
Characterization of Dental Pulp Stem Cell Responses to Functional Biomaterials Including Mineralized Trioxide Aggregates


Abstract
:1. Introduction
2. Methods
2.1. Study Approval
2.2. Cell Culture
2.3. RNA Isolation
2.4. PCR Screening
2.5. Proliferation Assays
2.6. Viability Assays
2.7. Experimental Factors
2.8. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Name |
| DPSC | Dental pulp stem cell |
| VEGF | vascular endothelial growth factor |
| BMP | bone morphogenic protein |
| MTA | mineralized trioxide aggregates |
| SHED | stem cells from human exfoliated deciduous teeth |
| SCAP | stem cells from the apical papilla |
| UNLV | University of Nevada, Las Vegas |
| OPRS | Office for the Protection of Research Subjects |
| SDM | School of Dental Medicine |
| DMSO | dimethyl sulfoxide |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| FBS | fetal bovine serum |
| rDT | rapid doubling time |
| iDT | intermediate doubling time |
| sDT | slow doubling time |
| PCR | polymerase chain reaction |
| ISCT | International Society for Cellular Therapy |
| MSC | mesenchymal stem cell |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| bFGF | basic fibroblast growth factor |
| FGF | fibroblast growth factor |
References
- Ahmed, G.M.; Abouauf, E.A.; Abubakr, N.; Dörfer, C.E.; El-Sayed, K.F. Tissue Engineering Approaches for Enamel, Dentin, and Pulp Regeneration: An Update. Stem Cells Int. 2020, 2020, 5734539. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.-L.; Liu, W.; Wu, Y.-M.; Sun, W.-L.; Dörfer, C.E.; El-Sayed, K.M.F. Oral Mesenchymal Stem/Progenitor Cells: The Immunomodulatory Masters. Stem Cells Int. 2020, 2020, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Tsutsui, T.W. Dental Pulp Stem Cells: Advances to Applications. Stem Cells Cloning Adv. Appl. 2020, 13, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Mao, J.; Liu, Y. Pulp stem cells derived from human permanent and deciduous teeth: Biological characteristics and therapeutic applications. Stem Cells Transl. Med. 2020, 9, 445–464. [Google Scholar] [CrossRef] [Green Version]
- Yamada, Y.; Nakamura-Yamada, S.; Kusano, K.; Baba, S. Clinical Potential and Current Progress of Dental Pulp Stem Cells for Various Systemic Diseases in Regenerative Medicine: A Concise Review. Int. J. Mol. Sci. 2019, 20, 1132. [Google Scholar] [CrossRef] [Green Version]
- Mendi, A.; Ulutürk, H.; Ataç, M.S.; Yılmaz, D. Stem Cells for the Oromaxillofacial Area: Could they be a promising source for regeneration in dentistry? Adv. Exp. Med. Biol. 2019, 1144, 101–121. [Google Scholar] [CrossRef] [PubMed]
- Aydin, S.; Şahin, F. Stem Cells Derived from Dental Tissues. Adv. Exp. Med. Biol. 2019, 1144, 123–132. [Google Scholar] [CrossRef]
- Berebichez-Fridman, R.; Montero-Olvera, P.R. Sources and Clinical Applications of Mesenchymal Stem Cells: State-of-the-art review. Sultan Qaboos Univ. Med. J. 2018, 18, 264–277. [Google Scholar] [CrossRef] [Green Version]
- Baniebrahimi, G.; Khanmohammadi, R.; Mir, F. Teeth-derived stem cells: A source for cell therapy. J. Cell. Physiol. 2019, 234, 2426–2435. [Google Scholar] [CrossRef]
- Cui, D.; Li, H.; Wan, M.; Peng, Y.; Xu, X.; Zhou, X.; Zheng, L. The Origin and Identification of Mesenchymal Stem Cells in Teeth: from Odontogenic to Non-odontogenic. Curr. Stem Cell Res. Ther. 2017, 13, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Macrin, D.; Joseph, J.P.; Pillai, A.A.; Devi, A. Eminent Sources of Adult Mesenchymal Stem Cells and Their Therapeutic Imminence. Stem Cell Rev. Rep. 2017, 13, 741–756. [Google Scholar] [CrossRef]
- Avinash, K.; Malaippan, S.; Dooraiswamy, J.N. Methods of Isolation and Characterization of Stem Cells from Different Regions of Oral Cavity Using Markers: A Systematic Review. Int. J. Stem Cells 2017, 10, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Rodas-Junco, B.A.; Villicaña, C. Dental Pulp Stem Cells: Current Advances in Isolation, Expansion and Preservation. Tissue Eng. Regen. Med. 2017, 14, 333–347. [Google Scholar] [CrossRef]
- Perlea, P.; Rusu, M.C.; Didilescu, A.C.; Pătroi, E.F.; Leonardi, R.M.; Imre, M.; Răducanu, A.M. Phenotype heterogeneity in dental pulp stem niches. Rom. J. Morphol. Embryol. Rev. Roum. Morphol. Embryol. 2016, 57, 1187–1193. [Google Scholar]
- Conde, M.C.M.; Chisini, L.A.; Grazioli, G.; Francia, A.; De Carvalho, R.V.; Alcázar, J.C.B.; Tarquinio, S.B.C.; Demarco, F.F. Does Cryopreservation Affect the Biological Properties of Stem Cells from Dental Tissues? A Systematic Review. Braz. Dent. J. 2016, 27, 633–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrelli, M.; Codispoti, B.; Shelton, R.M.; Scheven, B.A.; Cooper, P.R.; Tatullo, M.; Paduano, F. Dental Pulp Stem Cell Mechanoresponsiveness: Effects of Mechanical Stimuli on Dental Pulp Stem Cell Behavior. Front. Physiol. 2018, 9, 1685. [Google Scholar] [CrossRef]
- An, S. The emerging role of extracellular Ca2+ in osteo/odontogenic differentiation and the involvement of intracellular Ca2+ signaling: From osteoblastic cells to dental pulp cells and odontoblasts. J. Cell. Physiol. 2019, 234, 2169–2193. [Google Scholar] [CrossRef]
- Kim, S.G.; Zhou, J.; Solomon, C.; Zheng, Y.; Suzuki, T.; Chen, M.; Song, S.; Jiang, N.; Cho, S.; Mao, J.J. Effects of Growth Factors on Dental Stem/Progenitor Cells. Dent. Clin. N. Am. 2012, 56, 563–575. [Google Scholar] [CrossRef] [Green Version]
- Tóth, F.; Gáll, J.M.; Tőzsér, J.; Hegedűs, C. Effect of inducible bone morphogenetic protein 2 expression on the osteogenic differentiation of dental pulp stem cells in vitro. Bone 2020, 132, 115214. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Xu, W.; Li, J.; Chen, Y.; Pan, Y.; Wu, B. Effects of vascular endothelial growth factor and insulin growth factor-1 on proliferation, migration, osteogenesis and vascularization of human carious dental pulp stem cells. Mol. Med. Rep. 2019, 20, 3924–3932. [Google Scholar] [CrossRef]
- Javid, B.; Panahandeh, N.; Torabzadeh, H.; Nazarian, H.; Parhizkar, A.; Asgary, S. Bioactivity of endodontic biomaterials on dental pulp stem cells through dentin. Restor. Dent. Endod. 2020, 45, e3. [Google Scholar] [CrossRef]
- Dahake, P.T.; Panpaliya, N.P.; Kale, Y.J.; Dadpe, M.V.; Kendre, S.B.; Bogar, C. Response of stem cells from human exfoliated deciduous teeth (SHED) to three bioinductive materials—An in vitro experimental study. Saudi Dent. J. 2020, 32, 43–51. [Google Scholar] [CrossRef]
- Petta, T.D.M.; Pedroni, A.C.F.; Saavedra, D.F.; Faial, K.D.C.F.; Marques, M.M.; Couto, R.S.D. The effect of three different pulp capping cements on mineralization of dental pulp stem cells. Dent. Mater. J. 2020, 39, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Tomlin, A.; Sanders, M.B.; Kingsley, K. The effects of cryopreservation on human dental pulp-derived mesenchymal stem cells. Biomater. Biomech. Bioeng. 2016, 3, 105–114. [Google Scholar] [CrossRef]
- Shoff, M.; Booker, T.; Leavitt, B.; Harmon, D.; Kingsley, K.; Howard, K.M. Differential exosome miRNA expression in oral cancer stem cells. ExRNA 2020, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tomlin, A.; Nelson, B.; Kingsley, K. Dental Pulp Stem Cell Biomarkers for Cellular ViabilityFollowing Long-Term Cryopreserva-tion. Int. J. Cell Syst. Dev. Biol. 2018, 1, 1–6. [Google Scholar]
- Cinelli, J.; Nguyen, E.; Kingsley, K. Assessment of Dental Pulp Stem Cell (DPSC) Biomarkers Following Induction with Bone Morphogenic Protein 2 (BMP-2). J. Adv. Biol. Biotechnol. 2018, 19, 1–12. [Google Scholar] [CrossRef]
- Cinelli, J.; Mullins, E.; Kingsley, K. Differential Effects of Bone Morphogenic Protein (BMP) and Vascular Endothelial Growth Factor (VEGF) on Dental Pulp Stem Cell (DPSC) Subpopulations. EC Dent. Sci. 2020, 19, 01–10. [Google Scholar]
- Forgues, C.; Kingsley, K. Effects of Vascular Endothelial Growth Factor (VEGF) Alone and in Combination on Rapidly Dividing Dental Pulp Stem Cells (DPSC). EC Dent. Sci. 2019, 18, 576–582. [Google Scholar]
- Mullins, E.; Forgues, C.; Kingsley, K. Effects of Vascular Endothelial Growth Factor (VEGF) on Dental Pulp Stem Cells (DPSC). Curr. Res. Med. 2019, 9, 1–8. [Google Scholar] [CrossRef]
- Whiting, M.; Hatton, W.J.; Kingsley, K. Pluripotent Stem Cell Markers and microRNA Expression May Correlate with Dental Pulp Stem Cell Viability and Proliferation Rates. Asclepius Med. Res. Rev. 2019, 2, 1–7. [Google Scholar]
- Young, A.; Kingsley, K. Dental Pulp Stem Cells: A review of factors that influence the therapeutic potential of stem cell iso-lates. Biomater. Biomed. Eng. 2015, 2, 61–69. [Google Scholar] [CrossRef]
- Pisal, R.V.; Suchanek, J.; Siller, R.; Soukup, T.; Hrebikova, H.; Bezrouk, A.; Kunke, D.; Micuda, S.; Filip, S.; Sullivan, G.; et al. Directed reprogramming of comprehensively characterized dental pulp stem cells extracted from natal tooth. Sci. Rep. 2018, 8, 6168. [Google Scholar] [CrossRef] [PubMed]
- Ferro, F.; Spelat, R.; D’Aurizio, F.; Puppato, E.; Pandolfi, M.; Beltrami, A.P.; Cesselli, D.; Falini, G.; Beltrami, C.A.; Curcio, F. Dental Pulp Stem Cells Differentiation Reveals New Insights in Oct4A Dynamics. PLoS ONE 2012, 7, e41774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aksel, H.; Huang, G.T.-J. Combined Effects of Vascular Endothelial Growth Factor and Bone Morphogenetic Protein 2 on Odonto/Osteogenic Differentiation of Human Dental Pulp Stem Cells In Vitro. J. Endod. 2017, 43, 930–935. [Google Scholar] [CrossRef] [PubMed]
- D’ Alimonte, I.; Nargi, E.; Mastrangelo, F.; Falco, G.; Lanuti, P.; Marchisio, M.; Miscia, S.; Robuffo, I.; Capogreco, M.; Buccella, S.; et al. Vascular endothelial growth factor enhances in vitro proliferation and osteogenic differenti-ation of human dental pulp stem cells. J. Biol. Regul. Homeost. Agents 2011, 25, 57–69. [Google Scholar] [PubMed]
- Xie, H.; Dubey, N.; Shim, W.; Ramachandra, C.; Min, K.; Cao, T.; Rosa, V. Functional Odontoblastic-Like Cells Derived from Human iPSCs. J. Dent. Res. 2017, 97, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Feng, G.; Zhang, J.; Xing, J.; Huang, D.; Lian, M.; Zhang, W.; Wu, W.; Hu, Y.; Lu, X.; et al. Basic fibroblast growth factor promotes human dental pulp stem cells cultured in 3D porous chitosan scaffolds to neural differentiation. Int. J. Neurosci. 2020, 5, 1–9. [Google Scholar] [CrossRef]
- Yuan, X.; Liu, M.; Cao, X.; Yang, S. Ciliary IFT80 regulates dental pulp stem cells differentiation by FGF/FGFR1 and Hh/BMP2 signaling. Int. J. Biol. Sci. 2019, 15, 2087–2099. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Xu, S.; Dao, J.; Gan, Z.; Zeng, X. Differential expression of lncRNA/miRNA/mRNA and their related functional networks during the osteogenic/odontogenic differentiation of dental pulp stem cells. J. Cell. Physiol. 2020, 235, 3350–3361. [Google Scholar] [CrossRef]
- Shi, R.; Yang, H.; Lin, X.; Cao, Y.; Zhang, C.; Fan, Z.; Hou, B. Analysis of the characteristics and expression profiles of coding and noncoding RNAs of human dental pulp stem cells in hypoxic conditions. Stem Cell Res. Ther. 2019, 10, 89. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Zhong, Y.; Kong, Y.; Chen, Y.; Feng, J.; Zheng, J. Lineage-specific exosomes promote the odontogenic differentiation of human dental pulp stem cells (DPSCs) through TGFβ1/smads signaling pathway via transfer of microRNAs. Stem Cell Res. Ther. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]

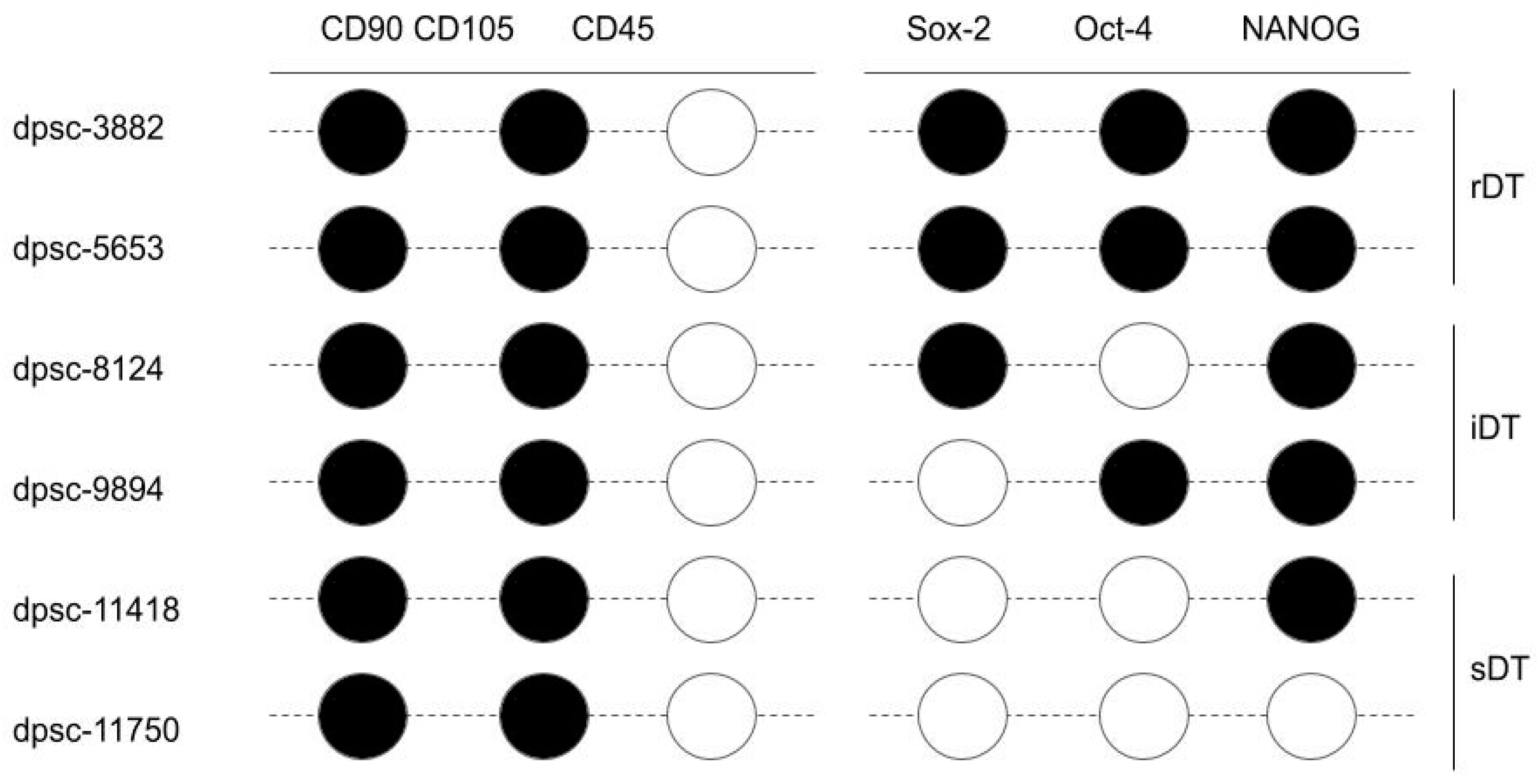
 indicates RT-PCR band intensity above the limit of detection;
indicates RT-PCR band intensity above the limit of detection;  indicates an RT-PCR result below the limit of detection.
indicates RT-PCR band intensity above the limit of detection; indicates an RT-PCR result below the limit of detection.
indicates an RT-PCR result below the limit of detection.
indicates RT-PCR band intensity above the limit of detection; indicates an RT-PCR result below the limit of detection.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DPSC Isolate | RNA Quantification | Statistical Analysis | RNA Purity |
|---|---|---|---|
| dpsc-3882 | 921.1 ng/μL | – | 1.74 |
| dpsc-5653 | 887.3 ng/μL | – | 1.77 |
| dpsc-8124 | 925.6 ng/μL | – | 1.72 |
| dpsc-9894 | 913.2 ng/μL | – | 1.65 |
| dpsc-11418 | 879.4 ng/μL | – | 1.91 |
| dpsc-11750 | 910.1 ng/μL | – | 1.83 |
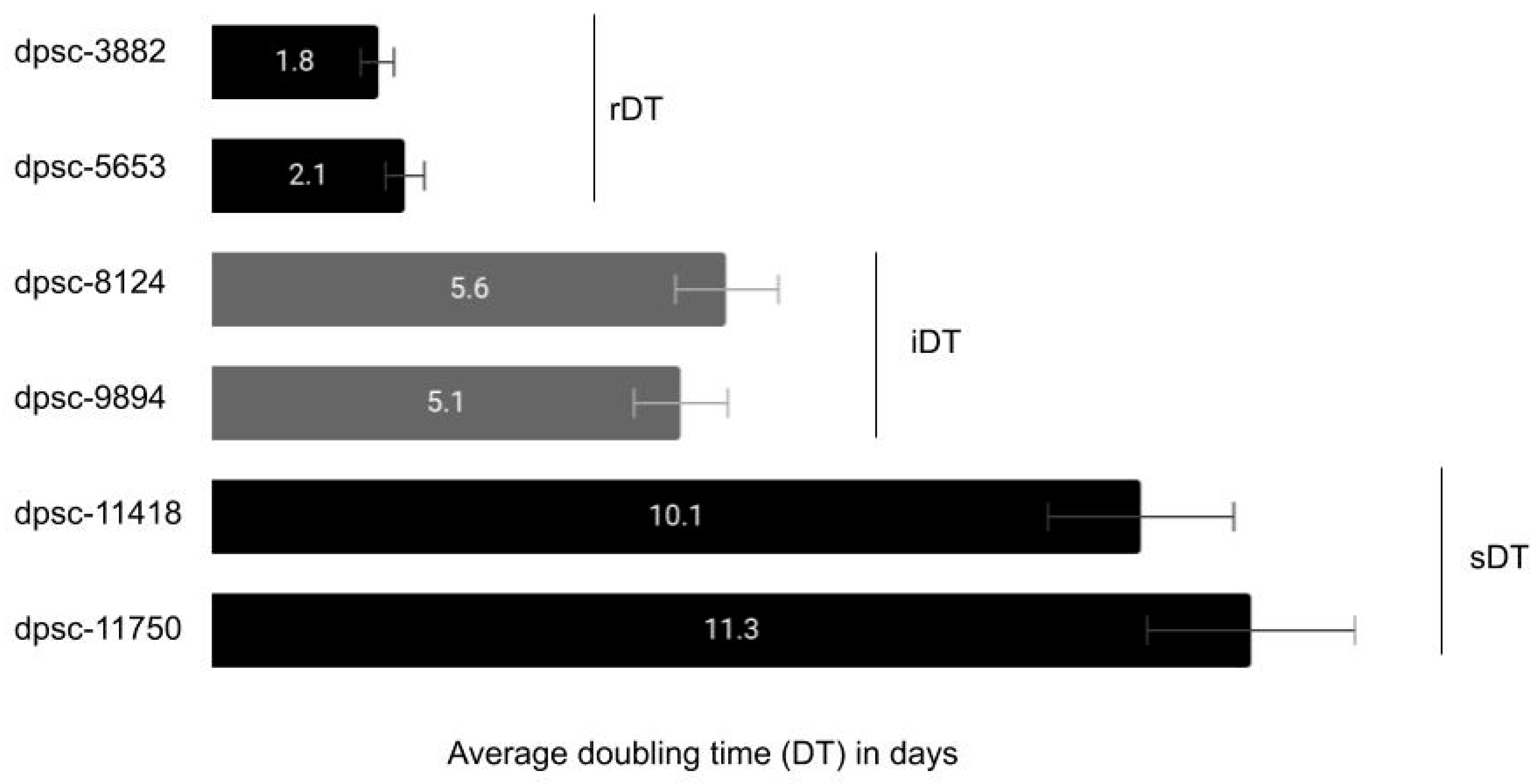
| rDT | 904.2 ng/μL | X2 = 0.325 | ave. = 1.75 |
| iDT | 919.4 ng/μL | d.f. = 2 | ave. = 1.69 |
| sDT | 894.75 ng/μL | p = 0.8502 | ave. = 1.87 |
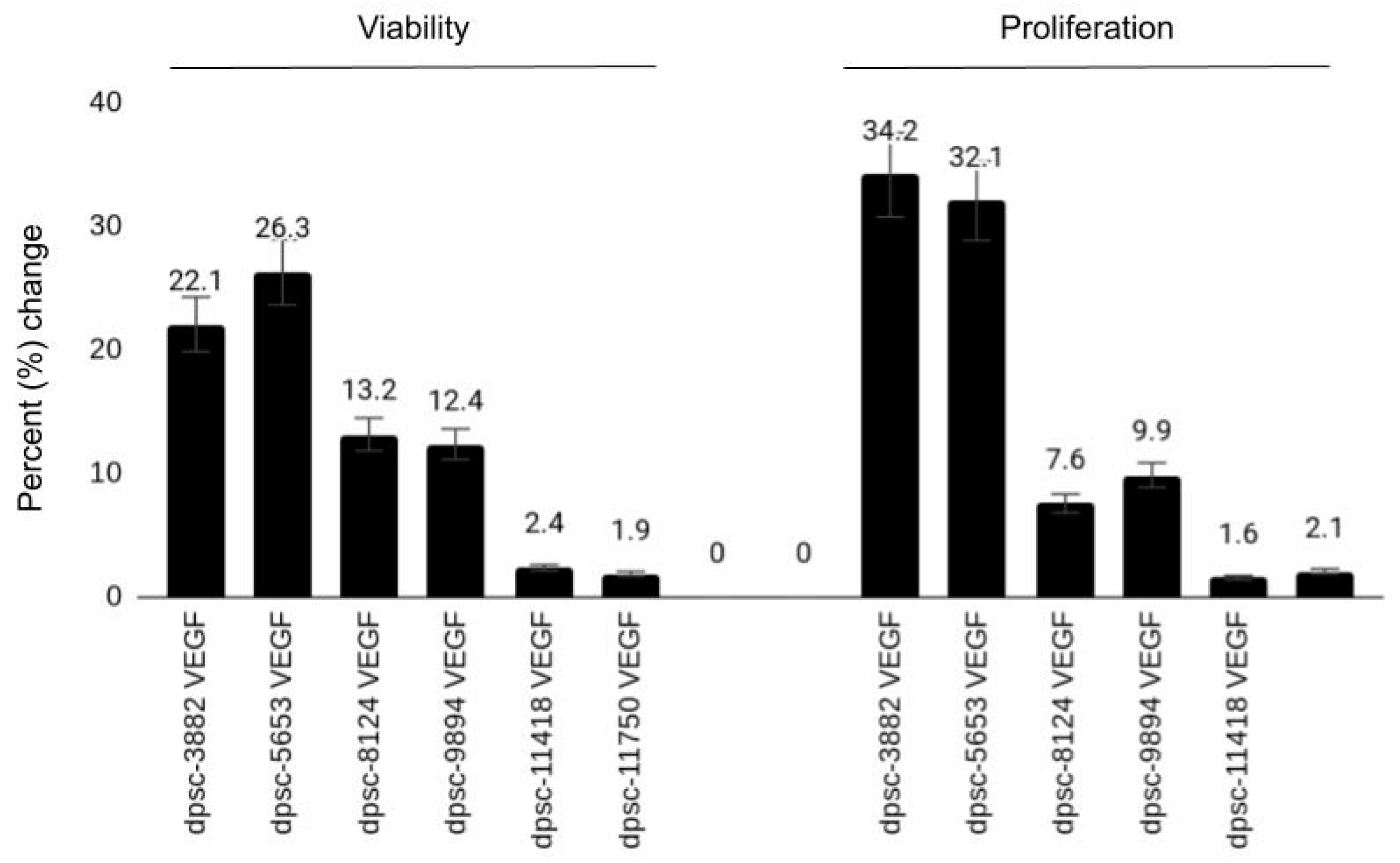
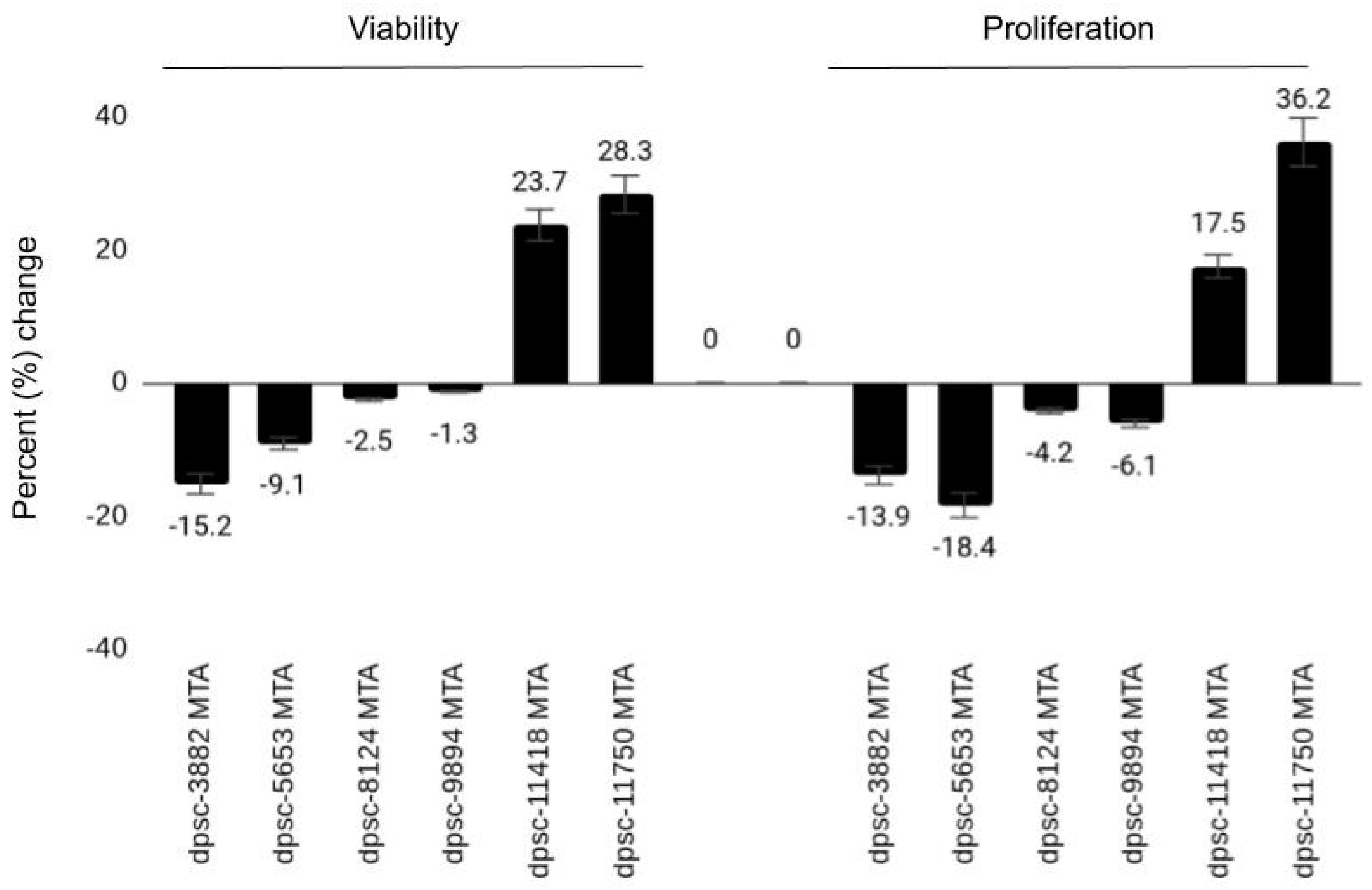
| DPSC Isolate | VEGF (Viability, Growth) | BMP-2 (Viability, Growth) | MTA (Viability, Growth) |
|---|---|---|---|
| dpsc-3882 | 22.1%, 34.2% | 1.2%, 2.4% | −15.2%, −13.9% |
| dpsc-5653 | 26.3%, 32.1% | 2.2%, 3.1% | −9.1%, −18.4% |
| dpsc-8124 | 13.2%, 7.6% | 3.5%, 2.2% | −2.5%, −4.2% |
| dpsc-9894 | 12.4%, 9.9% | 2.6%, 3.4% | −1.3%, −6.1% |
| dpsc-11418 | 2.4%, 1.6% | 31.6%, 29.1% | 23.7%, 17.5% |
| dpsc-11750 | 1.9%, 2.1% | 25.6%, 19.8% | 28.3%, 36.2% |
| rDT | ++, +++ | +, + | −−, −− |
| iDT | ++, + | +, + | −, − |
| sDT | +, + | ++, ++ | ++, ++ |
| Correlation (R2) | R2 = 0.868, R2 = 0.82 | R2 = 0.667, R2 = 0.588 | R2 = 0.904, R2 = 0.831 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, S.; Kang, B.; Lee, H.; Luu, H.; Mullins, E.; Kingsley, K. Characterization of Dental Pulp Stem Cell Responses to Functional Biomaterials Including Mineralized Trioxide Aggregates. J. Funct. Biomater. 2021, 12, 15. https://doi.org/10.3390/jfb12010015
Bae S, Kang B, Lee H, Luu H, Mullins E, Kingsley K. Characterization of Dental Pulp Stem Cell Responses to Functional Biomaterials Including Mineralized Trioxide Aggregates. Journal of Functional Biomaterials. 2021; 12(1):15. https://doi.org/10.3390/jfb12010015
Chicago/Turabian StyleBae, Sejin, Bueonguk Kang, Hyungbin Lee, Harrison Luu, Eric Mullins, and Karl Kingsley. 2021. "Characterization of Dental Pulp Stem Cell Responses to Functional Biomaterials Including Mineralized Trioxide Aggregates" Journal of Functional Biomaterials 12, no. 1: 15. https://doi.org/10.3390/jfb12010015