang, B.; Xing, L.; Li, M. Novel insight into the expression and function of the multicopper oxidases in Candida albicans. Microbiology 2013, 159, 1044–1055. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, L.; Terzulli, A.; Gaur, R.; McCarthy, R.; Kosman, D.J. Functional characterization of the ferroxidase, permease high-affinity iron transport complex from Candida albicans. Mol. Microbiol. 2011, 81, 473–485. [Google Scholar] [CrossRef] [Green Version]
- Erb-Downward, J.R.; Noverr, M.C. Characterization of prostaglandin E2 production by Candida albicans. Infect. Immun. 2007, 75, 3498–3505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toth, R.; Nosek, J.; Mora-Montes, H.M.; Gabaldon, T.; Bliss, J.M.; Nosanchuk, J.D.; Turner, S.A.; Butler, G.; Vagvolgyi, C.; Gacser, A. Candida parapsilosis: From Genes to the Bedside. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trofa, D.; Gacser, A.; Nosanchuk, J.D. Candida parapsilosis, an emerging fungal pathogen. Clin. Microbiol. Rev. 2008, 21, 606–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, T.; Thuer, E.; Heijink, M.; Toth, R.; Bodai, L.; Vagvolgyi, C.; Giera, M.; Gabaldon, T.; Gacser, A. Eicosanoid biosynthesis influences the virulence of Candida parapsilosis. Virulence 2018, 9, 1019–1035. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, T.; Toth, R.; Gacser, A. Eicosanoid production by Candida parapsilosis and other pathogenic yeasts. Virulence 2019, 10, 970–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fidel, P.L.J.; Vazquez, J.A.; Sobel, J.D. Candida glabrata: Review of epidemiology, pathogenesis, and clinical disease with comparison to C. albicans. Clin. Microbiol. Rev. 1999, 12, 80–96. [Google Scholar] [CrossRef] [Green Version]
- Kaur, R.; Domergue, R.; Zupancic, M.L.; Cormack, B.P. A yeast by any other name: Candida glabrata and its interaction with the host. Curr. Opin. Microbiol. 2005, 8, 378–384. [Google Scholar] [CrossRef]
- Klevay, M.J.; Horn, D.L.; Neofytos, D.; Pfaller, M.A.; Diekema, D.J. Initial treatment and outcome of Candida glabrata versus Candida albicans bloodstream infection. Diagn. Microbiol. Infect. Dis. 2009, 64, 152–157. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Diekema, D.J.; Gibbs, D.L.; Newell, V.A.; Ellis, D.; Tullio, V.; Rodloff, A.; Fu, W.; Ling, T.A. Results from the ARTEMIS DISK Global Antifungal Surveillance Study, 1997 to 2007: A 10.5-year analysis of susceptibilities of Candida Species to fluconazole and voriconazole as determined by CLSI standardized disk diffusion. J. Clin. Microbiol. 2010, 48, 1366–1377. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, V.K.; Suneetha, K.J.; Kaur, R. A systematic analysis reveals an essential role for high-affinity iron uptake system, haemolysin and CFEM domain-containing protein in iron homoeostasis and virulence in Candida glabrata. Biochem. J. 2014, 463, 103–114. [Google Scholar] [CrossRef]
- Kaur, R.; Ma, B.; Cormack, B.P. A family of glycosylphosphatidylinositol-linked aspartyl proteases is required for virulence of Candida glabrata. Proc. Natl. Acad. Sci. USA 2007, 104, 7628–7633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerwien, F.; Safyan, A.; Wisgott, S.; Hille, F.; Kaemmer, P.; Linde, J.J.; Brunke, S.; Kasper, L.; Hube, B. A novel hybrid iron regulation network combines features from pathogenic and nonpathogenic yeasts. MBio 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devaux, F.; Thiebaut, A. The regulation of iron homeostasis in the fungal human pathogen Candida glabrata. Microbiology 2019, 165, 1041–1060. [Google Scholar] [CrossRef]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Park, B.J.; Wannemuehler, K.A.; Marston, B.J.; Govender, N.; Pappas, P.G.; Chiller, T.M. Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS. AIDS 2009, 23, 525–530. [Google Scholar] [CrossRef]
- Kronstad, J.W.; Hu, G.; Jung, W.H. An encapsulation of iron homeostasis and virulence in Cryptococcus neoformans. Trends Microbiol. 2013, 21, 457–465. [Google Scholar] [CrossRef] [Green Version]
- Hu, G.; Cheng, P.-Y.; Sham, A.; Perfect, J.R.; Kronstad, J.W. Metabolic adaptation in Cryptococcus neoformans during early murine pulmonary infection. Mol. Microbiol. 2008, 69, 1456–1475. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Cho, Y.-J.; Do, E.; Choi, J.; Hu, G.; Cadieux, B.; Chun, J.; Lee, Y.; Kronstad, J.W.; Jung, W.H. A defect in iron uptake enhances the susceptibility of Cryptococcus neoformans to azole antifungal drugs. Fungal Genet. Biol. 2012, 49, 955–966. [Google Scholar] [CrossRef] [Green Version]
- Jung, W.H.; Hu, G.; Kuo, W.; Kronstad, J.W. Role of ferroxidases in iron uptake and virulence of Cryptococcus neoformans. Eukaryot. Cell 2009, 8, 1511–1520. [Google Scholar] [CrossRef] [Green Version]
- Missall, T.A.; Moran, J.M.; Corbett, J.A.; Lodge, J.K. Distinct Stress Responses of Two Functional Laccases in Cryptococcus neoformans Are Revealed in the Absence of the Thiol-Specific Antioxidant Tsa1. Eukaryot. Cell 2005, 4, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Eisenman, H.C.; Mues, M.; Weber, S.E.; Frases, S.; Chaskes, S.; Gerfen, G.; Casadevall, A. Cryptococcus neoformans laccase catalyses melanin synthesis from both D- and L-DOPA. Microbiology 2007, 153, 3954–3962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Tewari, R.P.; Williamson, P.R. Laccase Protects Cryptococcus neoformans from Antifungal Activity of Alveolar Macrophages. Infect. Immun. 1999, 67, 6034–6039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erb-Downward, J.R.; Noggle, R.M.; Williamson, P.R.; Huffnagle, G.B. The role of laccase in prostaglandin production by Cryptococcus neoformans. Mol. Microbiol. 2008, 68, 1428–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas, S.D.; Bennett, J.E.; Kwon-Chung, K.J.; Perfect, J.R.; Williamson, P.R. Effect of the laccase gene CNLAC1, on virulence of Cryptococcus neoformans. J. Exp. Med. 1996, 184, 377–386. [Google Scholar] [CrossRef]
- Petrikkos, G.; Skiada, A.; Lortholary, O.; Roilides, E.; Walsh, T.J.; Kontoyiannis, D.P. Epidemiology and clinical manifestations of mucormycosis. Clin. Infect. Dis. 2012, 54 (Suppl. 1), S23–S34. [Google Scholar] [CrossRef]
- Sridhara, S.R.; Paragache, G.; Panda, N.K.; Chakrabarti, A. Mucormycosis in immunocompetent individuals: An increasing trend. J. Otolaryngol. 2005, 34, 402–406. [Google Scholar] [CrossRef]
- Roden, M.M.; Zaoutis, T.E.; Buchanan, W.L.; Knudsen, T.A.; Sarkisova, T.A.; Schaufele, R.L.; Sein, M.; Sein, T.; Chiou, C.C.; Chu, J.H.; et al. Epidemiology and outcome of zygomycosis: A review of 929 reported cases. Clin. Infect. Dis. 2005, 41, 634–653. [Google Scholar] [CrossRef] [Green Version]
- Dannaoui, E. Antifungal resistance in mucorales. Int. J. Antimicrob. Agents 2017, 50, 617–621. [Google Scholar] [CrossRef]
- Orlowski, M. Mucor dimorphism. Microbiol. Rev. 1991, 55, 234–258. [Google Scholar] [CrossRef]
- Navarro-Mendoza, M.I.; Pérez-Arques, C.; Murcia, L.; Martínez-García, P.; Lax, C.; Sanchis, M.; Capilla, J.; Nicolás, F.E.; Garre, V. Components of a new gene family of ferroxidases involved in virulence are functionally specialized in fungal dimorphism. Sci. Rep. 2018, 8, 7660. [Google Scholar] [CrossRef] [Green Version]
- Maresca, B.; Kobayashi, G.S. Dimorphism in Histoplasma capsulatum: A model for the study of cell differentiation in pathogenic fungi. Microbiol. Rev. 1989, 53, 186–209. [Google Scholar] [CrossRef] [PubMed]
- Maiga, A.W.; Deppen, S.; Scaffidi, B.K.; Baddley, J.; Aldrich, M.C.; Dittus, R.S.; Grogan, E.L. Mapping Histoplasma capsulatum Exposure, United States. Emerg. Infect. Dis. 2018, 24, 1835–1839. [Google Scholar] [CrossRef] [PubMed]
- Newman, S.L.; Smulian, A.G. Iron uptake and virulence in Histoplasma capsulatum. Curr. Opin. Microbiol. 2013, 16, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Hilty, J.; George Smulian, A.; Newman, S.L. Histoplasma capsulatum utilizes siderophores for intracellular iron acquisition in macrophages. Med. Mycol. 2011, 49, 633–642. [Google Scholar]
- Kontoyiannis, D.P.; Chamilos, G.; Lewis, R.E.; Giralt, S.; Cortes, J.; Raad, I.I.; Manning, J.T.; Han, X. Increased bone marrow iron stores is an independent risk factor for invasive aspergillosis in patients with high-risk hematologic malignancies and recipients of allogeneic hematopoietic stem cell transplantation. Cancer 2007, 110, 1303–1306. [Google Scholar] [CrossRef]
- Abers, M.S.; Ghebremichael, M.S.; Timmons, A.K.; Warren, H.S.; Poznansky, M.C.; Vyas, J.M. A Critical Reappraisal of Prolonged Neutropenia as a Risk Factor for Invasive Pulmonary Aspergillosis. Open Forum Infect. Dis. 2016, 3, ofw036. [Google Scholar] [CrossRef] [Green Version]
- Dagenais, T.R.T.; Keller, N.P. Pathogenesis of Aspergillus fumigatus in Invasive Aspergillosis. Clin. Microbiol. Rev. 2009, 22, 447–465. [Google Scholar] [CrossRef] [Green Version]
- Lass-Florl, C.; Roilides, E.; Loffler, J.; Wilflingseder, D.; Romani, L. Minireview: Host defence in invasive aspergillosis. Mycoses 2013, 56, 403–413. [Google Scholar] [CrossRef]
- Lewis, R.E.; Lortholary, O.; Spellberg, B.; Roilides, E.; Kontoyiannis, D.P.; Walsh, T.J. How does antifungal pharmacology differ for mucormycosis versus aspergillosis? Clin. Infect. Dis. 2012, 54 (Suppl. 1), S67–S72. [Google Scholar] [CrossRef] [Green Version]
- van der Linden, J.W.M.; Snelders, E.; Kampinga, G.A.; Rijnders, B.J.A.; Mattsson, E.; Debets-Ossenkopp, Y.J.; Kuijper, E.J.; Van Tiel, F.H.; Melchers, W.J.G.; Verweij, P.E. Clinical implications of azole resistance in Aspergillus fumigatus, The Netherlands, 2007–2009. Emerg. Infect. Dis. 2011, 17, 1846–1854. [Google Scholar] [CrossRef]
- Schrettl, M.; Bignell, E.; Kragl, C.; Joechl, C.; Rogers, T.; Arst, H.N.J.; Haynes, K.; Haas, H. Siderophore biosynthesis but not reductive iron assimilation is essential for Aspergillus fumigatus virulence. J. Exp. Med. 2004, 200, 1213–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrettl, M.; Kim, H.S.; Eisendle, M.; Kragl, C.; Nierman, W.C.; Heinekamp, T.; Werner, E.R.; Jacobsen, I.; Illmer, P.; Yi, H.; et al. SreA-mediated iron regulation in Aspergillus fumigatus. Mol. Microbiol. 2008, 70, 27–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, H. Iron—A Key Nexus in the Virulence of Aspergillus fumigatus. Front. Microbiol. 2012, 3, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blatzer, M.; Binder, U.; Haas, H. The metalloreductase FreB is involved in adaptation of Aspergillus fumigatus to iron starvation. Fungal Genet. Biol. 2011, 48, 1027–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, S.; Torres, G.; Lin, X. Laccases involved in 1,8-dihydroxynaphthalene melanin biosynthesis in Aspergillus fumigatus are regulated by developmental factors and copper homeostasis. Eukaryot. Cell 2013, 12, 1641–1652. [Google Scholar] [CrossRef] [Green Version]
- Sugareva, V.; Hartl, A.; Brock, M.; Hubner, K.; Rohde, M.; Heinekamp, T.; Brakhage, A.A. Characterisation of the laccase-encoding gene abr2 of the dihydroxynaphthalene-like melanin gene cluster of Aspergillus fumigatus. Arch. Microbiol. 2006, 186, 345–355. [Google Scholar] [CrossRef] [Green Version]
- Kues, U.; Ruhl, M. Multiple multi-copper oxidase gene families in basidiomycetes—What for? Curr. Genom. 2011, 12, 72–94. [Google Scholar] [CrossRef] [Green Version]
- Saitoh, Y.; Izumitsu, K.; Morita, A.; Tanaka, C.; Shimizu, K. ChMCO1 of Cochliobolus heterostrophus is a new class of metallo-oxidase, playing an important role in DHN-melanization. Mycoscience 2010, 51, 327–336. [Google Scholar] [CrossRef]
- Tsai, H.F.; Washburn, R.G.; Chang, Y.C.; Kwon-Chung, K.J. Aspergillus fumigatus arp1 modulates conidial pigmentation and complement deposition. Mol. Microbiol. 1997, 26, 175–183. [Google Scholar] [CrossRef]
- Bailão, E.F.L.C.; Lima, P.D.S.; Silva-Bailão, M.G.; Bailão, A.M.; Fernandes, G.D.R.; Kosman, D.J.; Soares, C.M.D.A. Paracoccidioides spp. ferrous and ferric iron assimilation pathways. Front. Microbiol. 2015, 6, 821. [Google Scholar] [CrossRef] [Green Version]
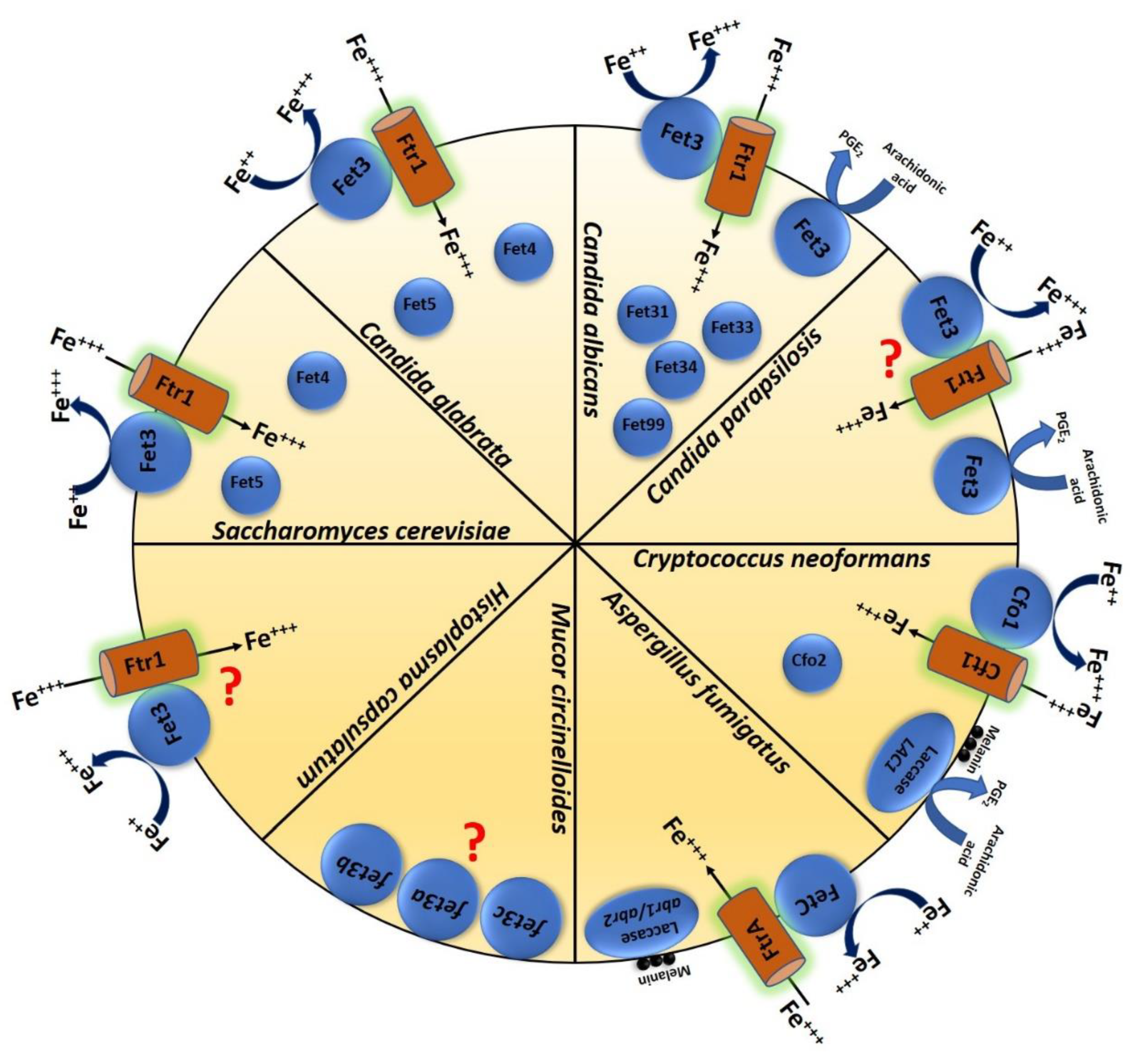
Figure 1.
Schematic image showing important multicopper oxidases in different fungal species such as S. cerevisiae, C. albicans, C. glabrata, C. parapsilosis, A. fumigatus, C. neoformans, M. circinelloides, and H. capsulatum and their function.
Figure 1.
Schematic image showing important multicopper oxidases in different fungal species such as S. cerevisiae, C. albicans, C. glabrata, C. parapsilosis, A. fumigatus, C. neoformans, M. circinelloides, and H. capsulatum and their function.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chakraborty, T.; Tóth, R.; Nosanchuk, J.D.; Gácser, A. Multicopper Oxidases in Saccharomyces cerevisiae and Human Pathogenic Fungi. J. Fungi 2020, 6, 56. https://doi.org/10.3390/jof6020056
AMA Style
Chakraborty T, Tóth R, Nosanchuk JD, Gácser A. Multicopper Oxidases in Saccharomyces cerevisiae and Human Pathogenic Fungi. Journal of Fungi. 2020; 6(2):56. https://doi.org/10.3390/jof6020056
Chicago/Turabian StyleChakraborty, Tanmoy, Renáta Tóth, Joshua D. Nosanchuk, and Attila Gácser. 2020. "Multicopper Oxidases in Saccharomyces cerevisiae and Human Pathogenic Fungi" Journal of Fungi 6, no. 2: 56. https://doi.org/10.3390/jof6020056
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.