
Eutrophication Driven by Aquaculture Fish Farms Controls Phytoplankton and Dinoflagellate Cyst Abundance in the Southern Coastal Waters of Korea

Abstract
:1. Introduction
2. Materials and Methods
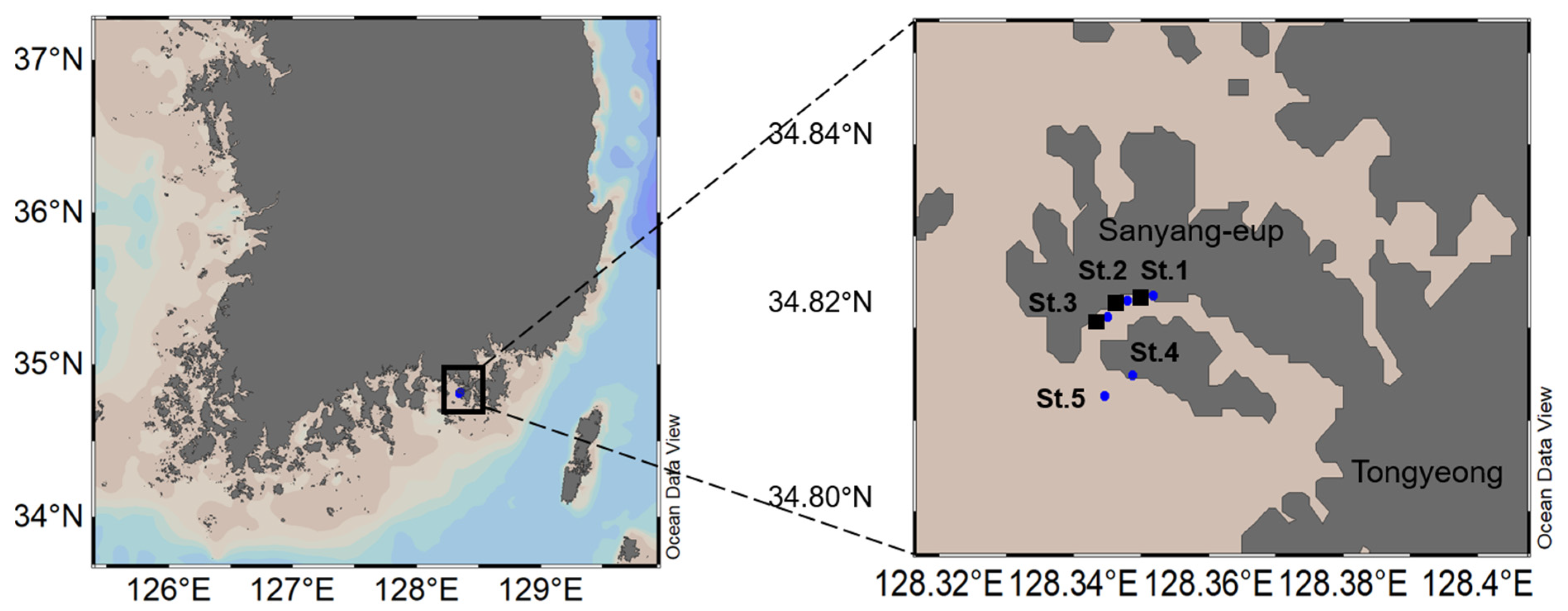
2.1. Study Region
2.2. Field Samples
2.3. Water and Sediment Sample Analysis for Environmental Variables
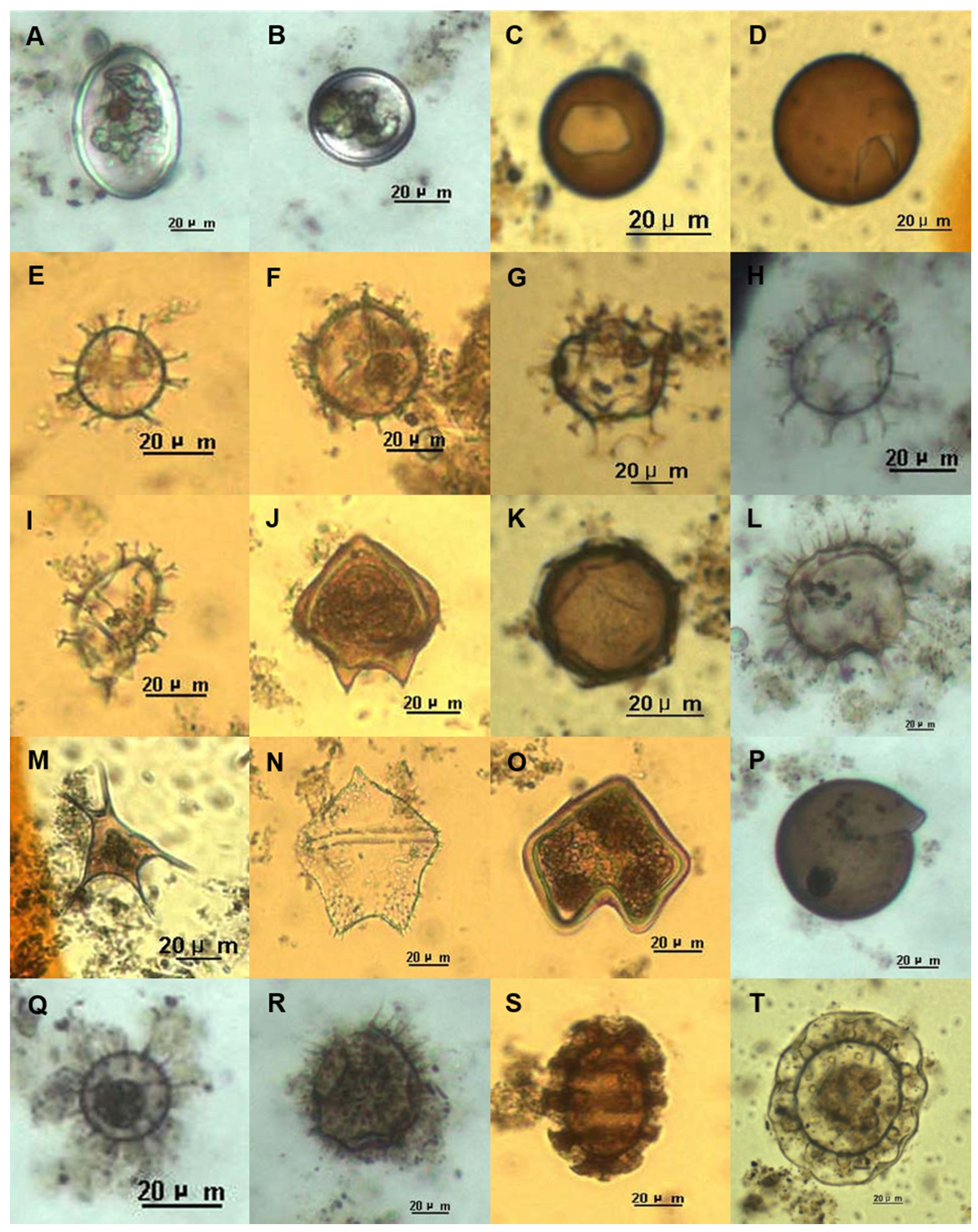
2.4. Phytoplankton and Dinoflagellate Cyst Assemblage Analysis
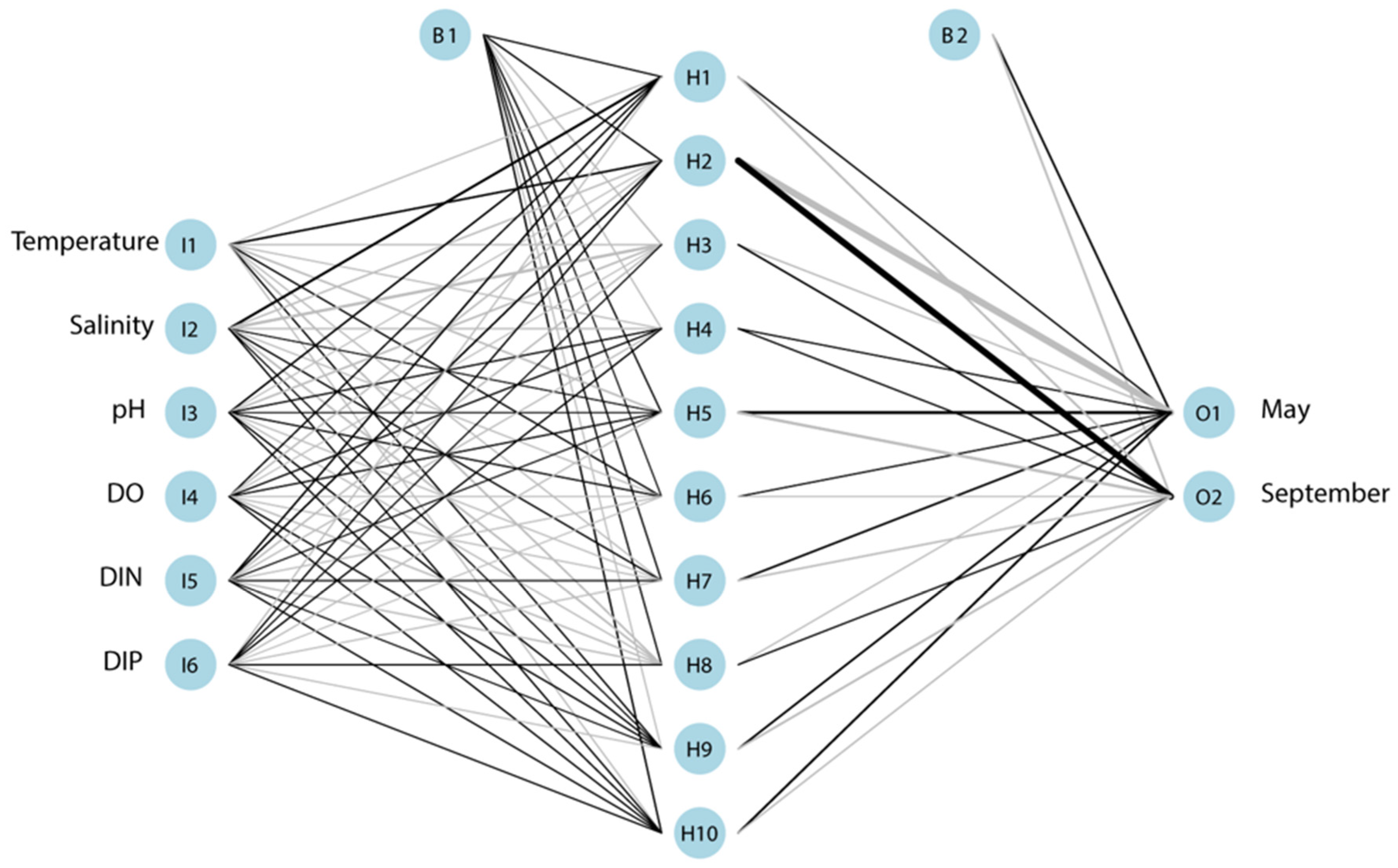
2.5. Data Analysis and Statistical Analysis
3. Results
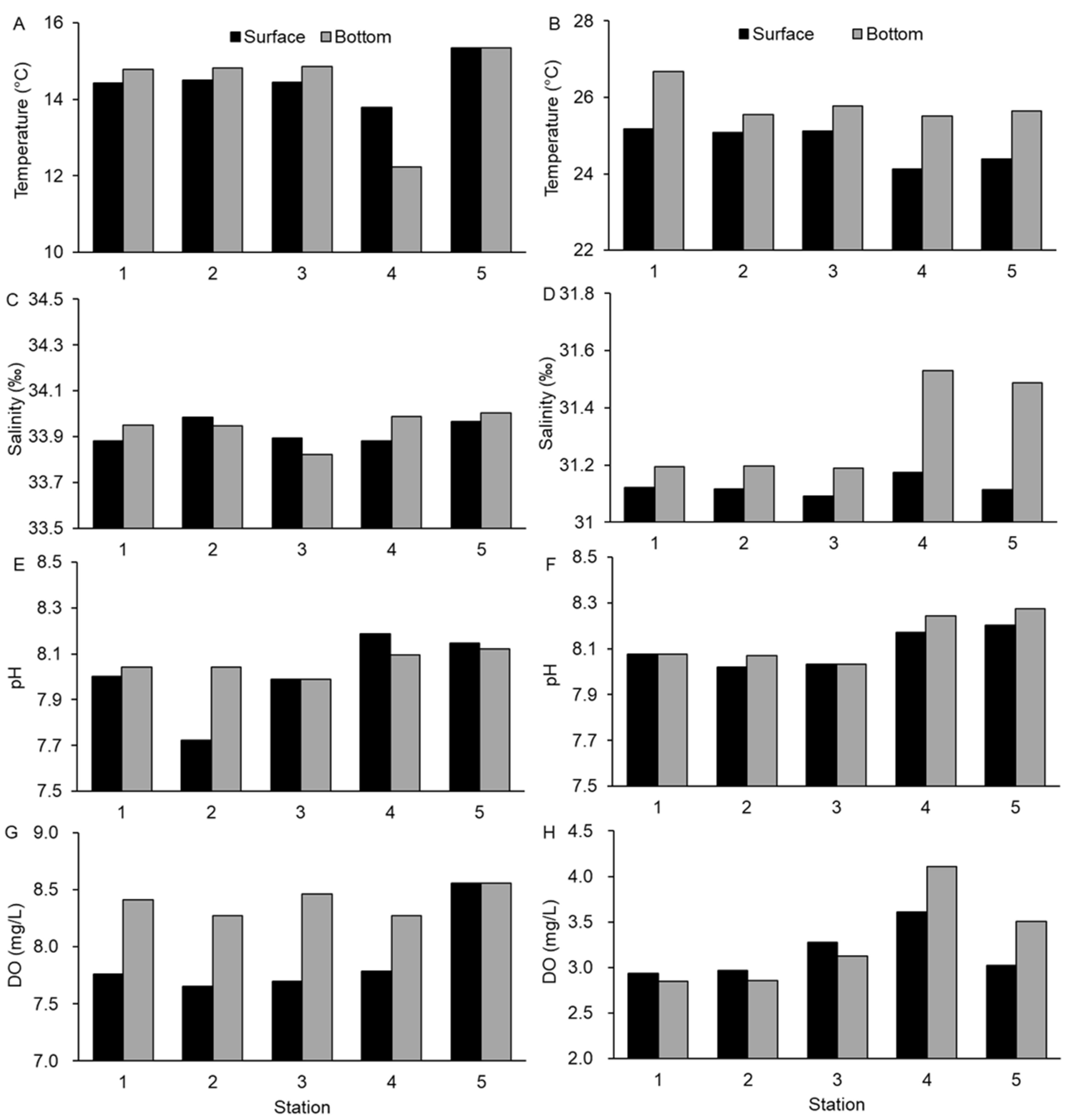
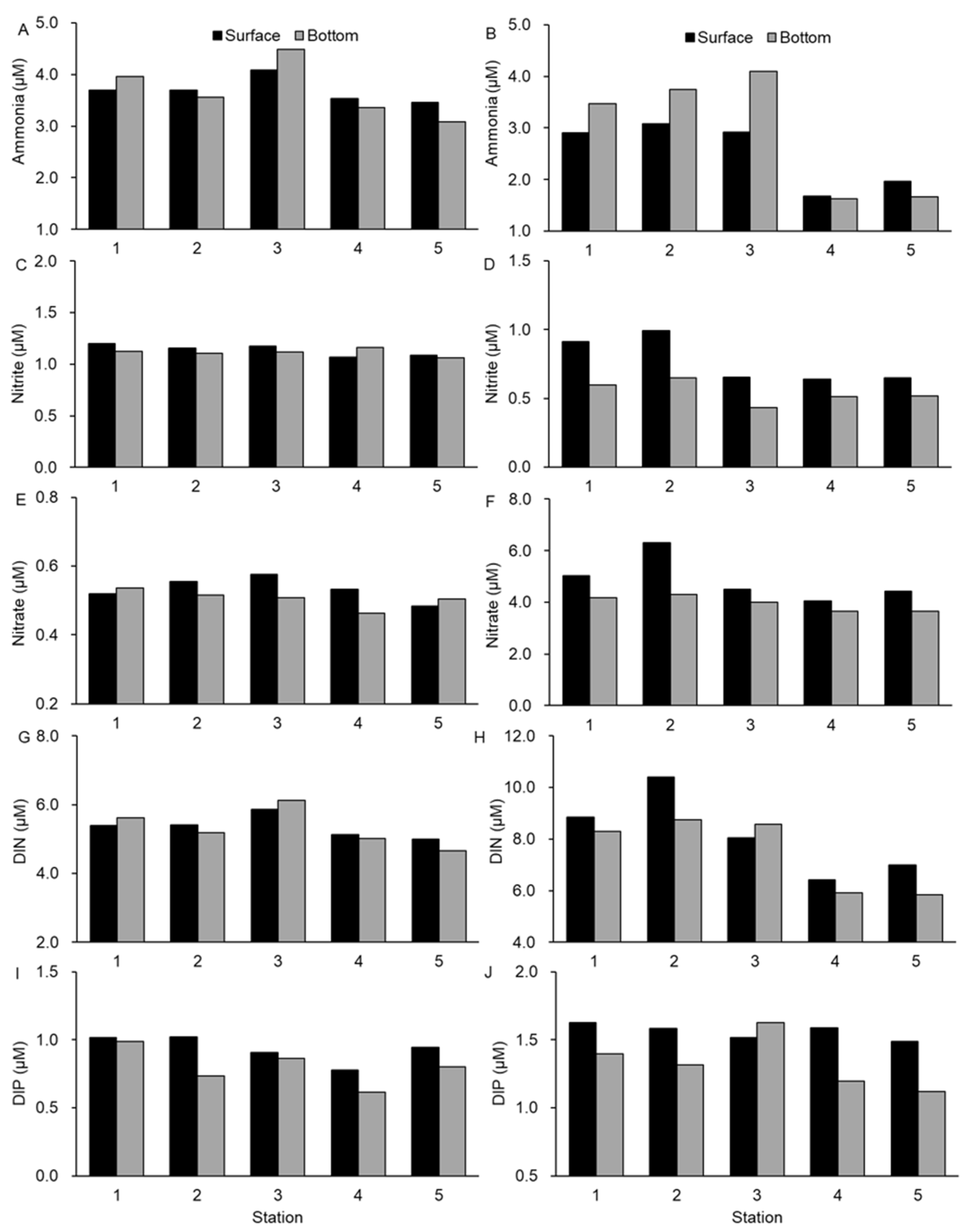
3.1. Environmental Conditions in the Water Column
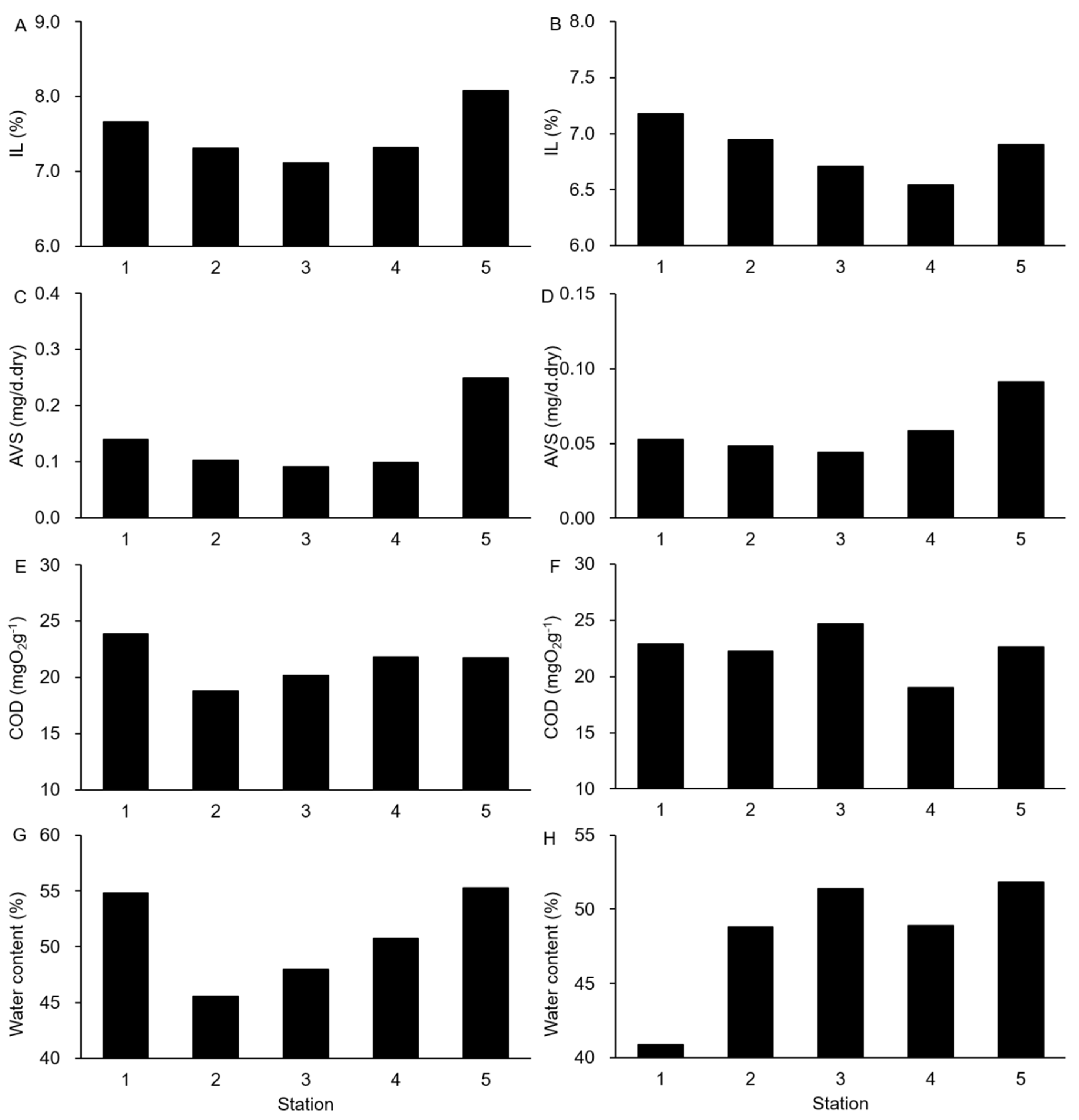
3.2. Environmental Conditions in Sediments
3.3. Characteristics of Phytoplankton Assemblages
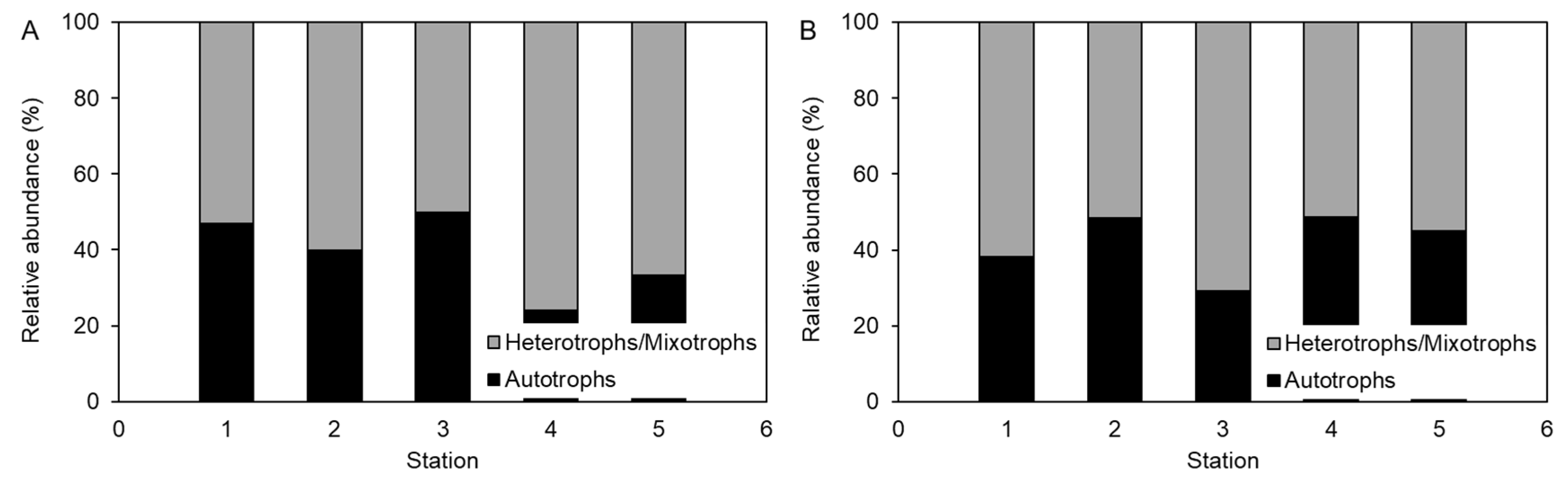
3.4. Characteristics of Dinoflagellate Cyst Assemblages
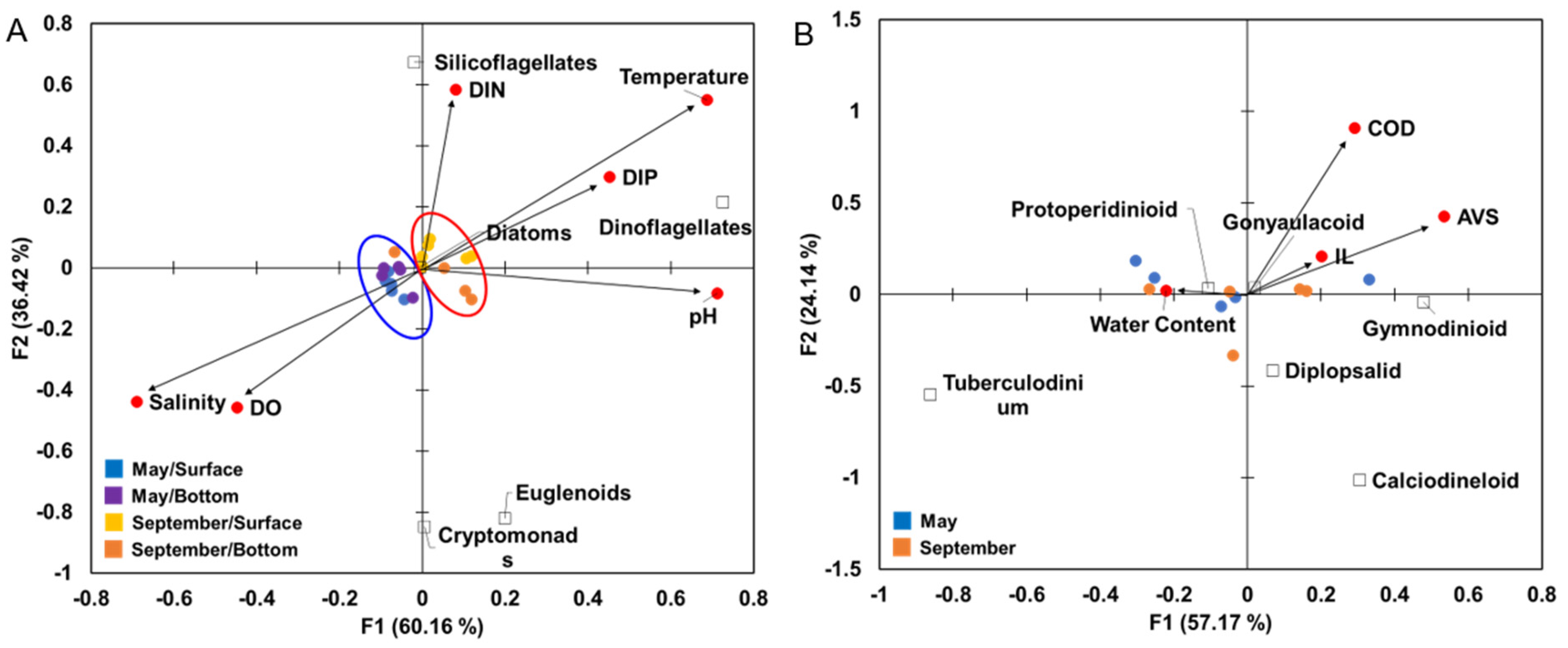
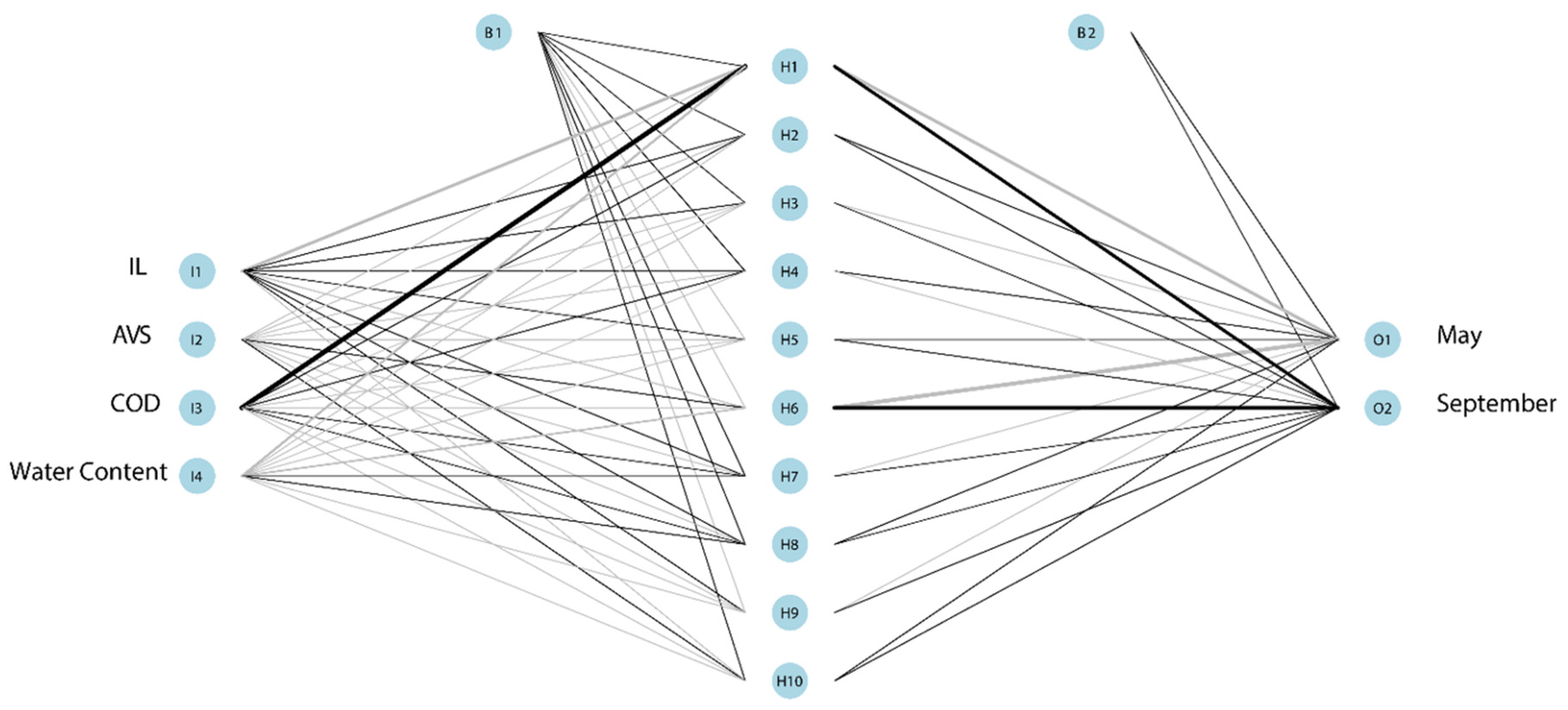
3.5. Relationship between Environmental Variables and Biotic Variables
4. Discussion
4.1. Relationship between Water Quality and Phytoplankton Assemblages
4.2. Relationship between Sediment Environments and Dinoflagellate Cyst Assemblages in Eutrophic Sediments
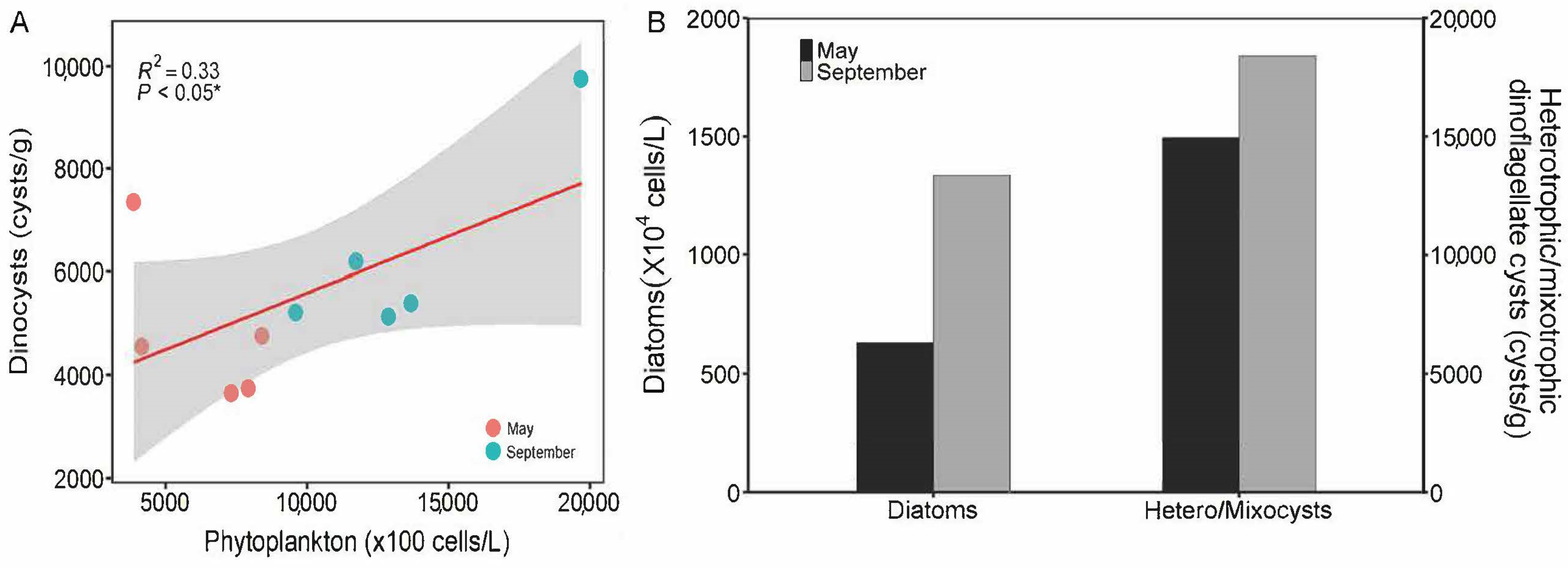
4.3. Relationship between Phytoplankton and Dinoflagellate Cysts
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Dale, B. Dinoflagellate resting cysts: “Benthic plankton”. In Survival Strategies of the Algae; Fryxell, G.A., Ed.; Cambridge University Press: Cambridge, UK, 1983; p. 144. [Google Scholar]
- Anderson, D.M.; Wall, D. Potential importance of benthic cysts of Gonyaulax tamarensis and G. excavata in initiating toxic dinoflagellate blooms. J. Phycol. 1978, 14, 224–234. [Google Scholar] [CrossRef]
- Hallegraeff, G.M.; Bolch, C.J. Transport of toxic dinoflagellate cysts via ships’ ballast water. Mar. Pollut. Bull. 1991, 22, 27–30. [Google Scholar] [CrossRef]
- Matsuoka, K.; Fukuyo, Y. Technical Guide for Modern Dinoflagellate Cyst Study; WESTPAC-HAB, Japan Society for the Promotion of Science: Tokyo, Japan, 2000. [Google Scholar]
- Anderson, D.M.; Coats, D.W.; Tyler, M.A. Encystment of the dinoflagellate Gyrodinium uncatenum: Temperature and nutrient effects. J. Phycol. 1985, 21, 200–206. [Google Scholar] [CrossRef]
- Balch, W.M.; Reid, P.C.; Surrey-Gent, S.C. Spatial and temporal variability of dinoflagellate cyst abundance in a tidal estuary. Can. J. Fish. Aquat. Sci. 1983, 40, s244–s261. [Google Scholar] [CrossRef]
- Kim, Y.-O.; Park, M.-H.; Han, M.-S. Role of cyst germination in the bloom initiation of Alexandrium tamarense (Dinophyceae) in Masan Bay, Korea. Aquat. Microb. Ecol. 2002, 29, 279–286. [Google Scholar] [CrossRef]
- Nehring, S. Dinoflagellate resting cysts from recent German coastal sediments. Bot. Mar. 1997, 40, 307–324. [Google Scholar] [CrossRef]
- Cho, H.-J.; Lee, J.-B.; Moon, C.-H. Dinoflagellate cyst distribution in the surface sediments from the East China sea around Jeju Island. Korean J. Environ. Biol. 2004, 22, 192–199. [Google Scholar]
- Zonneveld, K.A.; Bockelmann, F.; Holzwarth, U. Selective preservation of organic-walled dinoflagellate cysts as a tool to quantify past net primary production and bottom water oxygen concentrations. Mar. Geol. 2007, 237, 109–126. [Google Scholar] [CrossRef]
- Smayda, T. Biogeographical meaning; indicators. In Phytoplankton Manual; Sournia, A., Ed.; UNESCO: Paris, France, 1978; pp. 225–229. [Google Scholar]
- Matsuoka, K.; Kawami, H.; Nagai, S.; Iwataki, M.; Takayama, H. Re-examination of cyst–motile relationships of Polykrikos kofoidii and Polykrikos schwartzii Bütschli (Gymnodiniales, Dinophyceae). Rev. Palaeobot. Palynol. 2009, 154, 79–90. [Google Scholar] [CrossRef]
- Matsuoka, K.; Mizuno, A.; Iwataki, M.; Takano, Y.; Yamatogi, T.; Yoon, Y.H.; Lee, J.-B. Seed populations of a harmful unarmored dinoflagellate Cochlodinium polykrikoides Margalef in the East China Sea. Harmful Algae 2010, 9, 548–556. [Google Scholar] [CrossRef]
- Kang, Y.J.; Ko, T.H.; Lee, J.A.; Lee, J.-B.; Chung, I.K. The community dynamics of phytoplankton and distribution of dinoflagellate cysts in Tongyoung Bay, Korea. Algae 1999, 14, 43–54. [Google Scholar]
- Kim, S.-Y.; Moon, C.-H.; Cho, H.-J.; Lim, D.-I. Dinoflagellate cysts in coastal sediments as indicators of eutrophication: A case of Gwangyang Bay, South Sea of Korea. Estuaries Coasts 2009, 32, 1225–1233. [Google Scholar] [CrossRef]
- Kim, H.-J.; Moon, C.-H.; Cho, H.-J. Spatial-temporal characteristics of dinoflagellate cyst distribution in sediments of Busan Harbor. Sea 2005, 10, 196–203. [Google Scholar]
- Xiao, Y.-Z.; Wang, Z.-H.; Chen, J.-F.; Lu, S.-H.; Qi, Y.-Z. Seasonal dynamics of dinoflagellate cysts in sediments from Daya bay, the south China sea its Relation to the bloom of Scrippsiella trochoidea. Acta Hydrobiol. Sin. 2003, 27, 377–383. [Google Scholar]
- Nehring, S. Dinoflagellate resting cysts as factors in phytoplankton ecology of the North Sea. Helgol. Meeresun 1995, 49, 375–392. [Google Scholar] [CrossRef] [Green Version]
- Dale, B. The sedimentary record of dinoflagellate cysts: Looking back into the future of phytoplankton blooms. Sci. Mar. 2001, 65, 257–272. [Google Scholar] [CrossRef] [Green Version]
- Nehring, S. Scrippsiella spp. resting cysts from the German bight (North Sea): A tool for more complete check-lists of dinoflagellates. Neth. J. Sea Res. 1994, 33, 57–63. [Google Scholar] [CrossRef]
- Zonneveld, K.A.; Dale, B. The cyst-motile stage relationships of Protoperidinium monospinum (Paulsen) Zonneveld et Dale comb. nov. and Gonyaulax verior (Dinophyta, Dinophyceae) from the Oslo Fjord (Norway). Phycologia 1994, 33, 359–368. [Google Scholar] [CrossRef]
- Radi, T.; Pospelova, V.; de Vernal, A.; Vaughn Barrie, J. Dinoflagellate cysts as indicators of water quality and productivity in British Columbia estuarine environments. Mar. Micropaleontol. 2007, 62, 269–297. [Google Scholar] [CrossRef]
- Anglès, S.; Garcés, E.; Hattenrath-Lehmann, T.K.; Gobler, C.J. In situ life-cycle stages of Alexandrium fundyense during bloom development in Northport Harbor (New York, USA). Harmful Algae 2012, 16, 20–26. [Google Scholar] [CrossRef]
- Cremer, H.; Sangiorgi, F.; Wagner-Cremer, F.; McGee, V.; Lotter, A.F.; Visscher, H. Diatoms (Bacillariophyceae) and dinoflagellate cysts (Dinophyceae) from Rookery bay, Florida, USA. Caribb. J. Sci. 2007, 43, 23–58. [Google Scholar] [CrossRef]
- Anderson, D.M.; Burkholder, J.M.; Cochlan, W.P.; Glibert, P.M.; Gobler, C.J.; Heil, C.A.; Kudela, R.M.; Parsons, M.L.; Rensel, J.E.J.; Townsend, D.W.; et al. Harmful algal blooms and eutrophication: Examining linkages from selected coastal regions of the United States. Harmful Algae 2008, 8, 39–53. [Google Scholar] [CrossRef] [Green Version]
- Glibert, P.M.; Al-Azri, A.; Icarus Allen, J.; Bouwman, A.F.; Beusen, A.H.; Burford, M.A.; Harrison, P.J.; Zhou, M. Key questions and recent research advances on harmful algal blooms in relation to nutrients and eutrophication. Glob. Ecol. Oceanogr. Harmful Algal Bloom. 2018, 232, 229–259. [Google Scholar]
- Kang, Y.; Kang, H.-Y.; Kim, D.; Lee, Y.-J.; Kim, T.-I.; Kang, C.-K. Temperature-dependent bifurcated seasonal shift of phytoplankton community composition in the coastal water off southwestern Korea. Ocean Sci. J. 2019, 54, 467–486. [Google Scholar] [CrossRef]
- Kim, B.; Choi, A.; Kim, H.C.; Jung, R.H.; Lee, W.C.; Hyun, J.H. Rate of sulfate reduction an diron reduction in the sediment associated with ablone aquaculture in the southern coastal wateres of Korea. Ocean Polar Res. 2011, 33, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Shim, J.-H.; Kang, Y.-C.; Choi, J.-W. Chemical fluxes at the sediment-water interface below marine fish cages on the coastal waters off Tong-Young, South Coast of Korea. J. Korean Soc. Oceanogr. 1997, 2, 151–159. [Google Scholar]
- Ritz, D.; Lewis, M.; Shen, M. Response to organic enrichment of infaunal macrobenthic communities under salmonid seacages. Mar. Biol. 1989, 103, 211–214. [Google Scholar] [CrossRef]
- Pearson, T.; Rosenberg, R. Macrobenthic succession in relation to organic enrichment and pollution of the marine environment. Oceanogr. Mar. Biol. Annu. Rev. 1978, 16, 229–311. [Google Scholar]
- Park, H.-S.; Choi, J.-W.; Lee, H.-G. Community structure of macrobenthic fauna under marine fish culture cages near Tongyong, Southern Coast of Korea. Korean J. Fish. Aquat. Sci. 2000, 33, 1–8. [Google Scholar]
- Jang, Y.L.; Lee, H.J.; Moon, H.-B.; Lee, W.-C.; Kim, H.C.; Kim, G.B. Marine environmental characteristics in the coastal area surrounding Tongyeong cage-fish farms. J. Korean Soc. Mar. Environ. Energy 2015, 18, 74–80. [Google Scholar] [CrossRef] [Green Version]
- Pospelova, V.; Kim, S.-J. Dinoflagellate cysts in recent estuarine sediments from aquaculture sites of southern South Korea. Mar. Micropaleontol. 2010, 76, 37–51. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lim, W.A.; Jung, C.S.; Park, J. Spatial distributions and monthly variations of water quality in coastal seawater of Tongyeong, Korea. J. Korean Soc. Mar. Environ. Eng. 2011, 14, 154–162. [Google Scholar] [CrossRef]
- Jones, M.N. Nitrate reduction by shaking with cadmium—Alternative to cadmium columns. Water Res. 1984, 18, 643–646. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Chemical and Biological Methods for Seawater Analysis; Pergamon: Oxford, UK, 1984. [Google Scholar]
- Price, N.M.; Harrison, P.J. Comparison of methods for the analysis of dissolved urea in seawater. Mar. Biol. 1987, 94, 307–317. [Google Scholar] [CrossRef]
- Heiri, O.; Lotter, A.F.; Lemcke, G. Loss on ignition as a method for estimating organic and carbonate content in sediments: Reproducibility and comparability of results. J. Paleolimnol. 2001, 25, 101–110. [Google Scholar] [CrossRef]
- Bowman, G.T.; Delfino, J.J. Sediment oxygen demand techniques: A review and comparison of laboratory and in situ systems. Water Res. 1980, 14, 491–499. [Google Scholar] [CrossRef]
- KME. Water Quality Standards Handbook; Korean Ministry of Environment: Sejong, Korea, 2000; pp. 99–208.
- Rickard, D.; Morse, J.W. Acid volatile sulfide (AVS). Mar. Chem. 2005, 97, 141–197. [Google Scholar] [CrossRef]
- Okaichi, T. The cause of red-tide in neritic water. Jpn. Fish. Resour. Conserv. Assoc. 1985, 58–75. [Google Scholar]
- Kim, D.; Lim, D.-I.; Jeon, S.-K.; Jung, H.-S. Chemical characteristics and eutrophication in Cheonsu Bay, West Coast of Korea. Ocean Polar Res. 2005, 27, 45–58. [Google Scholar]
- Kim, H.-J.; Yeong Park, J.; Ho Son, M.; Moon, C.-H. Long-term variations of phytoplankton community in coastal waters of Kyoungju city area. J. Fishries Mar. Sci. Educ. 2016, 28, 1417–1434. [Google Scholar] [CrossRef] [Green Version]
- Shim, J.H. Ilustrated Encyclopedia of Fauna and Flora of Korea Vol.34 Marine Phytoplankton; Shin, J.H., Ed.; Korean Ministry of Education: Sejong, Korea, 1994.
- Tomas, C.R. Identifying Marine Phytoplankton; Elsevier: Amsterdam, The Netherlands, 1997. [Google Scholar]
- Bolch, C.; Hallegraeff, G. Dinoflagellate cysts in recent marine sediments from Tasmania, Australia. Bot. Mar. 1990, 33, 173–192. [Google Scholar] [CrossRef]
- Günther, F.; Fritsch, S. neuralnet: Training of neural networks. R J. 2010, 2, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S-PLUS; Springer Science & Business Media: Berlin, Germany, 2013. [Google Scholar]
- Bergmeir, C.N.; Benítez Sánchez, J.M. Neural networks in R using the Stuttgart neural network simulator: RSNNS. In Proceedings of the American Statistical Association, San Diego, CA, USA, 28 July–2 August 2012. [Google Scholar]
- Paruelo, J.; Tomasel, F. Prediction of functional characteristics of ecosystems: A comparison of artificial neural networks and regression models. Ecol. Model. 1997, 98, 173–186. [Google Scholar] [CrossRef]
- Olden, J.D.; Jackson, D.A. Illuminating the “black box”: A randomization approach for understanding variable contributions in artificial neural networks. Ecol. Model. 2002, 154, 135–150. [Google Scholar] [CrossRef]
- Rumelhart, D.E.; Hinton, G.E.; Williams, R.J. Learning representations by back-propagating errors. Nature 1986, 323, 533–536. [Google Scholar] [CrossRef]
- Lee, J.H.; Huang, Y.; Dickman, M.; Jayawardena, A.W. Neural network modelling of coastal algal blooms. Ecol. Model. 2003, 159, 179–201. [Google Scholar] [CrossRef]
- Beck, M.W. NeuralNetTools: Visualization and analysis tools for neural networks. J. Stat. Softw. 2018, 85, 1–20. [Google Scholar] [CrossRef]
- Olden, J.D. An artificial neural network approach for studying phytoplankton succession. Hydrobiologia 2000, 436, 131–143. [Google Scholar] [CrossRef]
- Millie, D.F.; Weckman, G.R.; Paerl, H.W.; Pinckney, J.L.; Bendis, B.J.; Pigg, R.J.; Fahnenstiel, G.L. Neural net modeling of estuarine indicators: Hindcasting phytoplankton biomass and net ecosystem production in the Neuse (North Carolina) and Trout (Florida) Rivers, USA. Ecol. Indic. 2006, 6, 589–608. [Google Scholar] [CrossRef]
- Millie, D.F.; Weckman, G.R.; Pigg, R.J.; Tester, P.A.; Dyble, J.; Wayne Litaker, R.; Carrick, H.J.; Fahnenstiel, G.L. Modeling phytoplankton abundance in Saginaw Bay, Lake Huron: Using artificial neural networks to discern functional influence of environmental variables and relevance to a Great Lakes observing system. J. Phycol. 2006, 42, 336–349. [Google Scholar] [CrossRef]
- Song, E.-S.; Lim, J.-S.; Chang, N.-I.; Sin, Y.-S. Relative importance of bottom-up vs. top-down controls on size-structured phytoplankton dynamics in a freshwater ecosystem: II. Investigation of controlling factors using statistical modeling analysis. Korean J. Ecol. Environ. 2005, 38, 445–453. [Google Scholar]
- Banse, K. Cell volumes, maximal growth rates of unicellular algae and ciliates, and the role of ciliates in the marine pelagial 1, 2. Limnol. Oceanogr. 1982, 27, 1059–1071. [Google Scholar] [CrossRef]
- Hecky, R.; Kilham, P. Nutrient limitation of phytoplankton in freshwater and marine environments: A review of recent evidence on the effects of enrichment 1. Limnol. Oceanogr. 1988, 33, 796–822. [Google Scholar] [CrossRef] [Green Version]
- Furnas, M.J. In situ growth rates of marine phytoplankton: Approaches to measurement, community and species growth rates. J. Plankton Res. 1990, 12, 1117–1151. [Google Scholar] [CrossRef]
- Smayda, T.J. Harmful algal blooms: Their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol. Oceanogr. 1997, 42, 1137–1153. [Google Scholar] [CrossRef]
- Gobler, C.J.; Berry, D.L.; Anderson, O.R.; Burson, A.; Koch, F.; Rodgers, B.S.; Moore, L.K.; Goleski, J.A.; Allam, B.; Bowser, P.; et al. Characterization, dynamics, and ecological impacts of harmful Cochlodinium polykrikoides blooms on eastern Long Island, NY, USA. Harmful Algae 2008, 7, 293–307. [Google Scholar] [CrossRef]
- Gobler, C.J.; Burson, A.; Koch, F.; Tang, Y.Z.; Mulholland, M.R. The role of nitrogenous nutrients in the occurrence of harmful algal blooms caused by Cochlodinium polykrikoides in New York estuaries (USA). Harmful Algae 2012, 17, 64–74. [Google Scholar] [CrossRef]
- Zhang, G.; Liang, S.; Shi, X.; Han, X. Dissolved organic nitrogen bioavailability indicated by amino acids during a diatom to dinoflagellate bloom succession in the Changjiang River estuary and its adjacent shelf. Mar. Chem. 2015, 176, 83–95. [Google Scholar] [CrossRef]
- Marañón, E.; Cermeño, P.; Huete-Ortega, M.; López-Sandoval, D.C.; Mouriño-Carballido, B.; Rodríguez-Ramos, T. Resource supply overrides temperature as a controlling factor of marine phytoplankton growth. PLoS ONE 2014, 9, e99312. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.K.; Aranguren-Gassis, M.; Kremer, C.T.; Gould, M.R.; Anderson, K.; Klausmeier, C.A.; Litchman, E. Temperature–nutrient interactions exacerbate sensitivity to warming in phytoplankton. Glob. Chang. Biol. 2017, 23, 3269–3280. [Google Scholar] [CrossRef]
- Pomeroy, L.R.; Wiebe, W.J. Temperature and substrates as interactive limiting factors for marine heterotrophic bacteria. Aquat. Microb. Ecol. 2001, 23, 187–204. [Google Scholar] [CrossRef] [Green Version]
- Lumb, C.M. Basic Concepts Concerning Assessments of Environmental Effects of Marine Fish Farms; Håkanson, L., Ervik, A., Makinen, T., Moller, B., Eds.; Council of Ministers: Copenhagen, Denmark, 1988; p. 103.
- Ackefors, H.; Enell, M. Discharge of nutrients from Swedish fish farming to adjacent sea areas. Ambio 1990, 19, 28–35. [Google Scholar]
- Kang, C.-K.; PARK, P.-Y.L.-J.-S.; KIM, P.-J. On the distribution of organic matter in the nearshore surface sediment of Korea. Bull. Korean Fish. Soc 1993, 26, 557–566. [Google Scholar]
- Yoon, Y. A study on the distributional characteristic of organic matters on the surface sediments and its origin in Keogeum-sudo, southern part of Korean Peninsula. J. Korean Environ. Sci. Soc. 2000, 9, 137–144. [Google Scholar]
- Kim, H.-S.; Matsuoka, K. Process of eutrophication estimated by dinoflagellate cyst assemblages in Omura Bay, Kyushu, West Japan. Bull. Plankton Soc. Jpn. 1998, 45, 133–147. [Google Scholar]
- Matsuoka, K. Eutrophication process recorded in dinoflagellate cyst assemblages—A case of Yokohama Port, Tokyo Bay, Japan. Sci. Total Environ. 1999, 231, 17–35. [Google Scholar] [CrossRef]
- Anderson, D.M. Dinoflagellate cyst dynamics in coastal and estuarine water. In Toxic Dinoflagellates; Anderson, D.M., Ed.; Elsevier: New York, NY, USA, 1985; pp. 219–224. [Google Scholar]
- Lee, J.-B.; Kim, D.Y.; Lee, J. Community dynamics and distribution of dinoflagellates and their cysts in Masan-Chinhae Bay, Korea. Fish. Aquat. Sci. 1998, 1, 283–292. [Google Scholar]
- Lee, M.H.; Lee, J.-B.; Lee, J.A.; Park, J.G. Community structure of flagellates and dynamics of resting cysts in Kamak Bay, Korea. Algae 1999, 14, 255–266. [Google Scholar]
- Park, J.S.; Yoon, Y.H.; Noh, I.H. Estimation on the variation of marine environment by the distribution of organic matter and dinoflagellate cyst in the vertical sediments in Gamak Bay, Korea. J. Korean Soc. Mar. Environ. Eng. 2004, 7, 164–173. [Google Scholar]
- Shin, H.H.; Yoon, Y.H.; Matsuoka, K. Modern dinoflagellate cysts distribution off the eastern part of Geoje Island, Korea. Ocean Sci. J. 2007, 42, 31–39. [Google Scholar] [CrossRef]
- Lee, J.-S.; Shin, I.-S.; Kim, Y.-M.; Chang, D.-S. Paralytic shellfish toxins in the museel, Mytilus edulis, caused the shellfish poisoning accident at Geoje, Korea in 1996. J. Korean Fish. Soc. 1997, 30, 158–160. [Google Scholar]
- Baek, S.H.; Choi, J.M.; Lee, M.; Park, B.S.; Zhang, Y.; Arakawa, O.; Takatani, T.; Jeon, J.-K.; Kim, Y.O. Change in paralytic shellfish toxins in the mussel Mytilus galloprovincialis depending on dynamics of harmful Alexandrium catenella (Group I) in the Geoje coast (South Korea) during bloom season. Toxins 2020, 12, 442. [Google Scholar] [CrossRef]
- Chang, D.-S.; Shin, I.-S.; Pyeun, J.-H.; Park, Y.-H. A Study on paralytic shellfish poison of sea mussel, Mytilus edulis. Food poisoning accident in Gamchun Bay, Pusan, Korea. Korean J. Fish. Aqua. Sci. 1986, 20, 293–299. [Google Scholar]
- Shin, H.H.; Yoon, Y.H.; Kawami, H.; Iwataki, M.; Matsuoka, K. The first appearance of toxic dinoflagellate Alexandrium tamarense (Gonyaulacales, Dinophyceae) responsible for the PSP contaminations in Gamak Bay, Korea. Algae 2008, 23, 251–255. [Google Scholar] [CrossRef]
- Hattenrath, T.K.; Anderson, D.M.; Gobler, C.J. The influence of anthropogenic nitrogen loading and meteorological conditions on the dynamics and toxicity of Alexandrium fundyense blooms in a New York (USA) estuary. Harmful Algae 2010, 9, 402–412. [Google Scholar] [CrossRef]
- Gaines, G.; Taylor, F. Extracellular digestion in marine dinoflagellates. J. Plankton Res. 1984, 6, 1057–1061. [Google Scholar] [CrossRef]
- Jacobson, D.M.; Anderson, D.M. Thecate heterophic dinoflagellates: Feeding behavior and mechanisms. J. Phycol. 1986, 22, 249–258. [Google Scholar] [CrossRef]
- Hansen, P.J. Prey size selection, feeding rates and growth dynamics of heterotrophic dinoflagellates with special emphasis on Gyrodinium Spirale. Mar. Biol. 1992, 114, 327–334. [Google Scholar] [CrossRef]
- Buskey, E.J. Behavioral components of feeding selectivity of the heterotrophic dinoflagellate Protoperidinium pellucidum. Mar. Ecol. Prog. Ser. 1997, 153, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Harrison, P.; Fulton, J.; Taylor, F.; Parsons, T. Review of the biological oceanography of the Strait of Georgia: Pelagic environment. Can. J. Fish. Aquat. Sci. 1983, 40, 1064–1094. [Google Scholar] [CrossRef]
- Fujii, R.; Matsuoka, K. Seasonal change of dinoflagellates cyst flux collected in a sediment trap in Omura Bay, West Japan. J. Plankton Res. 2006, 28, 131–147. [Google Scholar] [CrossRef] [Green Version]
- Hamel, D.; de Vernal, A.; Gosselin, M.; Hillaire-Marcel, C. Organic-walled microfossils and geochemical tracers: Sedimentary indicators of productivity changes in the North Water and northern Baffin Bay during the last centuries. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 5277–5295. [Google Scholar] [CrossRef]
- Marret, F.; Zonneveld, K.A. Atlas of modern organic-walled dinoflagellate cyst distribution. Rev. Palaeobot. Palynol. 2003, 125, 1–200. [Google Scholar] [CrossRef]
- Radi, T.; de Vernal, A. Dinocyst distribution in surface sediments from the northeastern Pacific margin (40–60 N) in relation to hydrographic conditions, productivity and upwelling. Rev. Palaeobot. Palynol. 2004, 128, 169–193. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species\Stations | Surface | Bottom | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | |
| Diatoms | ||||||||||
| Amphiprora sp. | 12 | |||||||||
| Asterionella glacialis | 321 | 384 | 144 | 96 | 192 | 288 | 204 | 312 | 192 | 180 |
| Chaetoceros affinis | 4788 | 8940 | 7800 | 2976 | 3132 | 5172 | 3024 | 3504 | 2400 | 1224 |
| Chaetoceros compressus | 264 | 96 | ||||||||
| Chaetoceros constrictus | 216 | 324 | 480 | 312 | 348 | 504 | 540 | 540 | ||
| Chaetoceros curvisetus | 144 | 96 | ||||||||
| Chaetoceros danicus | 36 | 12 | ||||||||
| Chaetoceros debilis | 96 | 252 | 228 | 84 | 144 | 372 | 120 | 300 | 72 | 36 |
| Chaetoceros didymus | 132 | 600 | 768 | 300 | 204 | 516 | 300 | 912 | 360 | 72 |
| Chaetoceros laciniosus | 132 | |||||||||
| Chaetoceros socialis | 36 | 48 | ||||||||
| Chaetoceros sp. | 180 | 120 | 60 | 96 | 12 | |||||
| Cylindrotheca closterium | 72 | 12 | 12 | 12 | 12 | 24 | 36 | 24 | ||
| Ditylum brightwellii | 12 | 12 | ||||||||
| Guinardia delicatula | 12 | 288 | ||||||||
| Leptocylindrus danicus | 60 | |||||||||
| Licmophora sp. | 12 | |||||||||
| Navicula sp. | 12 | 12 | 24 | 12 | 36 | 12 | 36 | 36 | ||
| Odontella longicruris | 360 | 480 | 24 | 132 | ||||||
| Paralia sulcata | 300 | |||||||||
| Pleurosigma angulataum | 72 | 12 | 108 | 120 | 72 | 36 | 72 | 132 | 36 | |
| Pseudo-nitzschia sp. | 288 | 468 | 120 | 144 | 204 | 60 | 96 | 72 | ||
| Pseudo-nitzschia spp.1 | 48 | 36 | 60 | 180 | 168 | |||||
| Pseudo-nitzschia spp.2 | 72 | 120 | 216 | 24 | 132 | 24 | 12 | |||
| Rhizosolenia setigera | 12 | 36 | 12 | 24 | ||||||
| Skeletonema costatum | 120 | 108 | 252 | 648 | 72 | 228 | 180 | |||
| Thalassionema nitzschioides | 96 | 48 | 72 | 96 | ||||||
| Thalassiosira rotula | 24 | 48 | ||||||||
| Thalassiosira sp. | 36 | 12 | 48 | |||||||
| Dinoflagellates | ||||||||||
| Gyrodinium spirale | 12 | 12 | 12 | |||||||
| Scrippsiella trochoidea | 12 | |||||||||
| Cryptomonads | ||||||||||
| Chroomonas sp. | 204 | 12 | 12 | 36 | 12 | 12 | ||||
| Total | 7653 | 11,304 | 10,056 | 3876 | 5604 | 7092 | 4596 | 6744 | 4452 | 2160 |
| Species\Stations | Surface | Bottom | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 | |
| Diatoms | ||||||||||
| Amphiprora sp. | 12 | |||||||||
| Asterionella glacialis | 276 | 96 | 96 | 72 | 108 | 84 | 48 | 96 | 24 | 96 |
| Bacteriastrum delicatulum | 1404 | 264 | 696 | 132 | 528 | 684 | 120 | 516 | 180 | 84 |
| Chaetoceros affinis | 2028 | 1800 | 684 | 876 | 468 | 120 | 108 | 324 | 168 | |
| Chaetoceros brevis | 108 | 84 | ||||||||
| Chaetoceros compressus | 3024 | 2256 | 1380 | 684 | 1068 | 2928 | 1488 | 1920 | 528 | 324 |
| Chaetoceros constrictus | 780 | 24 | 512 | 120 | 96 | |||||
| Chaetoceros curvisetus | 4488 | 2016 | 4164 | 2472 | 6288 | 2532 | 3120 | 2352 | 2508 | 3936 |
| Chaetoceros danicus | 24 | 36 | 36 | 24 | 120 | 60 | ||||
| Chacetoceros debilis | 516 | 60 | 108 | 84 | 288 | 168 | 396 | 48 | 144 | 120 |
| Chaetoceros decipiens | 252 | |||||||||
| Chaetoceros didymus | 2940 | 1800 | 1272 | 924 | 972 | 1692 | 1104 | 1236 | 1260 | 564 |
| Chaetoceros laciniosus | 4308 | 1584 | 1680 | 732 | 1380 | 1812 | 1884 | 1200 | 2700 | 420 |
| Chaetoceros lorenzianus | 552 | 480 | 228 | 360 | 120 | 420 | 492 | 348 | 216 | 228 |
| Chaetoceros socialis | 240 | 336 | ||||||||
| Chaetoceros sp. | 1188 | 648 | 588 | 72 | 360 | 240 | 96 | 324 | 336 | 96 |
| Cylindrotheca closterium | 36 | 480 | 48 | 48 | 96 | 36 | 24 | 60 | ||
| Dactyliosolen fragilissimus | 180 | 768 | 1032 | 336 | 456 | 804 | 396 | 396 | 264 | 360 |
| Ditylum brightwellii | 72 | 132 | 240 | 96 | 204 | 120 | 96 | 96 | 36 | 72 |
| Eucampia zodiacus | 24 | 12 | 12 | |||||||
| Grammatophora sp. | 24 | |||||||||
| Guinardia delicatula | 60 | 84 | 240 | 72 | 132 | 36 | 132 | 12 | ||
| Guinardia flaccida | 72 | 48 | 36 | 24 | 48 | 48 | 12 | |||
| Guinardia striata | 48 | 96 | 84 | 96 | 36 | 24 | 12 | 12 | ||
| Guinardia sp. | 108 | 48 | ||||||||
| Hemiaulus membranaceus | 24 | |||||||||
| Lauderia borialis | 48 | 24 | 48 | 72 | ||||||
| Leptocylindrus danicus | 2016 | 684 | 528 | 84 | 36 | 1104 | 444 | 660 | 120 | 240 |
| Leptocylindrus minimus | 156 | 204 | 156 | 108 | 96 | 96 | ||||
| Licmophora sp. | ||||||||||
| Navicula sp. | 24 | 24 | 240 | 24 | 36 | 36 | 24 | 72 | 36 | 72 |
| Odontella longicruris | 48 | 156 | 120 | 132 | 72 | 180 | 108 | 132 | 192 | 192 |
| Paralia sulcata | 72 | 240 | ||||||||
| Pleurosigma angulatum | 12 | 12 | 12 | 12 | 12 | 12 | 12 | |||
| Pleurosigma elongatum | 12 | 12 | ||||||||
| Pleurosigma sp. | 12 | |||||||||
| Proboscia alata | 12 | 24 | 12 | 12 | 24 | |||||
| Pseudo-nitzschia sp. | 216 | 24 | 36 | 96 | 24 | 120 | 240 | 240 | ||
| Pseudo-nitzschia spp.1 | 216 | 84 | 588 | 492 | 492 | 132 | 276 | 156 | 252 | 36 |
| Pseudo-nitzschia spp.2 | 300 | 168 | 192 | 216 | 300 | 240 | 84 | 216 | ||
| Rhizosolenia imbricata | 24 | |||||||||
| Rhizosolenia setigera | 12 | 12 | 12 | |||||||
| Rhizosolenia sp. | 24 | 12 | 12 | |||||||
| Skeletonema costatum | 108 | 228 | 156 | 60 | 360 | 348 | 240 | 216 | ||
| Stephanopyxis turris | 48 | 48 | 24 | 36 | 36 | |||||
| Thalassionema nitzschioides | 360 | 156 | 336 | 132 | 36 | 84 | 216 | 600 | 228 | |
| Thalassiosira sp. | 48 | |||||||||
| Thalassiothrix fraudenfeldii | 84 | 24 | 24 | 132 | 48 | 60 | 132 | |||
| Thalassiothrix sp. | ||||||||||
| Tropidoneis lepidoptera | 12 | |||||||||
| Dinoflagellates | ||||||||||
| Alexandrium sp. | 12 | |||||||||
| Chaettonella antiqua | 12 | |||||||||
| Chaetonella sp. | 12 | |||||||||
| Gymnodinium sp. | 24 | 12 | ||||||||
| Gyrodinium spirale | 72 | 60 | 48 | 36 | 12 | 12 | ||||
| Gyrodinium sp. | 12 | 12 | ||||||||
| Heterocapsa triquetra | 12 | |||||||||
| Heterosigma akashiwo | 36 | 36 | 48 | 24 | ||||||
| Margalefidinium polykrikoides | 12 | |||||||||
| Noctiluca scintillanse | 12 | |||||||||
| Prorocentrum micans | 12 | 12 | 24 | 12 | ||||||
| Prorocentrum minimum | 72 | 12 | 240 | |||||||
| Prorocentrum sp. | 12 | 12 | 24 | 12 | ||||||
| Prorocentrum triestinum | 24 | 24 | 12 | |||||||
| Protoperidinium pacificum | 12 | 12 | ||||||||
| Protoperidinium sp. | 12 | 12 | 12 | 48 | 12 | 36 | 36 | 12 | 12 | |
| Scrippsiella trochoidea | 48 | 12 | 12 | 12 | ||||||
| Cryptomonads | ||||||||||
| Chroomonas sp. | 12 | 60 | 156 | 24 | 12 | |||||
| Euglenoids | ||||||||||
| Eutreptiella gymnastica | 48 | 48 | 24 | |||||||
| Silico-flagellates | ||||||||||
| Dictyocha fibula | 12 | |||||||||
| Total | 24,732 | 15,432 | 14,904 | 8556 | 14,748 | 14,652 | 11,936 | 10,932 | 10,644 | 8712 |
| Species\Stations | May | September | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Paleontological Name | Biological Name | 1 | 2 | 3 | 4 | 5 | 1 | 2 | 3 | 4 | 5 |
| AUTOTROPHS | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Calciodineloid group | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Scrippsiella trochoidea | Scrippsiella trochoidea | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 200 |
| Scrippsiella sp. | Scrippsiella sp. | 0 | 0 | 0 | 0 | 0 | 160 | 0 | 0 | 0 | 0 |
| Gonyaulacoid group | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Alexandrium affine | Alexandrium affine | 0 | 0 | 0 | 0 | 0 | 680 | 1400 | 420 | 1140 | 1380 |
| Alexandrium tamarense/catenella | Alexandrium tamarense/catenella | 0 | 0 | 0 | 0 | 0 | 0 | 400 | 220 | 200 | 600 |
| Alexandrium sp. | Alexandrium sp. | 0 | 0 | 180 | 220 | 0 | 0 | 0 | 0 | 200 | 200 |
| Linglodinium machaerophorum | Linglodinium polyedrum | 0 | 0 | 0 | 0 | 0 | 160 | 0 | 0 | 0 | 0 |
| Spiniferites bentori | Gonyaulax digitalis | 0 | 180 | 740 | 220 | 0 | 0 | 0 | 420 | 380 | 0 |
| Spiniferites bulloideus | Gonyaulax scrippsea | 1220 | 1140 | 740 | 640 | 1960 | 340 | 200 | 220 | 200 | 600 |
| Spiniferites membranaceus | Gonyaulax spinifera complex | 480 | 0 | 360 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Spiniferites mirabilis | Gonyaulax spinifera | 0 | 0 | 0 | 0 | 0 | 340 | 0 | 0 | 0 | 0 |
| Spiniferites ramosus | Gonyaulax spinifera | 0 | 0 | 360 | 0 | 0 | 0 | 0 | 0 | 0 | 200 |
| Spiniferites sp. | Gonyaulax sp. | 0 | 0 | 0 | 0 | 500 | 500 | 200 | 220 | 380 | 1180 |
| Tuberculodinium group | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Tuberculodinium vacampoae | Pyrophacus steinii | 0 | 180 | 0 | 0 | 0 | 160 | 400 | 0 | 0 | 0 |
| HETEROTROPHS/MIXOTROPHS | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Diplopsalid group | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Diplopsalis sp. | Diplopsalis sp. | 0 | 0 | 360 | 0 | 0 | 0 | 200 | 0 | 0 | 0 |
| Oblea acantocysta | Diplopsalis parva | 0 | 0 | 0 | 0 | 240 | 340 | 0 | 220 | 200 | 0 |
| Gymnodinioid group | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Margalefidinium sp. | Margalefidinium sp. | 0 | 0 | 0 | 0 | 240 | 0 | 0 | 220 | 200 | 200 |
| Gymnodinium catenatum | Gymnodinium catenatum | 0 | 0 | 0 | 0 | 0 | 500 | 0 | 0 | 0 | 600 |
| Gymnodinium sp. | Gymnodinium sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 200 | 0 |
| Polykrikos hartmannii | Polykrikos hartmannii | 0 | 0 | 0 | 0 | 500 | 160 | 0 | 0 | 200 | 200 |
| Polykrikos kofoidii/schwartzii complex | Polykrikos kofoidii/schwartzii complex | 240 | 0 | 560 | 220 | 740 | 160 | 200 | 0 | 380 | 0 |
| Protoperidinioid group | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Brigantedinium caracoense | Protoperidinium avellanum | 0 | 940 | 180 | 640 | 980 | 680 | 600 | 220 | 0 | 400 |
| Brigantedinium simplex | Protoperidinium conicoides | 0 | 0 | 0 | 220 | 0 | 160 | 0 | 0 | 0 | 0 |
| Brigantedinium sp. | Protoperidinium sp. | 980 | 1120 | 920 | 1300 | 1720 | 680 | 1000 | 1920 | 1140 | 2180 |
| Protoperidinium americanum | Protoperidinium pellucidum | 0 | 0 | 0 | 0 | 240 | 0 | 0 | 0 | 0 | 600 |
| Protoperidinium sp. | Protoperidinium sp. | 240 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Quinquecuspis concreta | Protoperidinium leonis | 240 | 0 | 0 | 220 | 0 | 0 | 600 | 0 | 200 | 400 |
| Selenopemphix quanta | Protoperidinium conicum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 640 | 0 | 200 |
| Selenopemphix sp. | Protoperidinium sp. | 240 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Stelladinium reidii | Protoperidinium compressum | 0 | 0 | 0 | 440 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trinovantedinium applanatum | Protoperidinium pentagonum | 0 | 0 | 0 | 220 | 0 | 680 | 200 | 0 | 0 | 200 |
| Votadinium calvum | Protoperdinium oblongum | 0 | 180 | 360 | 220 | 240 | 500 | 0 | 420 | 200 | 400 |
| Total | 3640 | 3740 | 4760 | 4560 | 7360 | 6200 | 5400 | 5140 | 5220 | 9740 | |
| Station | May | August | ||
|---|---|---|---|---|
| Surface | Bottom | Surface | Bottom | |
| 1 | 3.35 | 2.22 | 5.90 | 5.24 |
| 2 | 4.16 | 1.57 | 7.50 | 4.53 |
| 3 | 3.60 | 2.02 | 4.86 | 6.78 |
| 4 | 2.27 | 1.13 | 4.04 | 2.76 |
| 5 | 2.78 | 1.38 | 3.57 | 2.61 |
| Region | May | September | |
|---|---|---|---|
| DIN (μM) | Tongyeong offshore | 2.11 | 4.11 |
| Geoje | 2.46 | 4.50 | |
| Sachoen | 3.00 | 4.79 | |
| Study region | 5.32 * | 7.76 * | |
| DIP (μM) | Tongyeong offshore | 0.42 | 0.35 |
| Geoje | 0.40 | 0.26 | |
| Sachoen | 0.39 | 0.37 | |
| Study region | 0.86 * | 1.44 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, Y.; Kim, H.-J.; Moon, C.-H. Eutrophication Driven by Aquaculture Fish Farms Controls Phytoplankton and Dinoflagellate Cyst Abundance in the Southern Coastal Waters of Korea. J. Mar. Sci. Eng. 2021, 9, 362. https://doi.org/10.3390/jmse9040362
Kang Y, Kim H-J, Moon C-H. Eutrophication Driven by Aquaculture Fish Farms Controls Phytoplankton and Dinoflagellate Cyst Abundance in the Southern Coastal Waters of Korea. Journal of Marine Science and Engineering. 2021; 9(4):362. https://doi.org/10.3390/jmse9040362
Chicago/Turabian StyleKang, Yoonja, Hyun-Jung Kim, and Chang-Ho Moon. 2021. "Eutrophication Driven by Aquaculture Fish Farms Controls Phytoplankton and Dinoflagellate Cyst Abundance in the Southern Coastal Waters of Korea" Journal of Marine Science and Engineering 9, no. 4: 362. https://doi.org/10.3390/jmse9040362