Influence of Bone Marrow-Derived Mesenchymal Stem Cell Therapy on Oxidative Stress Intensity in Minimally Conscious State Patients

 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Patient Characteristic
2.2. Bone Marrow Collection and Isolation of MSC
2.3. Plasma and Cerebrospinal Fluid Collection
2.4. Antioxidant Capacity Measurement
2.5. Statistical Analysis
3. Results
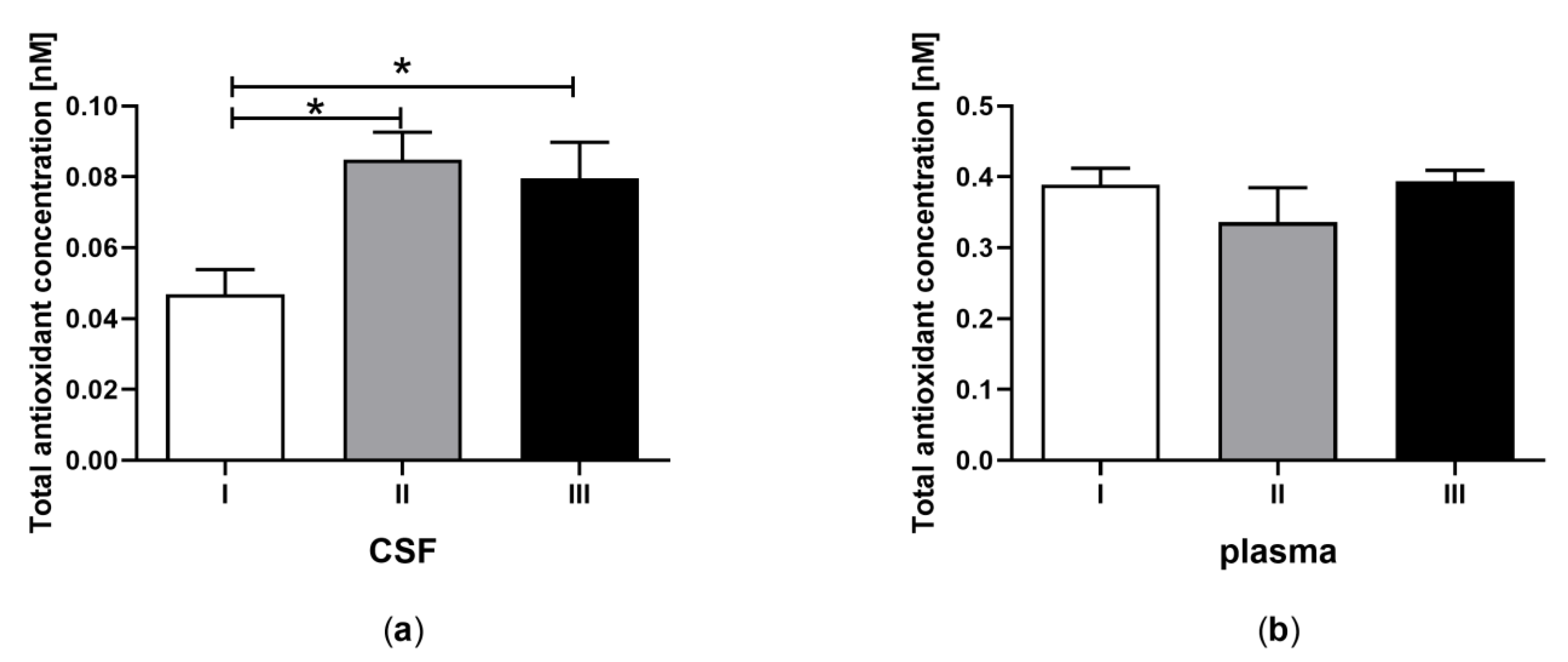
3.1. Total Antioxidant Capacity in CSF and Plasma of the Patients in MCS Undergoing BM-MSC Treatment
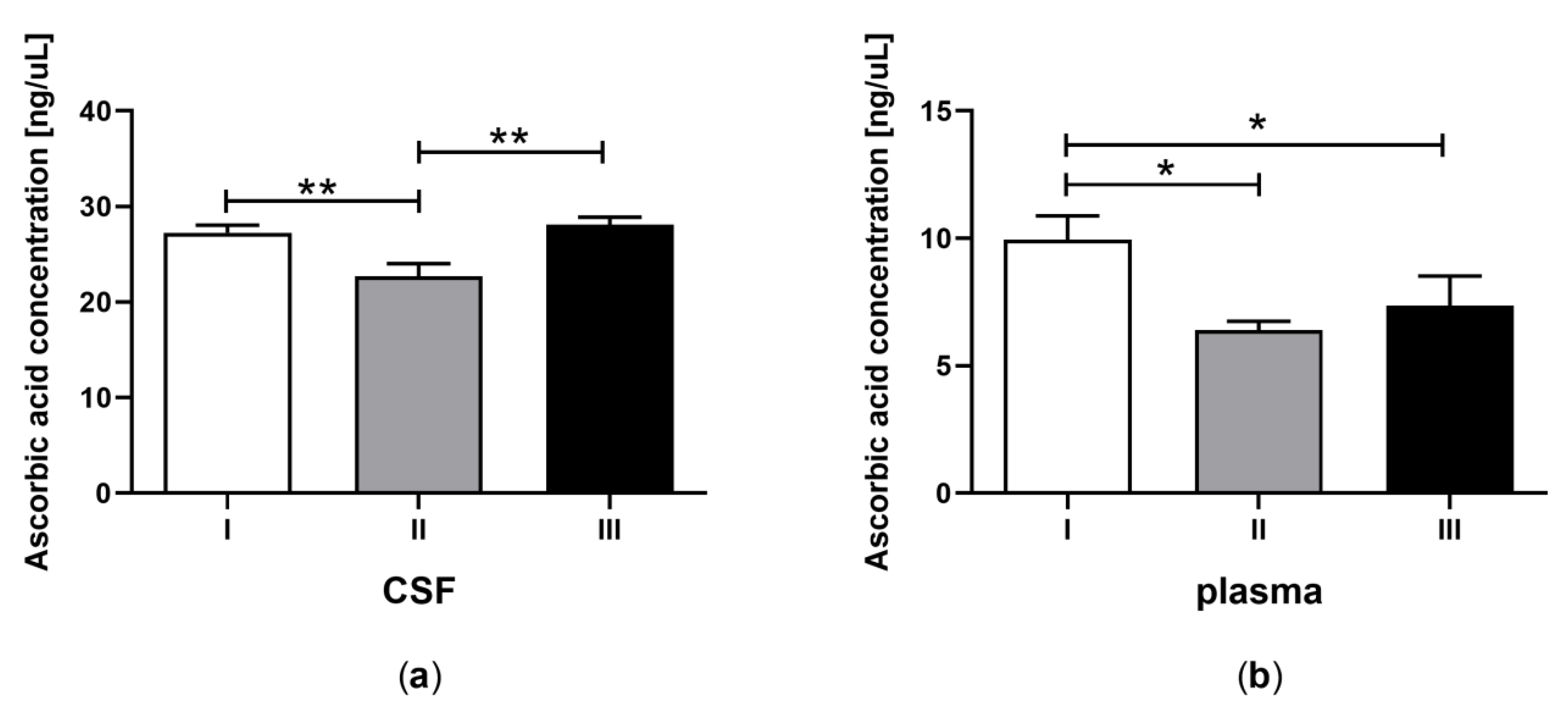
3.2. Ascorbic Acid Concentration in CSF and Plasma of the Patients in MCS Undergoing BM-MSC Treatment
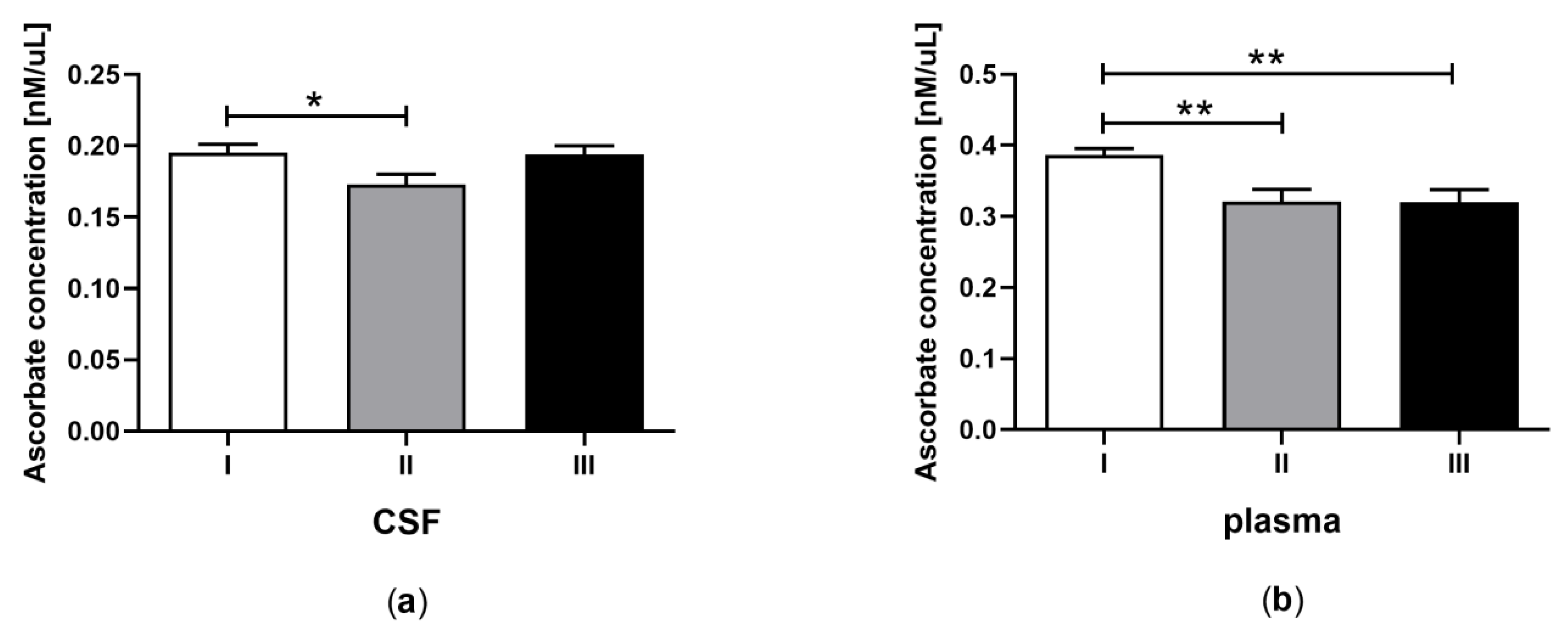
3.3. Total Ascorbate Concentration in CSF and Plasma of the Patients in MCS Undergoing BM-MSC Treatment
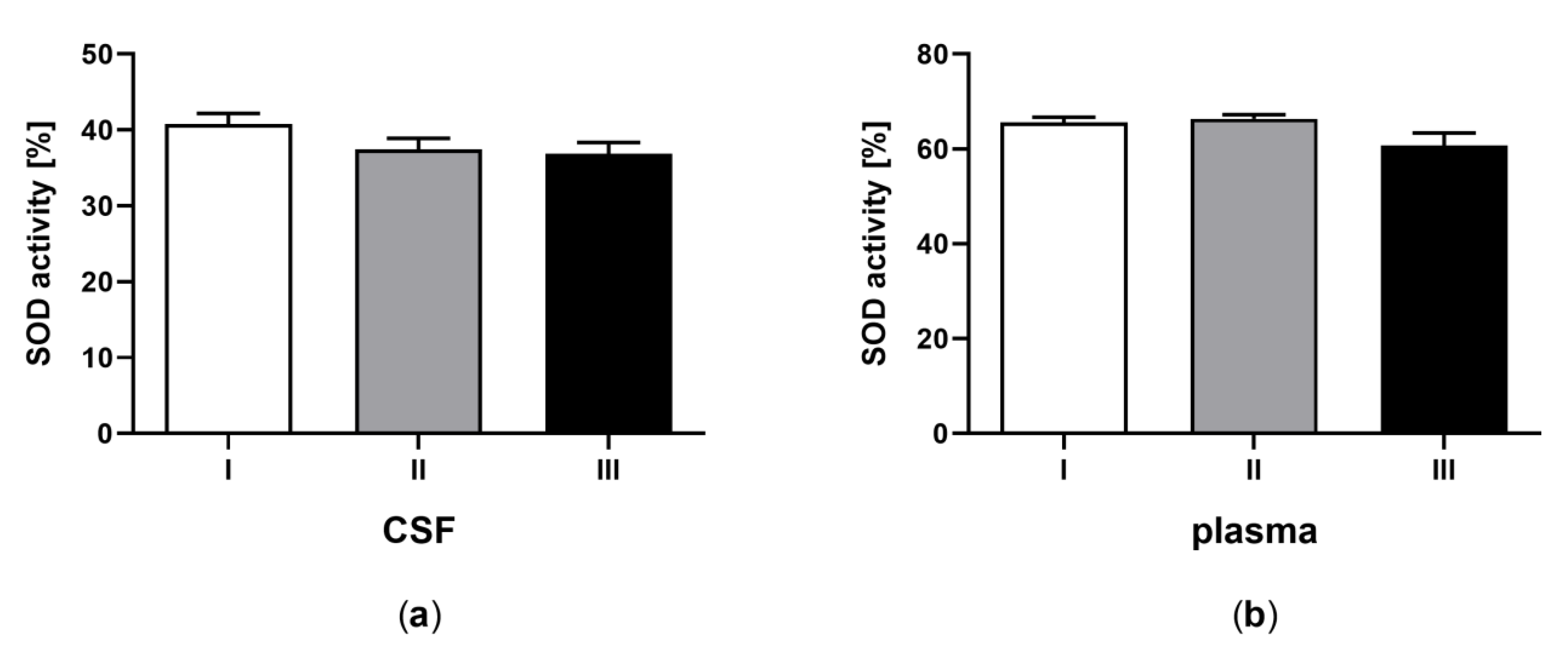
3.4. Superoxide Dismutase Activity in CSF and Plasma of the Patients in MCS Undergoing BM-MSC Treatment
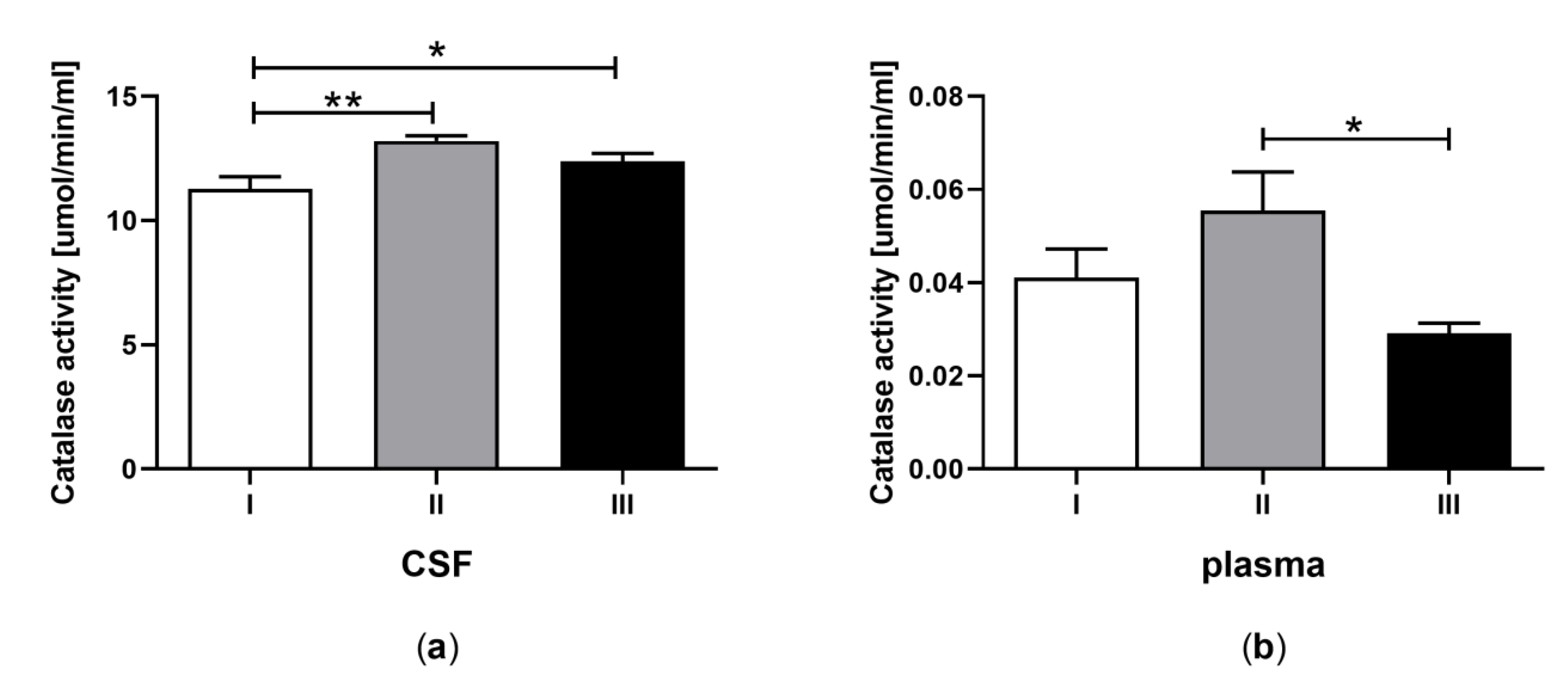
3.5. Catalase Activity in CSF and Plasma of the Patients in MCS Undergoing BM-MSC Treatment
3.6. Peroxidase Activity in CSF and Plasma of the Patients in MCS Undergoing BM-MSC Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wijdicks, E.F.M. The bare essentials: Coma. Pract. Neurol. 2010, 10, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Traub, S.J.; Wijdicks, E.F. Initial Diagnosis and Management of Coma. Emerg. Med. Clin. North Am. 2016, 34, 777–793. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, L.; McWhirter, L.; Williams, S.; Derry, C.; Stone, J. Functional coma. Handb. Clin. Neurol. 2016, 139, 313–327. [Google Scholar] [PubMed]
- Giacino, J.T.; Ashwal, S.; Childs, N.; Cranford, R.; Jennett, B.; Katz, D.I.; Kelly, J.P.; Rosenberg, J.H.; Whyte, J.; Zafonte, R.D.; et al. The minimally conscious state: Definition and diagnostic criteria. Neurology 2002, 58, 349–353. [Google Scholar] [CrossRef]
- Bhat, A.H.; Dar, K.B.; Anees, S.; Zargar, M.A.; Masood, A.; Sofi, M.A.; Ganie, S.A. Oxidative stress, mitochondrial dysfunction and neurodegenerative diseases; a mechanistic insight. Biomed. Pharmacother. 2015, 74, 101–110. [Google Scholar] [CrossRef]
- Islam, M.T. Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol. Res. 2016, 39, 73–82. [Google Scholar] [CrossRef]
- Salim, S. Oxidative Stress and the Central Nervous System. J. Pharmacol. Exp. Ther. 2017, 360, 201–205. [Google Scholar] [CrossRef]
- Angelova, P.R.; Abramov, A.Y. Role of mitochondrial ROS in the brain: From physiology to neurodegeneration. FEBS Lett. 2018, 592, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Nissanka, N.; Moraes, C.T. Mitochondrial DNA damage and reactive oxygen species in neurodegenerative disease. FEBS Lett. 2017, 592, 728–742. [Google Scholar] [CrossRef]
- Zhuo, M.; Small, S.A.; Kandel, E.R.; Hawkins, R.D. Nitric oxide and carbon monoxide produce activity-dependent long-term synaptic enhancement in hippocampus. Science 1993, 260, 1946–1950. [Google Scholar] [CrossRef]
- Massaad, C.A.; Klann, E. Reactive Oxygen Species in the Regulation of Synaptic Plasticity and Memory. Antioxid. Redox. Signal. 2011, 14, 2013–2054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Dell, T.J.; Hawkins, R.D.; Kandel, E.R.; Arancio, O. Tests of the roles of two diffusible substances in long-term potentiation: Evidence for nitric oxide as a possible early retrograde messenger. Proc. Natl. Acad. Sci. USA 1991, 88, 11285–11289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, A.; Hirsch, D.J.; Glatt, C.E.; Ronnett, G.V.; Snyder, S.H. Carbon monoxide: A putative neural messenger. Science 1993, 259, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Knapp, L.T.; Klann, E. Role of reactive oxygen species in hippocampal long-term potentiation: Contributory or inhibitory? J. Neurosci. Res. 2002, 70, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kohen, R.; Beit-Yannai, E.; Berry, E.M.; Tirosh, O. Overall low molecular weight antioxidant activity of biological fluids and tissues by cyclic voltammetry. Meth. Enzymol. 1999, 300, 285–296. [Google Scholar] [PubMed]
- Kohen, R.; Vellaichamy, E.; Hrbac, J.; Gati, I.; Tirosh, O. Quantification of the overall reactive oxygen species scavenging capacity of biological fluids and tissues. Free Radic. Biol. Med. 2000, 28, 871–879. [Google Scholar] [CrossRef]
- Nualart, F.; Mack, L.; García, A.; Cisternas, P.; Bongarzone, E.R.; Heitzer, M.; Jara, N.; Martínez, F.; Ferrada, L.; Espinoza, F.; et al. Vitamin C Transporters, Recycling and the Bystander Effect in the Nervous System: SVCT2 versus Gluts. J. Stem Cell Res. Ther. 2014, 4, 209. [Google Scholar] [CrossRef] [Green Version]
- Chance, B.; Schoener, B.; Oshino, R.; Itshak, F.; Nakase, Y. Oxidation-reduction ratio studies of mitochondria in freeze-trapped samples. NADH and flavoprotein fluorescence signals. J. Biol. Chem. 1979, 254, 4764–4771. [Google Scholar]
- Griendling, K.K.; Sorescu, D.; Lassègue, B.; Ushio-Fukai, M. Modulation of protein kinase activity and gene expression by reactive oxygen species and their role in vascular physiology and pathophysiology. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 2175–2183. [Google Scholar] [CrossRef] [Green Version]
- Saso, L.; Firuzi, O. Pharmacological applications of antioxidants: Lights and shadows. Curr. Drug Targets 2014, 15, 1177–1199. [Google Scholar] [CrossRef]
- Strauss, D.J.; Ashal, S.; Day, S.M.; Schavelle, M.R. Life expectancy of children in vegetative and minimally conscious states. Pediatr. Neurol. 2000, 23, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Laureys, S.; Owen, A.M.; Schiff, N.D. Behavioural improvements with thalamic stimulation after severe traumatic brain injury. Nature 2007, 448, 600–603. [Google Scholar]
- Angelakis, E.; Liouta, E.; Andreadis, N.; Korfias, S.; Ktonas, P.; Stranjalis, G.; Sakas, D.E. Transcranial direct current stimulation (tDCS) effects in disorders of consciousness. Arch. Phys. Med. Rehabil. 2014, 95, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Thibaut, A.; Bruno, M.A.; Ledoux, D.; Demertzi, A.; Laureys, S. tDCS in patients with disorders of consciousness: Sham-controlled randomized double-blind study. Neurology 2014, 82, 1112–1118. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Ringden, O. Immunobiology of human mesenchymal stem cells and future use in hematopoietic stem cell transplantation. Am. Soc. Blood Marrow Trans. 2005, 11, 321–334. [Google Scholar] [CrossRef] [Green Version]
- Yokokawa, K.; Iwahara, N.; Hisahara, S.; Emoto, M.C.; Saito, T.; Suzuki, H.; Manabe, T.; Matsumura, A.; Matsushita, T.; Suzuki, S.; et al. Transplantation of Mesenchymal Stem Cells Improves Amyloid-β Pathology by Modifying Microglial Function and Suppressing Oxidative Stress. J. Alzheimers Dis. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurozumi, K.; Nakamura, K.; Tamiya, T.; Kawano, Y.; Ishii, K.; Kobune, M.; Hirai, S.; Uchida, H.; Sasaki, K.; Ito, Y.; et al. Mesenchymal stem cells that produce neurotrophic factors reduce ischemic damage in the rat middle cerebral artery occlusion model. Mol. Ther. 2005, 11, 96–104. [Google Scholar] [CrossRef]
- Sharma, S.; Yang, B.; Strong, R.; Xi, X.; Brenneman, M.; Grotta, J.C.; Aronowski, J.; Savitz, S.I. Bone marrow mononuclear cells protect neurons and modulate microglia in cell culture models of ischemic stroke. J. Neurosci. Res. 2010, 88, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Savitz, S.I.; Misra, V.; Kasam, M.; Juneja, H.; Cox, C.S.; Alderman, S., Jr. Intravenous autologous bone marrow mononuclear cells for ischemic stroke. Ann. Neurol. 2011, 70, 59–69. [Google Scholar] [CrossRef]
- Galgano, M.; Toshkezi, G.; Qiu, X.; Russell, T.; Chin, L.; Zhao, L.R. Traumatic Brain Injury: Current Treatment Strategies and Future Endeavors. Cell Transplant. 2017, 26, 1118–1130. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, A.; Kemp, K.; Ginty, M.; Hares, K.; Mallam, E.; Scolding, N. Human bone marrow-derived mesenchymal stem cells secrete brain-derived neurotrophic factor which promotes neuronal survival in vitro. Stem Cell Res. 2009, 3, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, X.; Feng, M.; Wei, J.; Han, Q.; Zhao, H.; Li, G.; Zhu, Z.; Xing, H.; An, Y.; Qin, C.; et al. Transplantation of Flk-1+ human bone marrow-derived mesenchymal stem cells promote angiogenesis and neurogenesis after cerebral ischemia in rats. Eur. J. Neurosci. 2011, 34, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Bedi, S.S.; Walker, P.A.; Shah, S.K.; Jimenez, F.; Thomas, C.P.; Smith, P.; Hetz, R.A.; Xue, H.; Pati, S.; Dash, P.K.; et al. Autologous bone marrow mononuclear cells therapy attenuates activated microglial/macrophageresponse and improves spatial learning after traumatic brain injury. J. Trauma Acute Care Surg. 2013, 75, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Urao, N.; Ushio-Fukai, M. Redox regulation of stem/progenitor cells and bone marrow niche. Free Rad. Biol. Med. 2013, 54, 26–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, J.M.; Lee, H.J.; Jung, Y.H.; Lee, K.H.; Kim, D.I.; Kim, J.Y.; Ko, S.H.; Choi, G.E.; Chai, I.I.; Song, E.J.; et al. Regulation of stem cell fate by ROS-mediated alteration of metabolism. Int. J. Stem Cells 2015, 8, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Brenneman, M.; Sharma, S.; Harting, M.; Strong, R.; Cox, C.S., Jr.; Aronowski, J.; Grotta, J.C.; Savitz, S.I. Autologous bone marrow mononuclear cells enhance recovery after acute ischemic stroke in young and middle-aged rats. J. Cereb. Blood Flow Metab. 2010, 30, 140–149. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Nagai, A.; Sheikh, A.M.; Shiota, Y.; Narantuya, D.; Watanabe, T.; Masuda, J.; Kobayashi, S.; Kim, S.U.; Yamaguchi, S. Transplantation of human mesenchymal stem cells promotes functional improvement and increased expression of neurotrophic factors in a rat focal cerebral ischemia model. J. Neurosci. Res. 2010, 88, 1017–1025. [Google Scholar] [CrossRef]
- Chen, M.F.; Lin, C.T.; Chen, W.C.; Yang, C.T.; Chen, C.C.; Liao, S.K.; Liu, J.M.; Lu, C.H.; Lee, K.D. The sensitivity of human mesenchymal stem cells to ionizing radiation. Int. J. Radiat. Oncol. Biol. Phys. 2006, 66, 244–253. [Google Scholar] [CrossRef]
- Valle-Prieto, A.; Conget, P.A. Human Mesenchymal Stem Cells Efficiently Manage Oxidative Stress. Stem Cells Dev. 2010, 19, 1885–1893. [Google Scholar] [CrossRef]
- Grisendi, G.; Annerén, C.; Cafarelli, L.; Sternieri, R.; Veronesi, E.; Cervo, G.L.; Luminari, S.; Maur, M.; Frassoldati, A.; Palazzi, G.; et al. GMP-manufactured density gradient media for optimized mesenchymal stromal/stem cell isolation and expansion. Cytotherapy 2010, 12, 466–477. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F. Minimal criteria for defining multipotent mesenchymal stromal cells. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Maehly, A.C.; Chance, B. The assay of catalases and peroxidases. Methods Biochem. Anal. 1954, 1, 357–424. [Google Scholar] [PubMed]
- Shannon, L.M.; Kay, E.; Lew, J.Y. Peroxidase isoenzymes from horseradish roots. I. Isolation and physical properties. J. Biol. Chem. 1966, 241, 2166–2172. [Google Scholar]
- Bayır, H.; Kochanek, P.M.; Kagan, V.E. Oxidative Stress in Immature Brain after Traumatic Brain Injury. Dev. Neurosci. 2006, 28, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Lagowska-Lenard, M.; Bielewicz, J.; Raszewski, G.; Stelmasiak, Z.; Bartosik-Psujek, H. Oxidative stress in cerebral stroke. Pol. Merk. Lek. 2008, 147, 205. [Google Scholar]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Liu, Y.; Wong, N.K.; Xiao, J.; So, K.F. Oxidative Stress in Stem Cell Aging. Cell Transplant. 2017, 26, 1483–1495. [Google Scholar] [CrossRef]
- Carvalho, A.N.; Firuzi, O.; Gama, M.J.; Horssen, J.V.; Saso, L. Oxidative Stress and Antioxidants in Neurological Diseases: Is There Still Hope? Curr. Drug Targets 2017, 18, 705–718. [Google Scholar] [CrossRef]
- Rebec, G.V.; Pierce, R.C. A vitamin as neuromodulator: Ascorbate release into the extracellular fluid of the brain regulates dopaminergic and glutamatergic transmission. Prog. Neurobiol. 1994, 43, 537–565. [Google Scholar] [CrossRef]
- Karanth, S.; Yu, W.H.; Walczewska, A.; Mastronardi, C.; McCann, S.M. Ascorbic acid acts as an inhibitory transmitter in the hypothalamus to inhibit stimulated luteinizing hormone-releasing hormone release by scavenging nitric oxide. Proc. Natl. Acad. Sci. USA 2000, 97, 1891–1896. [Google Scholar] [CrossRef] [Green Version]
- Qiu, S.; Li, L.; Weeber, E.J.; May, J.M. Ascorbate transport by primary cultured neurons and its role in neuronal function and protection against excitotoxicity. J. Neurosci. Res. 2007, 85, 1046–1056. [Google Scholar] [CrossRef] [PubMed]
- Pastor, P.; Cisternas, P.; Salazar, K.; Silva-Alvarez, C.; Oyarce, K.; Jara, N.; Espinoza, F.; Martínez, A.D.; Nualart, F. SVCT2 vitamin C transporter expression in progenitor cells of the postnatal neurogenic niche. Front Cell Neurosci. 2013, 7, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farbstein, D.; Kozak -Blickstein, A.; Levy, A.P. Antioxidant vitamins and their use in preventing cardiovascular disease. Molecules 2010, 15, 8098–8110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spector, R.; Lorenzo, A.V. Ascorbic acid homeostasis in the central nervous system. Am. J. Physiol. 1973, 225, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Englard, S.; Seifter, S. The biochemical functions of ascorbic acid. Annu. Rev. Nutr. 1986, 6, 365–406. [Google Scholar] [CrossRef] [PubMed]
- Rice, M.E. Ascorbate regulation and its neuroprotective role in the brain. Trends Neurosci. 2000, 23, 209–216. [Google Scholar] [CrossRef]
- DeMenezes, C.C.; Dorneles, A.G.; Sperotto, R.L.; Duarte, M.M.; Schetinger, M.R.; Loro, V.L. Oxidative stress in cerebrospinal fluid of patients with aseptic and bacterial meningitis. Neurochem. Res. 2009, 34, 1255–1260. [Google Scholar] [CrossRef]
- Bowman, G.L.; Dodge, H.; Frei, B.; Calabrese, C.; Oken, B.S.; Kaye, J.A.; Quinn, J.F. Ascorbic acid and rates of cognitive decline in Alzheimer’s disease. J. Alzheimers Dis. 2009, 16, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Arlt, S.; Müller-Thomsen, T.; Beisiegel, U.; Kontush, A. Effect of one-year vitamin C- and E-supplementation on cerebrospinal fluid oxidation parametersand clinical course in Alzheimer’s disease. Neurochem. Res. 2012, 37, 2706–2714. [Google Scholar] [CrossRef]
- Prasad, R.; Mishra, O.P.; Mishra, S.P.; Upadhyay, R.S.; Singh, T.B. Oxidative stress in children with neurocysticercosis. Pediatr. Infect. Dis. J. 2012, 31, 1012–1015. [Google Scholar] [CrossRef]
- Arlt, S.; Kontush, A.; Zerr, I.; Buhmann, C.; Jacobi, C.; Schröter, A.; Poser, S.; Beisiegel, U. Increased lipid peroxidation in cerebrospinal fluid and plasma from patients with Creutzfeldt-Jakob disease. Neurobiol. Dis. 2002, 10, 150–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voigt, K.; Kontush, A.; Stuerenburg, H.J.; Muench-Harrach, D.; Hansen, H.C.; Kunze, K. Decreased plasma and cerebrospinal fluid ascorbate levels in patients with septic encephalopathy. Free Radic Res. 2002, 36, 735–739. [Google Scholar] [CrossRef] [PubMed]
- Paraskevas, G.P.; Kapaki, E.; Libitaki, G.; Zournas, C.; Segditsa, I.; Papageorgiou, C. Ascorbate in healthy subjects, amyotrophic lateral sclerosis and Alzheimer’s disease. Acta Neurol. Scand. 1997, 96, 88–90. [Google Scholar] [CrossRef] [PubMed]
- Buhmann, C.; Arlt, S.; Kontush, A.; Möller-Bertram, T.; Sperber, S.; Oechsner, M.; Stuerenburg, H.J.; Beisiegel, U. Plasma and CSF markers of oxidative stress are increased in Parkinson’s disease and influenced by antiparkinsonian medication. Neurobiol. Dis. 2004, 15, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Reiber, H.; Ruff, M.; Uhr, M. Ascorbate concentration in human cerebrospinal fluid (CSF) and serum. Intrathecal accumulation and CSF flow rate. Clin. Chim. Acta 1993, 217, 163–173. [Google Scholar] [CrossRef]
- Tallaksen, C.M.; Bøhmer, T.; Bell, H. Concentrations of the water-soluble vitamins thiamin, ascorbic acid, and folic acid in serum and cerebrospinal fluid of healthy individuals. Am. J. Clin. Nutr. 1992, 56, 559–564. [Google Scholar] [CrossRef]
- Brau, R.H.; García-Castiñeiras, S.; Rifkinson, N. Cerebrospinal fluid ascorbic acid levels in neurological disorders. Neurosurgery 1984, 14, 142–146. [Google Scholar] [CrossRef]
- Polidori, M.C.; Mecocci, P.; Frei, B. Plasma vitamin C levels are decreased and correlated with brain damage in patients with intracranial hemorrhage or head trauma. Stroke 2001, 32, 898–902. [Google Scholar] [CrossRef] [Green Version]
- Rice, M.E.; Russo-Menna, I. Differential compartmentalization of brain ascorbate and glutathione between neurons and glia. Neuroscience 1998, 82, 1213–1223. [Google Scholar] [CrossRef]
- Nayak, C.; Nayak, D.; Raja, A.; Rao, A. Time-level relationship between indicators of oxidative stress and Glasgow Coma Scale scores of severe head injury patients. Clin. Chem. Lab. Med. 2006, 44, 460–463. [Google Scholar] [CrossRef]
- Alho, H.; Leinonen, J.S.; Erhola, M.; Lönnrot, K.; Aejmelaeus, R. Assay of antioxidant capacity of human plasma and CSF in aging and disease. Restor. Neurol. Neurosci. 1998, 12, 159–165. [Google Scholar] [PubMed]
- Liu, J.T.; Tan, W.C.; Liao, W.J. Effects of electrical cervical spinal cord stimulation on cerebral blood perfusion, cerebrospinal fluid catecholamine levels, and oxidative stress in comatose patients. Acta Neurochir. Suppl. 2008, 101, 71–76. [Google Scholar] [PubMed]
- Spector, R.; Spector, A.Z.; Snodgrass, S.R. Model for transport in the central nervous system. Am. J. Physiol. 1977, 232, R73–R79. [Google Scholar] [CrossRef] [PubMed]
- Spector, R.; Johanson, C.E. The nexus of vitamin homeostasis and DNA synthesis and modification in mammalian brain. Mol. Brain 2014, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Angelow, S.; Haselbach, M.; Galla, H.J. Functional characterisation of the active ascorbic acid transport into cerebrospinal fluid using primary cultured choroid plexus cells. Brain Res. 2003, 988, 105–113. [Google Scholar] [CrossRef]
- Nualart, F.; Salazar, K.; Oyarce, K.; Cisternas, P.; Jara, N.; Silva-Álvarez, C.; Pastor, P.; Martínez, F.; García, A.; García-Robles Mde, L.; et al. Typical and atypical stem cells in the brain, vitamin C effect and neuropathology. Biol. Res. 2012, 45, 243–256. [Google Scholar] [CrossRef]
- May, J.M. Vitamin C transport and its role in the central nervous system. Subcell. Bochem. 2012, 56, 85–103. [Google Scholar]
- Vera, J.C.; Rivas, C.I.; Fischbarg, J.; Golde, D.W. Mammalian facilitative hexose transporters mediate the transport of dehydroascorbic acid. Nature 1993, 364, 79–82. [Google Scholar] [CrossRef]
- Rumsey, S.C.; Kwon, O.; Xu, G.W.; Burant, C.F.; Simpson, I.; Levine, M. Glucose transporter isoforms GLUT1 and GLUT3 transport dehydroascorbic acid. J. Biol. Chem. 1997, 272, 18982–18989. [Google Scholar] [CrossRef] [Green Version]
- Cisternas, P.; Silva-Alvarez, C.; Martinez, F.; Fernandez, E.; Ferrada, L.; Oyarce, K.; Salazar, K.; Bolaños, J.P.; Nualart, F. The oxidized form of vitamin C, dehydroascorbic acid, regulates neuronal energy metabolism. J. Neurochem. 2014, 129, 663–671. [Google Scholar] [CrossRef]
- Astuya, A.; Caprile, T.; Castro, M.; Salazar, K.; de Garcia, M.L.; Reinicke, K.; Rodríguez, F.; Vera, J.C.; Millán, C.; Ulloa, V.; et al. Vitamin C uptake and recycling among normal and tumor cells from the central nervous system. J. Neurosci. Res. 2005, 79, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.A.; Pozo, M.; Cortes, C.; de Garcia, M.L.; Concha, I.I.; Nualart, F. Intracellular ascorbic acid inhibits transport of glucose by neurons, but not by astrocytes. J. Neurochem. 2007, 102, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Arlt, S.; Müller-Thomsen, T.; Beisiegel, U. Use of Vitamin C and E in the Treatment of Alzheimer’s Disease. Drug Dev. Res. 2002, 56, 452–457. [Google Scholar] [CrossRef]
- Schippling, S.; Kontush, A.; Arlt, S.; Buhmann, C.; Stürenburg, H.J.; Mann, U.; Müller-Thomsen, T.; Beisiegelm, U. Increased lipoprotein oxidation in Alzheimer’s disease. Free Radic Biol. Med. 2000, 28, 351–360. [Google Scholar] [CrossRef]
- Aygul, R.; Kotan, D.; Demirbas, F.; Ulvi, H.; Deniz, O. Plasma oxidants and antioxidants in acute ischemic stroke. J. Int. Med. Res. 2006, 34, 413–418. [Google Scholar] [CrossRef]
- Warner, D.S.; Sheng, H.; Batinic-Haberle, I. Oxidants, antioxidants and the ischemic brain. J. Exp. Biol. 2004, 207, 3221–3231. [Google Scholar] [CrossRef] [Green Version]
- Jacobsson, J.; Jonsson, P.A.; Andersen, P.M.; Forsgren, L.; Marklund, S.L. Superoxide dismutase in CSF from amyotrophic lateral sclerosis patients with and without CuZn-superoxide dismutase mutations. Brain 2001, 124, 1461–1466. [Google Scholar] [CrossRef] [Green Version]
- Younus, H. Therapeutic potentials of superoxide dismutase. Int. J. Health Sci. (Qassim) 2018, 12, 88–93. [Google Scholar]
- Strand, T.; Marklund, S.L. Release of Superoxide Dismutase into Cerebrospinal Fluid as a Marker of Brain Lesion in Acute Cerebral Infarction. Stroke 1992, 23, 515–518. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, E.; Mokuno, K.; Aoki, S.; Takahashi, A.; Riku, S.; Murayama, T.; Yanagi, T.; Kato, K. Cerebrospinal fluid levels of superoxide dismutases in neurological diseases detected by sensitive enzyme immunoassays. J. Neurol. Sci. 1994, 124, 25–31. [Google Scholar] [CrossRef]
- Kemp, K.; Hares, K.; Mallam, E.; Heesom, K.J.; Scolding, N.; Wilkins, A. Mesenchymal stem cell-secreted superoxide dismutase promotes cerebellar neuronal survival. J. Neurochem. 2010, 114, 1569–1580. [Google Scholar] [CrossRef] [PubMed]
- Whone, A.L.; Kemp, K.; Suna, M.; Wilkins, A.; Scolding, N.J. Human bone marrow mesenchymal stem cells protect catecholaminergic and serotonergic neuronal perikarya and transporter function from oxidative stress by the secretion of glial-derived neurotrophic factor. Brain Res. 2012, 1431, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Alhazzani, A.; Rajagopalan, P.; Albarqi, Z.; Devaraj, A.; Mohamed, M.H.; Al-Hakami, A.; Chandramoorthy, H.C. Mesenchymal Stem Cells (MSCs) Coculture Protects [Ca2+]i Orchestrated Oxidant Mediated Damage in Differentiated Neurons In Vitro. Cells 2018, 7, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapoynikova, N.; Asatiani, N.; Kartvelishvili, T.; Kalandadze, I.; Tsiskaridze, A. Plasma Antioxidant Activity as a Marker for a Favourable Outcome in Acute Ischemic Stroke. In Antioxidant Enzyme; El-Missiry, M.A., Ed.; IntechOpen: London, UK, 2012. [Google Scholar]
- Spranger, M.; Krempien, S.; Schwab, S.; Donneberg, S.; Werner, H. Superoxide Dismutase Activity in Serum of Patients with Acute Cerebral Ischemic Injury. Correlation with Clinical Course and Infarct Size. Stroke 1997, 28, 2425–2428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaynar, M.Y.; Taner, T.; Rahsan, K.; Pinar, A.; Gumustas, K. Cerebrospinal fluid superoxide dismutase and serum malondialdehyde levels in patients with aneurysmal subarachnoid hemorrhage: Preliminary results. Neurol. Res. 2005, 27, 562–567. [Google Scholar] [CrossRef]
- Bayır, A.; Kıyıcı, A. The Relation between Glasgow Coma Score and Blood Superoxide Dismutase Activity in Patients with Traumatic Brain Injury. J. Head Trauma Rehabil. 2011, 26, 406. [Google Scholar]
- Gruener, N.; Gross, B.; Gozlan, O.; Barak, M. Increase in superoxide dismutase after cerebrovascular accident. Life Sci. 1994, 54, 711–713. [Google Scholar] [CrossRef]
- Baud, O.; Greene, A.E.; Li, J.; Wang, H.; Volpe, J.J. Rosenberg PA. Glutathione peroxidase-catalase cooperativity is required for resistance to hydrogen peroxide by mature rat oligodendrocytes. J. Neurosci. 2004, 24, 1531–1540. [Google Scholar] [CrossRef] [Green Version]
- Kodydková, J.; Vávrová, L.; Kocík, M.; Žák, A. Human catalase, its polymorphisms, regulation and changes of its activity in different diseases. Folia. Biol. (Praha) 2014, 60, 153–167. [Google Scholar]
- Cornelius, C.; Crupi, R.; Calabrese, V.; Graziano, A.; Milone, P.; Pennisi, G.; Radak, Z.; Calabrese, E.J.; Cuzzocrea, S. Traumatic brain injury: Oxidative stress and neuroprotection. Antioxid. Redox Signal. 2013, 19, 836–853. [Google Scholar] [CrossRef]
- Gebicka, L.; Krych-Madej, J. The role of catalases in the prevention/promotion of oxidative stress. J. Inorg. Biochem. 2019, 197, 110699. [Google Scholar] [CrossRef] [PubMed]
- Da Costa Gonçalves, F.; Grings, M.; Nunes, N.S.; Pinto, F.O.; Garcez, T.N.; Visioli, F.; Leipnitz, G.; Paz, A.H. Antioxidant properties of mesenchymal stem cells against oxidative stress in a murine model of colitis. Biotechnol. Lett. 2017, 39, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Singhal, A.; Morris, V.B.; Labhasetwar, V.; Ghorpade, A. Nanoparticle-mediated catalase delivery protects human neurons from oxidative stress. Cell Death Dis. 2013, 4, e903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanza, C.; Morando, S.; Voci, A.; Canesi, L.; Principato, M.C.; Serpero, L.D.; Mancardi, G.; Uccelli, A.; Vergani, L. Neuroprotective mesenchymal stem cells are endowed with a potent antioxidant effect in vivo. J. Neurochem. 2009, 110, 1674–1684. [Google Scholar] [CrossRef]
- Yoo, D.Y.; Kim, D.W.; Chung, J.Y.; Jung, H.Y.; Kim, J.W.; Yoon, Y.S.; Hwang, I.K.; Choi, J.H.; Choi, G.M.; Choi, S.Y.; et al. Cu, Zn-Superoxide Dismutase Increases the Therapeutic Potential of Adipose-derived Mesenchymal Stem Cells by Maintaining Antioxidant Enzyme Levels. Neurochem. Res. 2016, 41, 3300–3307. [Google Scholar] [CrossRef]
- Xu, J.; Huang, Z.; Lin, L.; Fu, M.; Gao, Y.; Shen, Y.; Zou, Y.; Sun, A.; Qian, J.; Ge, J. miR-210 over-expression enhances mesenchymal stem cell survival in an oxidative stress environment through antioxidation and c-Met pathway activation. Sci. China Life Sci. 2014, 57, 989–997. [Google Scholar] [CrossRef] [Green Version]
- Pincherle, J.J.; Chatelle, C.; Pignat, J.M.; Du Pasquier, R.; Ryvlin, P.; Oddo, M.; Diserens, K. Motor behavior unmasks residual cognition in disorders of consciousness. Ann. Neurol. 2019, 85, 443–447. [Google Scholar] [CrossRef]
- Schiff, N.D. Cognitive Motor Dissociation Following Severe Brain Injuries. JAMA Neurol. 2015, 72, 1413–1415. [Google Scholar] [CrossRef]
- Pignat, J.M.; Mauron, E.; Jöhr, J.; Gilart de Keranflec’h, C.; Van De Ville, D.; Preti, M.G.; Meskaldji, D.E.; Hömberg, V.; Laureys, S.; Draganski, B.; et al. Outcome Prediction of Consciousness Disorders in the Acute Stage Based on a Complementary Motor Behavioural Tool. PLoS ONE 2016, 11, e0156882. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient 1 | Patient 2 | Patient 3 | Patient 4 | Patient 5 | Patient 6 | Patient 7 | Patient 8 | Patient 9 | |
|---|---|---|---|---|---|---|---|---|---|
| Age (years) | 22 | 19 | 45 | 19 | 30 | 23 | 36 | 35 | 38 |
| Gender (M/F) | M | M | M | M | F | F | F | F | M |
| Mechanism of MCS induction | Head injury after traffic accident | Head injury after traffic accident | Head injury after traffic accident | Head injury after traffic accident | Head injury after traffic accident | Listeriosis encephalitis with damage to the thalamus and basal ganglia | Ischemia with cardiac arrest | Hypoglycemia after poisoning | Head injury after trafficaccident |
| Time in MCS before the clinical trial (months) | 3 | 8 | 7 | 6 | 12 | 10 | 14 | 6 | 6 |
| Before Treatment | I Application of MSC | II Application of MSC | |||||
|---|---|---|---|---|---|---|---|
| CSF | plasma | CSF | plasma | CSF | plasma | ||
| A | Total antioxidant capacity (nM) | 0.05 ± 0.015 a | 0.39 ± 0.04 b | 0.08 ± 0.014 a | 0.34 ± 0.04 b | 0.08 ± 0.012 a | 0.39 ± 0.04 b |
| B | Ascorbic acid concentration (ng/μL) | 27.25 ± 1.31 a | 9.96 ± 1.44 b | 22.73 ± 1.42 a | 6.39 ± 1.32 b | 28.13 ± 1.22 a | 7.36 ± 1.56 b |
| C | Total ascorbate concentration (nM/μL) | 0.20 ± 0.009 a | 0.39 ± 0.02 b | 0.17 ± 0.008 a | 0.32 ± 0.02 b | 0.19 ± 0.01 a | 0.32 ± 0.02 b |
| D | Superoxide dismutase (SOD) activity (%) | 40.82 ± 2.05 a | 65.66 ± 2.27 b | 37.42 ± 1.97 a | 66.35 ±2.27 b | 36.87 ± 2.11 a | 60.73 ± 2.41 b |
| E | Catalase (CAT) activity (μmol/min/mL) | 11.29 ± 0.60 a | 0.04 ± 0.008 b | 13.20 ± 0.51 a | 0.06 ± 0.008 b | 12.38 ± 0.59 a | 0.03 ± 0.009 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jezierska-Wozniak, K.; Sinderewicz, E.; Czelejewska, W.; Wojtacha, P.; Barczewska, M.; Maksymowicz, W. Influence of Bone Marrow-Derived Mesenchymal Stem Cell Therapy on Oxidative Stress Intensity in Minimally Conscious State Patients. J. Clin. Med. 2020, 9, 683. https://doi.org/10.3390/jcm9030683
Jezierska-Wozniak K, Sinderewicz E, Czelejewska W, Wojtacha P, Barczewska M, Maksymowicz W. Influence of Bone Marrow-Derived Mesenchymal Stem Cell Therapy on Oxidative Stress Intensity in Minimally Conscious State Patients. Journal of Clinical Medicine. 2020; 9(3):683. https://doi.org/10.3390/jcm9030683
Chicago/Turabian StyleJezierska-Wozniak, Katarzyna, Emilia Sinderewicz, Wioleta Czelejewska, Pawel Wojtacha, Monika Barczewska, and Wojciech Maksymowicz. 2020. "Influence of Bone Marrow-Derived Mesenchymal Stem Cell Therapy on Oxidative Stress Intensity in Minimally Conscious State Patients" Journal of Clinical Medicine 9, no. 3: 683. https://doi.org/10.3390/jcm9030683