Abnormal Microvascular Architecture, Fibrosis, and Pericyte Characteristics in the Calf Muscle of Peripheral Artery Disease Patients with Claudication and Critical Limb Ischemia

 , ,
, ,
Abstract
:1. Introduction
2. Experimental Section
2.1. Human Subjects
2.1.1. Control Group
2.1.2. PAD Groups
2.2. Biopsy
2.3. Multi-Spectral Brightfield Microscopy: Quantification of Inter-Myofiber Collagen
2.3.1. Specimen Staining
2.3.2. Image Acquisition
2.4. Quantitative Fluorescence Microscopy
2.4.1. General
2.4.2. Quantification of Micro-Vessel Associated Collagen I (Principal Collagen of the Extracellular Matrix) and Collagen IV (Marker of the Basement Membrane)
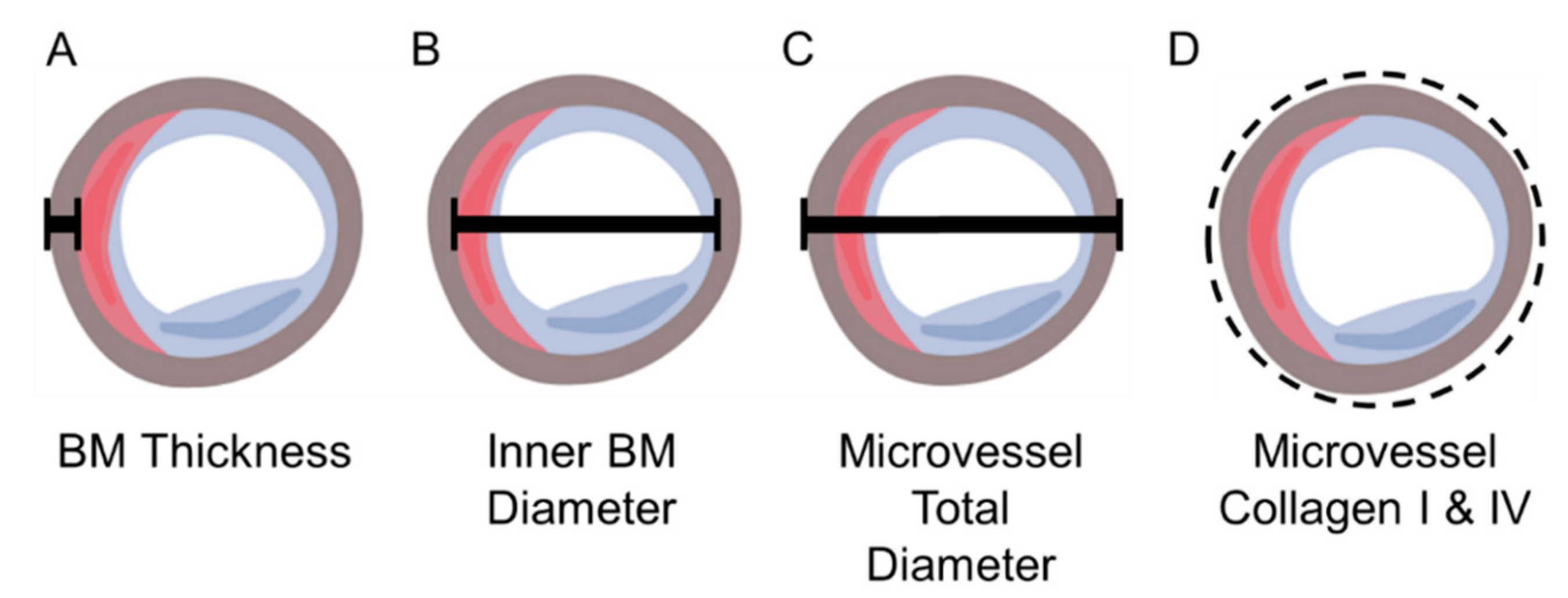
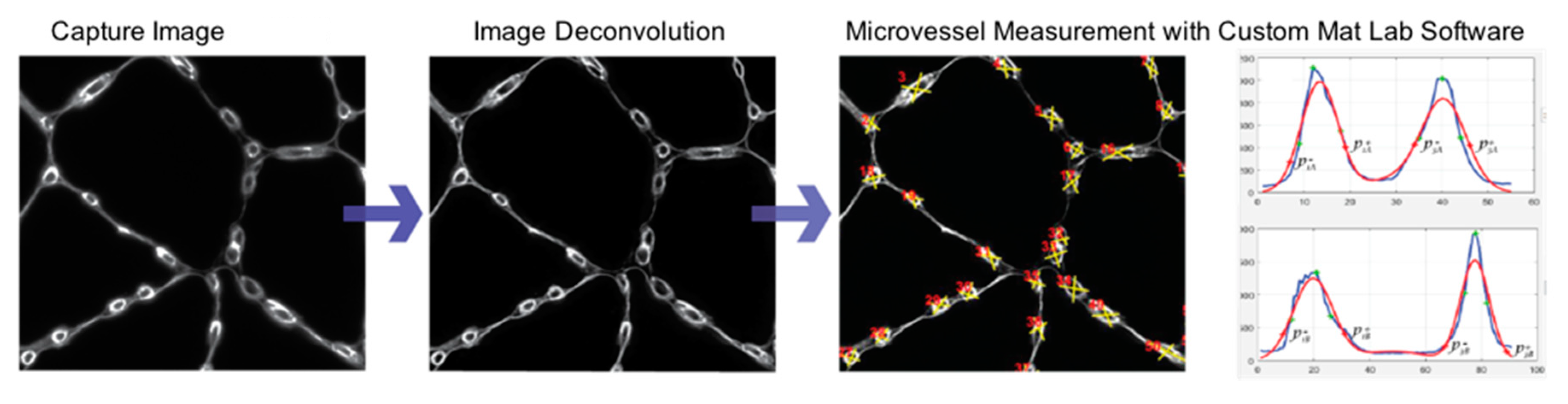
2.4.3. Measurement of Micro-Vessel Basement Membrane Thickness, Inner Diameter, and Overall Diameter
2.4.4. Identification of Micro-Vessel Associated Pericytes
2.5. Micro-Vessel Density
2.6. Statistical Analysis
3. Results
3.1. Patient Demographics
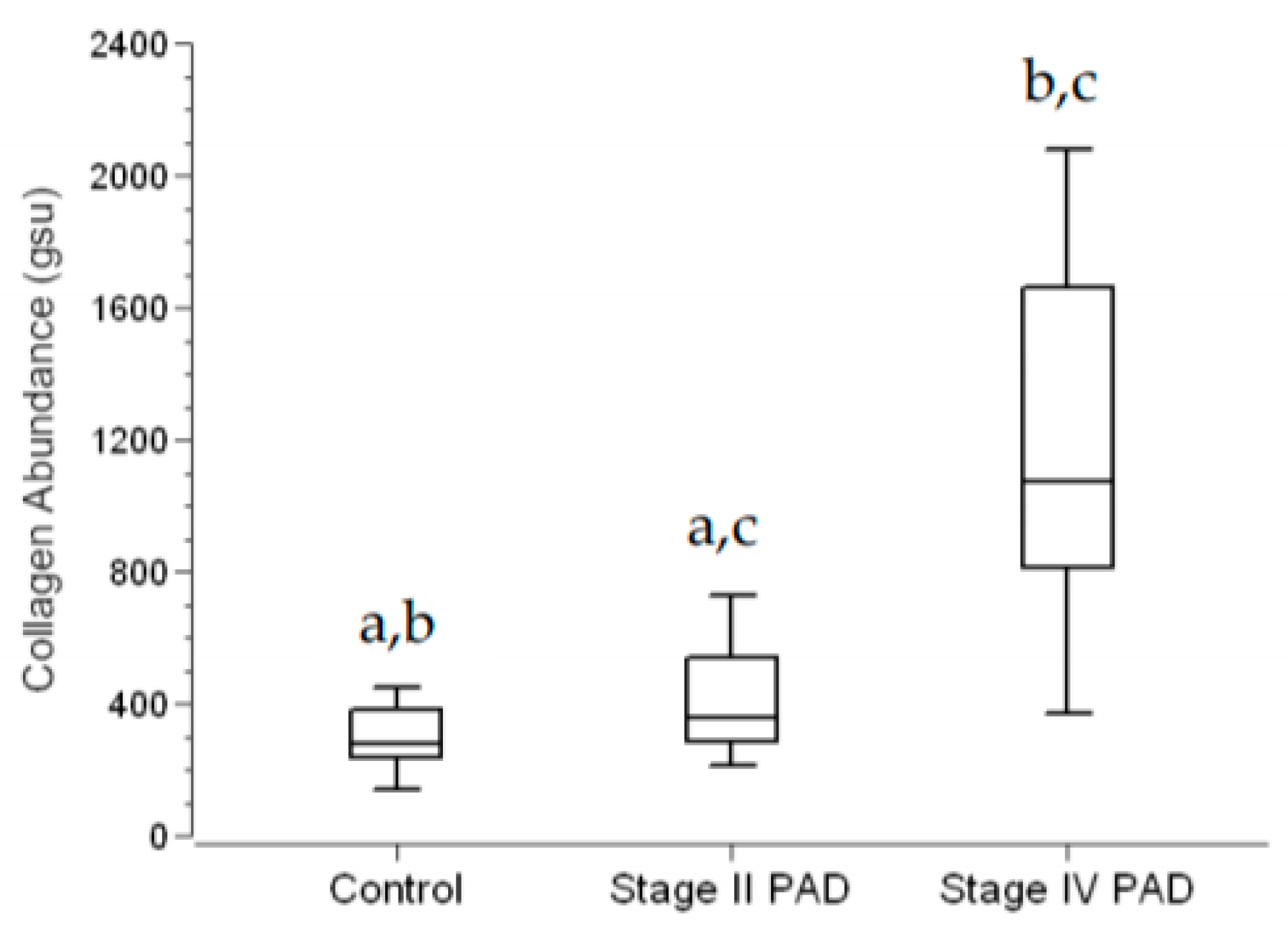
3.2. Fibrosis Is Increased in Calf Muscle of Patients with PAD
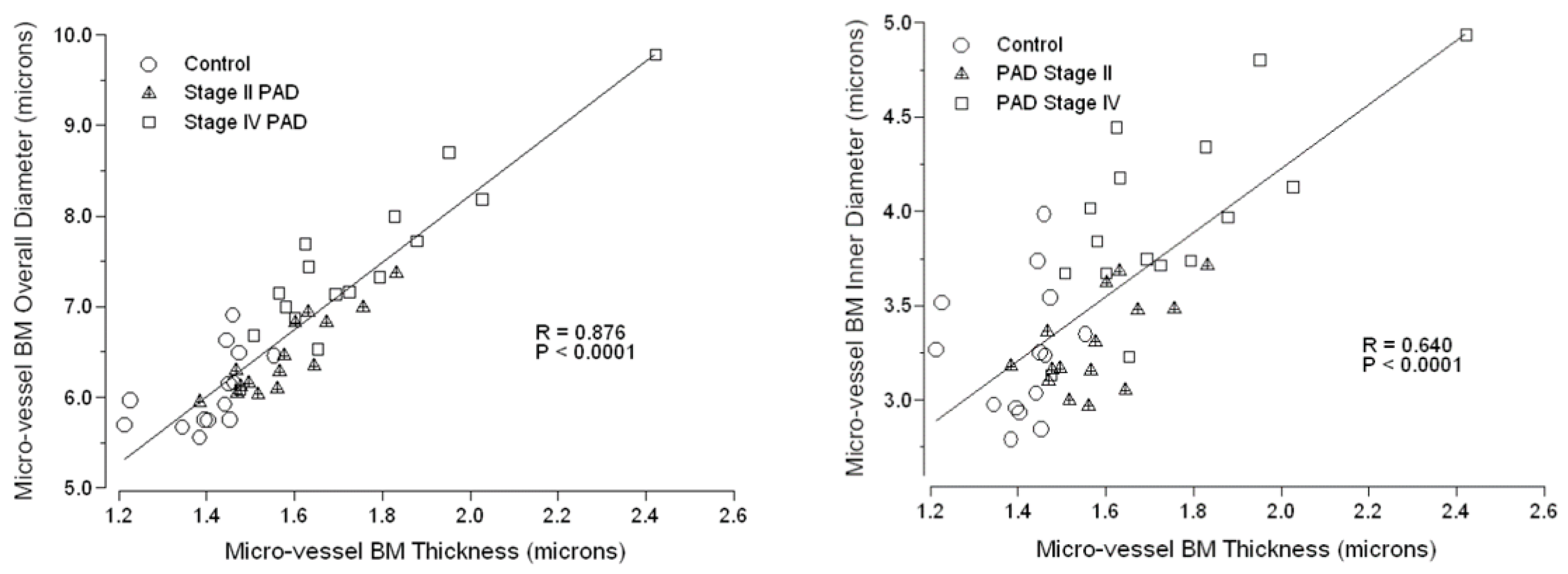
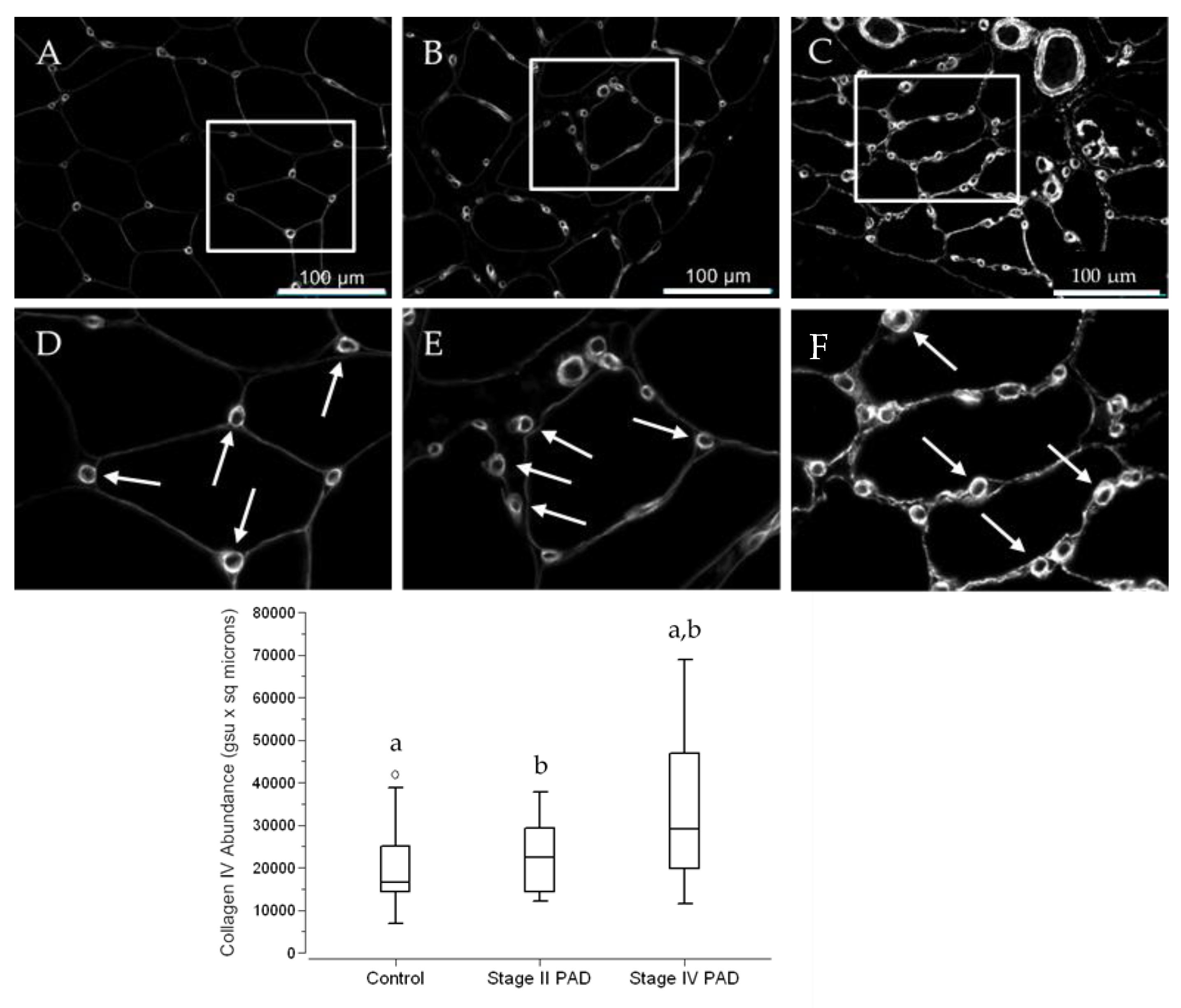
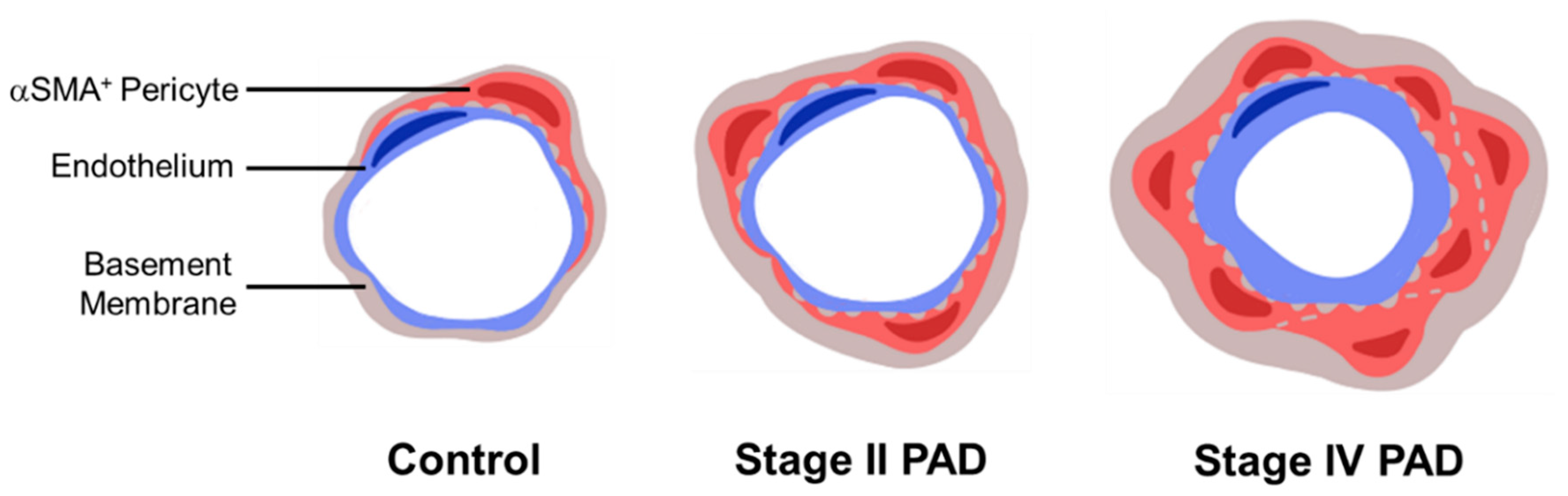
3.3. Micro-Vessel Basement Membrane Architecture Is Altered in Calf Muscle of Patients with PAD
3.4. The Abundance of Collagen IV in Calf Muscle Micro-Vessels across Control Subjects and PAD Patients Reflects the Architectural Measurements of Micro-Vessel Basement Membranes (BM)
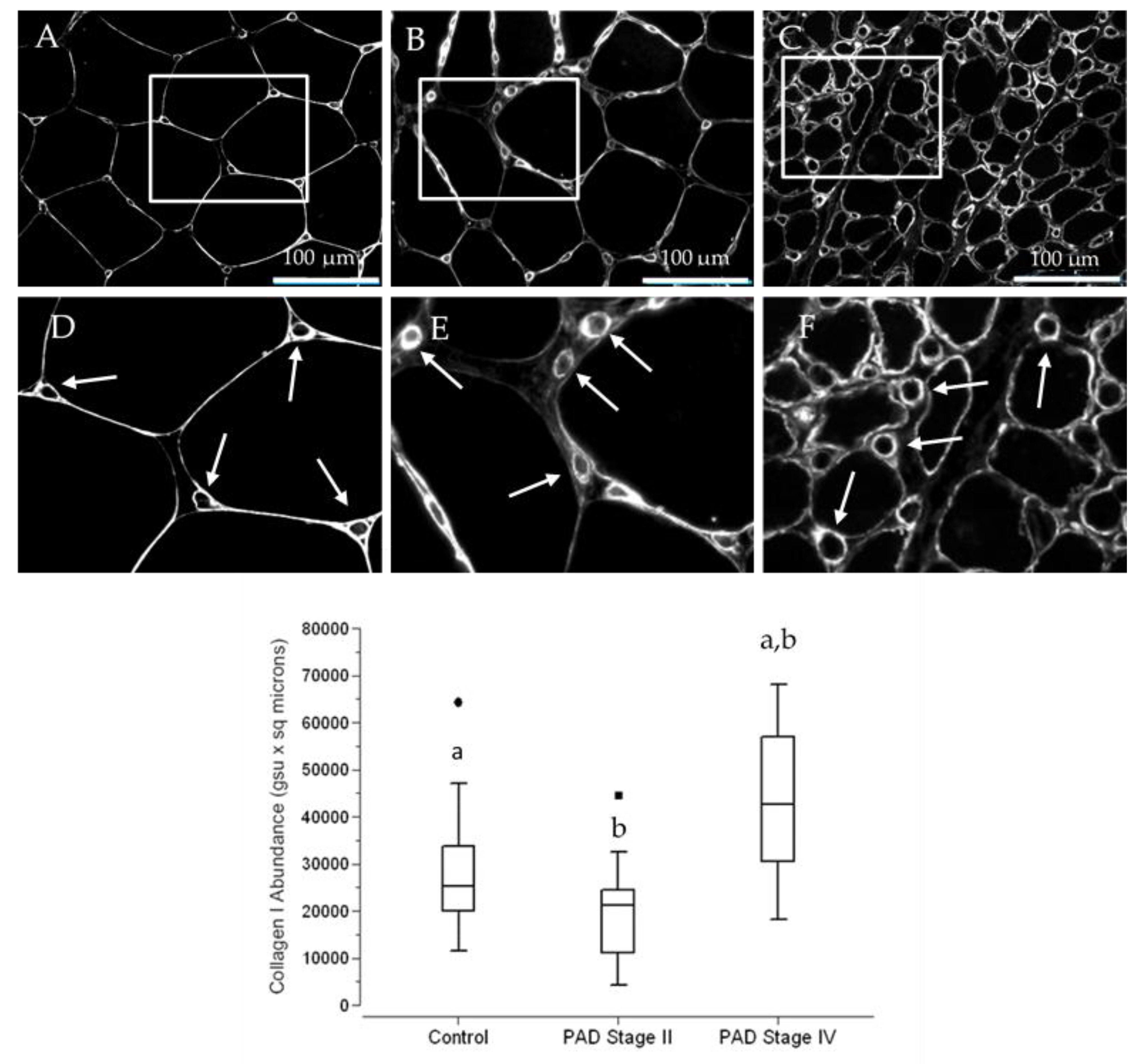
3.5. The Abundance of Collagen I, a Principal Contributor to Fibrosis, Is Increased in Terminal Micro-Vessels in Calf Muscle of PAD Patients with More Advanced Fibrosis
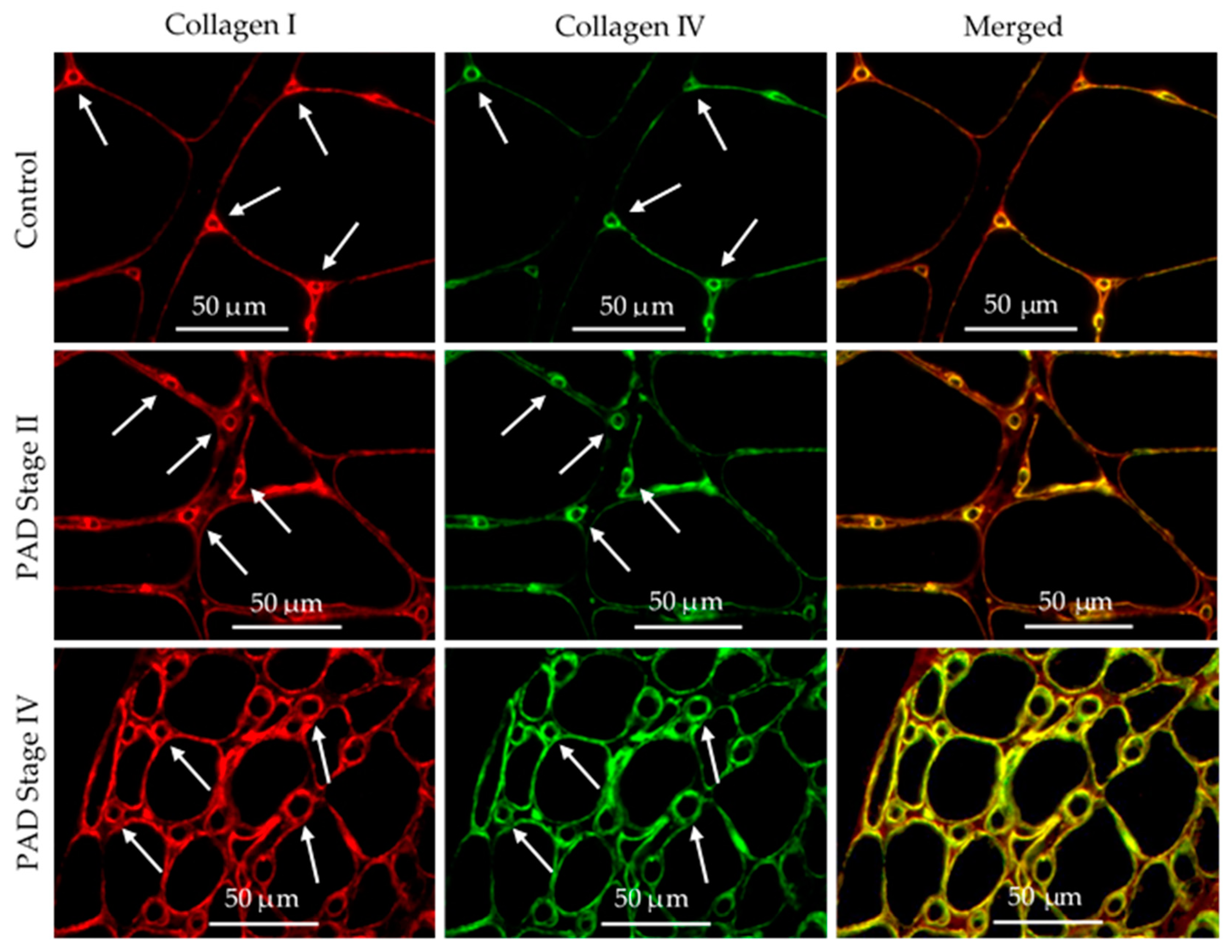
3.6. Collagen I and Collagen IV Were Co-Localized in the Basement Membrane of Calf Muscle Biopsies from Control Subjects and Patients with PAD
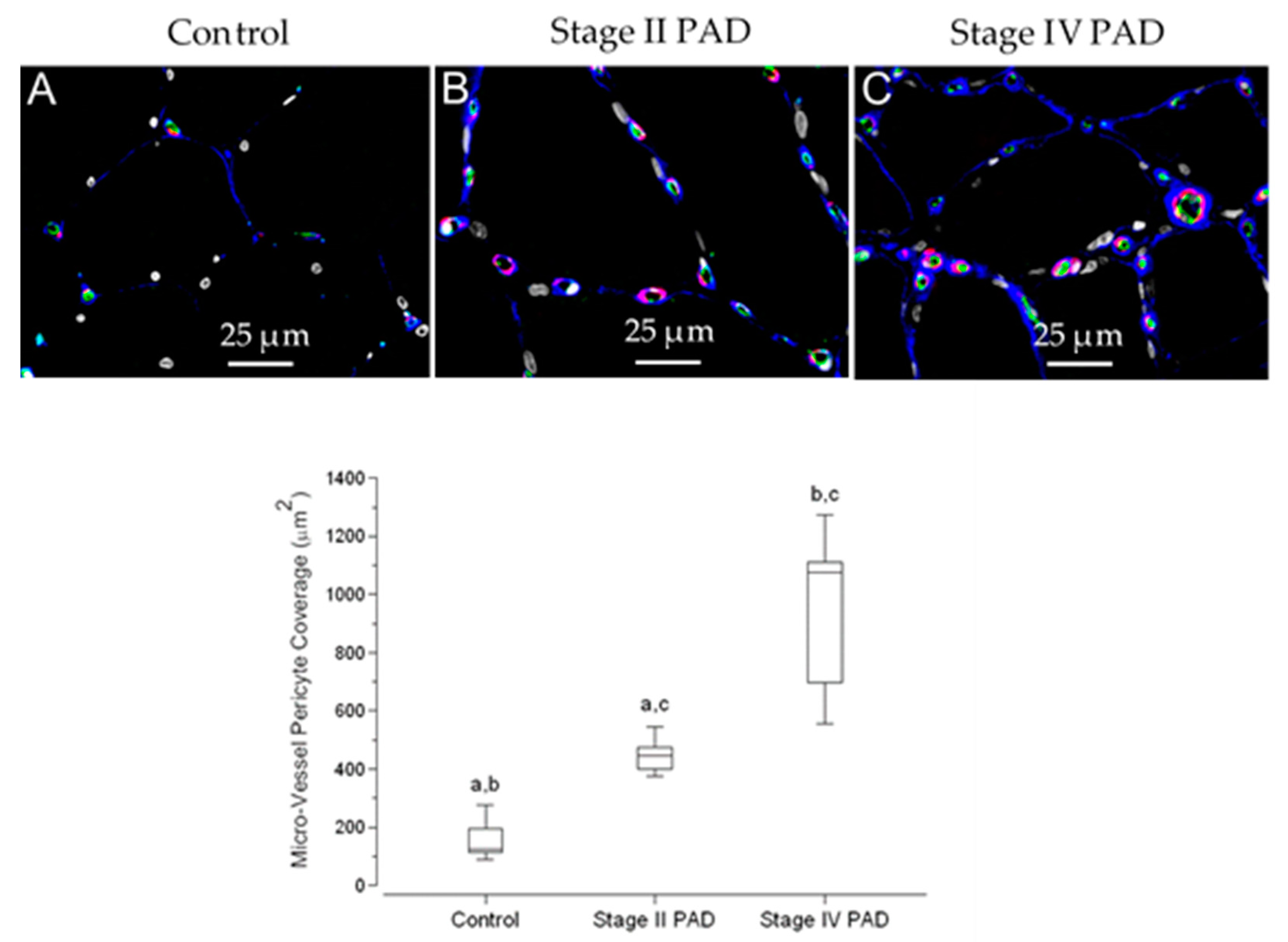
3.7. Pericyte Coverage of Micro-Vessels in Calf Muscle Specimens Was Greater in Patients with Stage IV Disease Compared to Controls and Patients with Stage II Disease
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rontoyanni, V.G.; Nunez Lopez, O.; Fankhauser, G.T.; Cheema, Z.F.; Rasmussen, B.B.; Porter, C. Mitochondrial Bioenergetics in the Metabolic Myopathy Accompanying Peripheral Artery Disease. Front. Physiol. 2017, 8, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brevetti, G.; Giugliano, G.; Brevetti, L.; Hiatt, W.R. Inflammation in peripheral artery disease. Circulation 2010, 122, 1862–1875. [Google Scholar] [CrossRef] [PubMed]
- McDermott, M.M. Lower extremity manifestations of peripheral artery disease: The pathophysiologic and functional implications of leg ischemia. Circ. Res. 2015, 116, 1540–1550. [Google Scholar] [CrossRef]
- Gardner, A.W.; Afaq, A. Management of lower extremity peripheral arterial disease. J. Cardiopulm. Rehabil. Prev. 2008, 28, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Hiatt, W.R.; Armstrong, E.J.; Larson, C.J.; Brass, E.P. Pathogenesis of the limb manifestations and exercise limitations in peripheral artery disease. Circ. Res. 2015, 116, 1527–1539. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, R.; Kim, M.; Kieny, R. Surgical treatment of peripheral circulation disorders. Helv. Chir. Acta 1954, 21, 499–533. [Google Scholar]
- Dormandy, J.A.; Rutherford, R.B. Management of peripheral arterial disease (PAD). TASC Working Group. TransAtlantic Inter-Society Consensus (TASC). J. Vasc. Surg. 2000, 31, S1–S296. [Google Scholar]
- Novo, S. Classification, epidemiology, risk factors, and natural history of peripheral arterial disease. Diabetes Obes. Metab. 2002, 4, S1–S6. [Google Scholar] [CrossRef]
- Pipinos, I.I.; Judge, A.R.; Selsby, J.T.; Zhu, Z.; Swanson, S.A.; Nella, A.A.; Dodd, S.L. The myopathy of peripheral arterial occlusive disease: Part 2. Oxidative stress, neuropathy, and shift in muscle fiber type. Vasc. Endovasc. Surg. 2008, 42, 101–112. [Google Scholar] [CrossRef]
- Regensteiner, J.G.; Wolfel, E.E.; Brass, E.P.; Carry, M.R.; Ringel, S.P.; Hargarten, M.E.; Stamm, E.R.; Hiatt, W.R. Chronic changes in skeletal muscle histology and function in peripheral arterial disease. Circulation 1993, 87, 413–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cluff, K.; Miserlis, D.; Naganathan, G.K.; Pipinos, I.I.; Koutakis, P.; Samal, A.; McComb, R.D.; Subbiah, J.; Casale, G.P. Morphometric analysis of gastrocnemius muscle biopsies from patients with peripheral arterial disease: Objective grading of muscle degeneration. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R291–R299. [Google Scholar] [CrossRef] [PubMed]
- Koutakis, P.; Weiss, D.J.; Miserlis, D.; Shostrom, V.K.; Papoutsi, E.; Ha, D.M.; Carpenter, L.A.; McComb, R.D.; Casale, G.P.; Pipinos, I.I. Oxidative damage in the gastrocnemius of patients with peripheral artery disease is myofiber type selective. Redox Biol. 2014, 2, 921–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koutakis, P.; Myers, S.A.; Cluff, K.; Ha, D.M.; Haynatzki, G.; McComb, R.D.; Uchida, K.; Miserlis, D.; Papoutsi, E.; Johanning, J.M.; et al. Abnormal myofiber morphology and limb dysfunction in claudication. J. Surg. Res. 2015, 196, 172–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, D.J.; Casale, G.P.; Koutakis, P.; Nella, A.A.; Swanson, S.A.; Zhu, Z.; Miserlis, D.; Johanning, J.M.; Pipinos, I.I. Oxidative damage and myofiber degeneration in the gastrocnemius of patients with peripheral arterial disease. J. Transl. Med. 2013, 11, 230. [Google Scholar] [CrossRef] [Green Version]
- Ha, D.M.; Carpenter, L.C.; Koutakis, P.; Swanson, S.A.; Zhu, Z.; Hanna, M.; DeSpiegelaere, H.K.; Pipinos, I.I.; Casale, G.P. Transforming growth factor-beta 1 produced by vascular smooth muscle cells predicts fibrosis in the gastrocnemius of patients with peripheral artery disease. J. Transl. Med. 2016, 14, 39. [Google Scholar] [CrossRef] [Green Version]
- Birbrair, A.; Zhang, T.; Files, D.C.; Mannava, S.; Smith, T.; Wang, Z.M.; Messi, M.L.; Mintz, A.; Delbono, O. Type-1 pericytes accumulate after tissue injury and produce collagen in an organ-dependent manner. Stem Cell Res. 2014, 5, 122. [Google Scholar] [CrossRef] [Green Version]
- Gai, Z.; Chu, L.; Xu, Z.; Song, X.; Sun, D.; Kullak-Ublick, G.A. Farnesoid X receptor activation protects the kidney from ischemia-reperfusion damage. Sci. Rep. 2017, 7, 9815. [Google Scholar] [CrossRef]
- Stratman, A.N.; Davis, G.E. Endothelial cell-pericyte interactions stimulate basement membrane matrix assembly: Influence on vascular tube remodeling, maturation, and stabilization. Microsc. Microanal. 2012, 18, 68–80. [Google Scholar] [CrossRef] [Green Version]
- Alarcon-Martinez, L.; Yilmaz-Ozcan, S.; Yemisci, M.; Schallek, J.; Kilic, K.; Can, A.; Di Polo, A.; Dalkara, T. Capillary pericytes express alpha-smooth muscle actin, which requires prevention of filamentous-actin depolymerization for detection. eLife 2018, 7, e34861. [Google Scholar] [CrossRef]
- Norgren, L.; Hiatt, W.R.; Dormandy, J.A.; Nehler, M.R.; Harris, K.A.; Fowkes, F.G.; Group, T.I.W. Inter-Society Consensus for the Management of Peripheral Arterial Disease (TASC II). J. Vasc. Surg. 2007, 45, S5–S67. [Google Scholar] [CrossRef] [Green Version]
- Baum, O.; Djonov, V.; Ganster, M.; Widmer, M.; Baumgartner, I. Arteriolization of capillaries and FGF-2 upregulation in skeletal muscles of patients with chronic peripheral arterial disease. Microcirculation 2005, 12, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Dulauroy, S.; Di Carlo, S.E.; Langa, F.; Eberl, G.; Peduto, L. Lineage tracing and genetic ablation of ADAM12(+) perivascular cells identify a major source of profibrotic cells during acute tissue injury. Nat. Med. 2012, 18, 1262–1270. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, S.N.; Iredale, J.P.; Henderson, N.C. Origins of fibrosis: Pericytes take centre stage. F1000Prime Rep. 2013, 5, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birbrair, A.; Zhang, T.; Wang, Z.M.; Messi, M.L.; Mintz, A.; Delbono, O. Type-1 pericytes participate in fibrous tissue deposition in aged skeletal muscle. Am. J. Physiol. Cell Physiol. 2013, 305, C1098–C1113. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.F.; Chiang, W.C.; Lai, C.F.; Chang, F.C.; Chen, Y.T.; Chou, Y.H.; Wu, T.H.; Linn, G.R.; Ling, H.; Wu, K.D.; et al. Transforming growth factor beta-1 stimulates profibrotic epithelial signaling to activate pericyte-myofibroblast transition in obstructive kidney fibrosis. Am. J. Pathol. 2013, 182, 118–131. [Google Scholar] [CrossRef] [Green Version]
- Pipinos, I.I.; Judge, A.R.; Selsby, J.T.; Zhu, Z.; Swanson, S.A.; Nella, A.A.; Dodd, S.L. The myopathy of peripheral arterial occlusive disease: Part 1. Functional and histomorphological changes and evidence for mitochondrial dysfunction. Vasc. Endovasc. Surg. 2007, 41, 481–489. [Google Scholar] [CrossRef]
- Ismail, S.; Sturrock, A.; Wu, P.; Cahill, B.; Norman, K.; Huecksteadt, T.; Sanders, K.; Kennedy, T.; Hoidal, J. NOX4 mediates hypoxia-induced proliferation of human pulmonary artery smooth muscle cells: The role of autocrine production of transforming growth factor-{beta}1 and insulin-like growth factor binding protein-3. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L489–L499. [Google Scholar] [CrossRef]
- Sturrock, A.; Cahill, B.; Norman, K.; Huecksteadt, T.P.; Hill, K.; Sanders, K.; Karwande, S.V.; Stringham, J.C.; Bull, D.A.; Gleich, M.; et al. Transforming growth factor-beta1 induces Nox4 NAD(P)H oxidase and reactive oxygen species-dependent proliferation in human pulmonary artery smooth muscle cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 290, L661–L673. [Google Scholar] [CrossRef]
- Ricard, N.; Tu, L.; Le Hiress, M.; Huertas, A.; Phan, C.; Thuillet, R.; Sattler, C.; Fadel, E.; Seferian, A.; Montani, D.; et al. Increased pericyte coverage mediated by endothelial-derived fibroblast growth factor-2 and interleukin-6 is a source of smooth muscle-like cells in pulmonary hypertension. Circulation 2014, 129, 1586–1597. [Google Scholar] [CrossRef] [Green Version]
- Hayden, M.R.; Sowers, J.R.; Tyagi, S.C. The central role of vascular extracellular matrix and basement membrane remodeling in metabolic syndrome and type 2 diabetes: The matrix preloaded. Cardiovasc. Diabetol. 2005, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Wei, K.; Jayaweera, A.R.; Firoozan, S.; Linka, A.; Skyba, D.M.; Kaul, S. Quantification of myocardial blood flow with ultrasound-induced destruction of microbubbles administered as a constant venous infusion. Circulation 1998, 97, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Amarteifio, E.; Weber, M.A.; Wormsbecher, S.; Demirel, S.; Krakowski-Roosen, H.; Jores, A.; Braun, S.; Delorme, S.; Bockler, D.; Kauczor, H.U.; et al. Dynamic contrast-enhanced ultrasound for assessment of skeletal muscle microcirculation in peripheral arterial disease. Investig. Radiol. 2011, 46, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Duerschmied, D.; Olson, L.; Olschewski, M.; Rossknecht, A.; Freund, G.; Bode, C.; Hehrlein, C. Contrast ultrasound perfusion imaging of lower extremities in peripheral arterial disease: A novel diagnostic method. Eur. Heart J. 2006, 27, 310–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundi, R.; Prior, S.J.; Addison, O.; Lu, M.; Ryan, A.S.; Lal, B.K. Contrast-Enhanced Ultrasound Reveals Exercise-Induced Perfusion Deficits in Claudicants. J. Vasc. Endovasc. Surg. 2017, 2, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, B.P.; Belcik, J.T.; Landry, G.; Linden, J.; Lindner, J.R. Exercise versus vasodilator stress limb perfusion imaging for the assessment of peripheral artery disease. Echocardiography 2017, 34, 1187–1194. [Google Scholar] [CrossRef]
- Tilton, R.G.; Faller, A.M.; Burkhardt, J.K.; Hoffmann, P.L.; Kilo, C.; Williamson, J.R. Pericyte degeneration and acellular capillaries are increased in the feet of human diabetic patients. Diabetologia 1985, 28, 895–900. [Google Scholar] [CrossRef] [Green Version]
- Brunner, G.; Bismuth, J.; Nambi, V.; Ballantyne, C.M.; Taylor, A.A.; Lumsden, A.B.; Morrisett, J.D.; Shah, D.J. Calf muscle perfusion as measured with magnetic resonance imaging to assess peripheral arterial disease. Med. Biol. Eng. Comput. 2016, 54, 1667–1681. [Google Scholar] [CrossRef] [Green Version]
- Mathew, R.C.; Kramer, C.M. Recent advances in magnetic resonance imaging for peripheral artery disease. Vasc. Med. 2018, 23, 143–152. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Stage II PAD | Stage IV PAD | |
|---|---|---|---|
| Number of Subjects | 14 | 15 | 16 |
| Mean Age (years) | 67.3 ± 1.3 | 64.3 ± 1.2 | 69.4 ± 1.0 |
| Ankle Brachial Index | 1.09 ± 0.03 | 0.60 ± 0.03 ** | 0.18 ± 0.06 **,ǂ |
| Gender (male/female) | 14/0 | 15/0 | 15/1 |
| Smoking (never/current/former) | 6/6/2 | 0/6/9 | 2/9/5 |
| Coronary artery disease | 4/14 | 3/15 | 8/16 |
| Cerebrovascular disease (none/TIA A/stroke) | 13/1/0 | 13/0/2 | 12/1/3 |
| Obesity B | 6 | 3 | 4 |
| Dyslipidemia | 10 | 8 | 7 |
| Diabetes Mellitus | 2 | 2 | 8 *,† |
| Chronic Obstructive Pulmonary Disease | 0 | 3 | 1 |
| Renal insufficiency C | 1 | 0 | 7 *,ǂ |
| Statins | 9 | 12 | 9 |
| Family history of cardiovascular disease | 5 | 7 | 14 *,† |
| Hypertension | 8 | 9 | 14 |
| Micro-Vessel BM Measurements (μm) | Control | Stage II PAD | Stage IV PAD |
|---|---|---|---|
| Thickness | 1.408 ± 0.025 | 1.577 ± 0.030 * | 1.747 ± 0.060 *,† |
| Inner Diameter | 3.243 ± 0.094 | 3.295 ± 0.064 | 3.972 ± 0.124 *,ǂ |
| Overall Diameter | 6.058 ± 0.111 | 6.448 ± 0.112 | 7.466 ± 0.224 *,ǂ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mietus, C.J.; Lackner, T.J.; Karvelis, P.S.; Willcockson, G.T.; Shields, C.M.; Lambert, N.G.; Koutakis, P.; Fuglestad, M.A.; Hernandez, H.; Haynatzki, G.R.; et al. Abnormal Microvascular Architecture, Fibrosis, and Pericyte Characteristics in the Calf Muscle of Peripheral Artery Disease Patients with Claudication and Critical Limb Ischemia. J. Clin. Med. 2020, 9, 2575. https://doi.org/10.3390/jcm9082575
Mietus CJ, Lackner TJ, Karvelis PS, Willcockson GT, Shields CM, Lambert NG, Koutakis P, Fuglestad MA, Hernandez H, Haynatzki GR, et al. Abnormal Microvascular Architecture, Fibrosis, and Pericyte Characteristics in the Calf Muscle of Peripheral Artery Disease Patients with Claudication and Critical Limb Ischemia. Journal of Clinical Medicine. 2020; 9(8):2575. https://doi.org/10.3390/jcm9082575
Chicago/Turabian StyleMietus, Constance J., Timothy J. Lackner, Petros S. Karvelis, Gregory T. Willcockson, Christina M. Shields, Nicholas G. Lambert, Panagiotis Koutakis, Matthew A. Fuglestad, Hernan Hernandez, Gleb R. Haynatzki, and et al. 2020. "Abnormal Microvascular Architecture, Fibrosis, and Pericyte Characteristics in the Calf Muscle of Peripheral Artery Disease Patients with Claudication and Critical Limb Ischemia" Journal of Clinical Medicine 9, no. 8: 2575. https://doi.org/10.3390/jcm9082575