Distribution Patterns of Odonate Assemblages in Relation to Environmental Variables in Streams of South Korea

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ecological Data
2.2. Data Analysis
3. Results
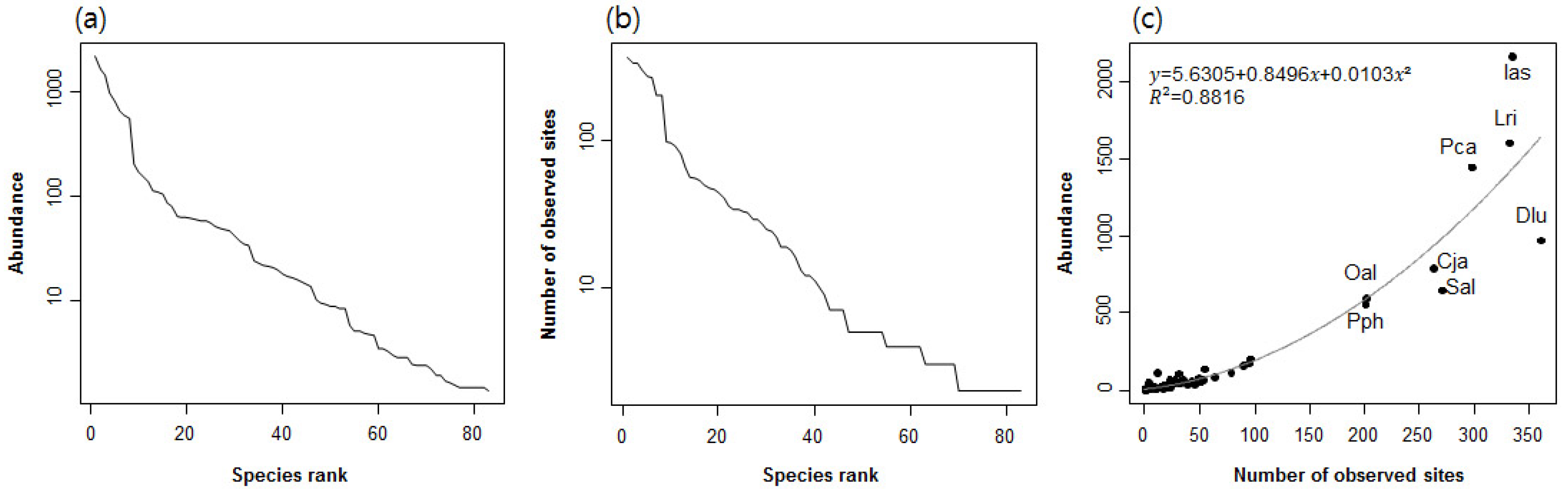
3.1. Distribution Patterns of Species
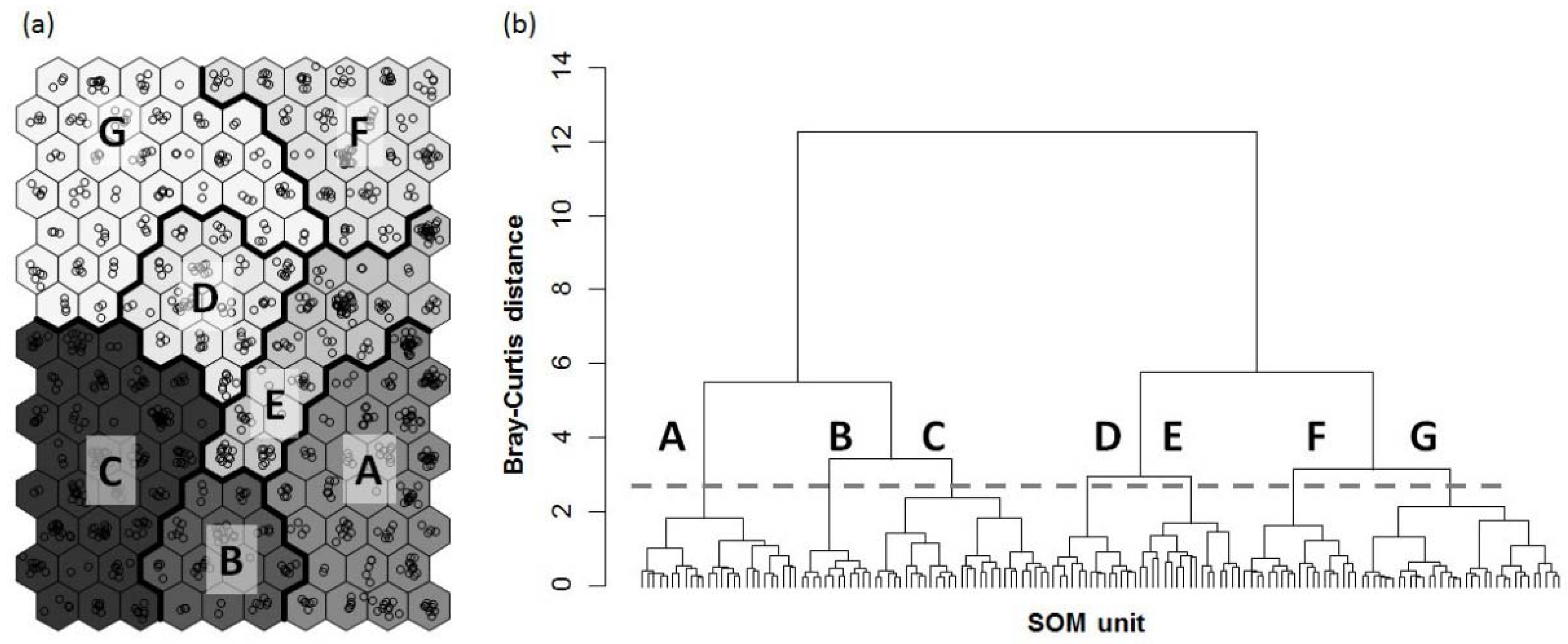
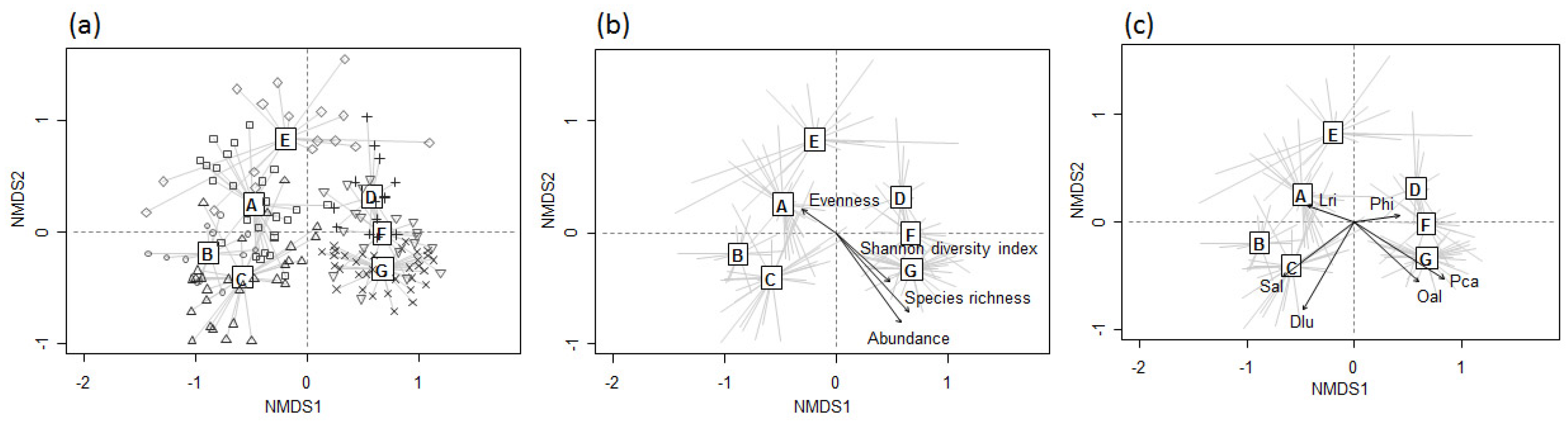
3.2. Patterns in Odonate Assemblages
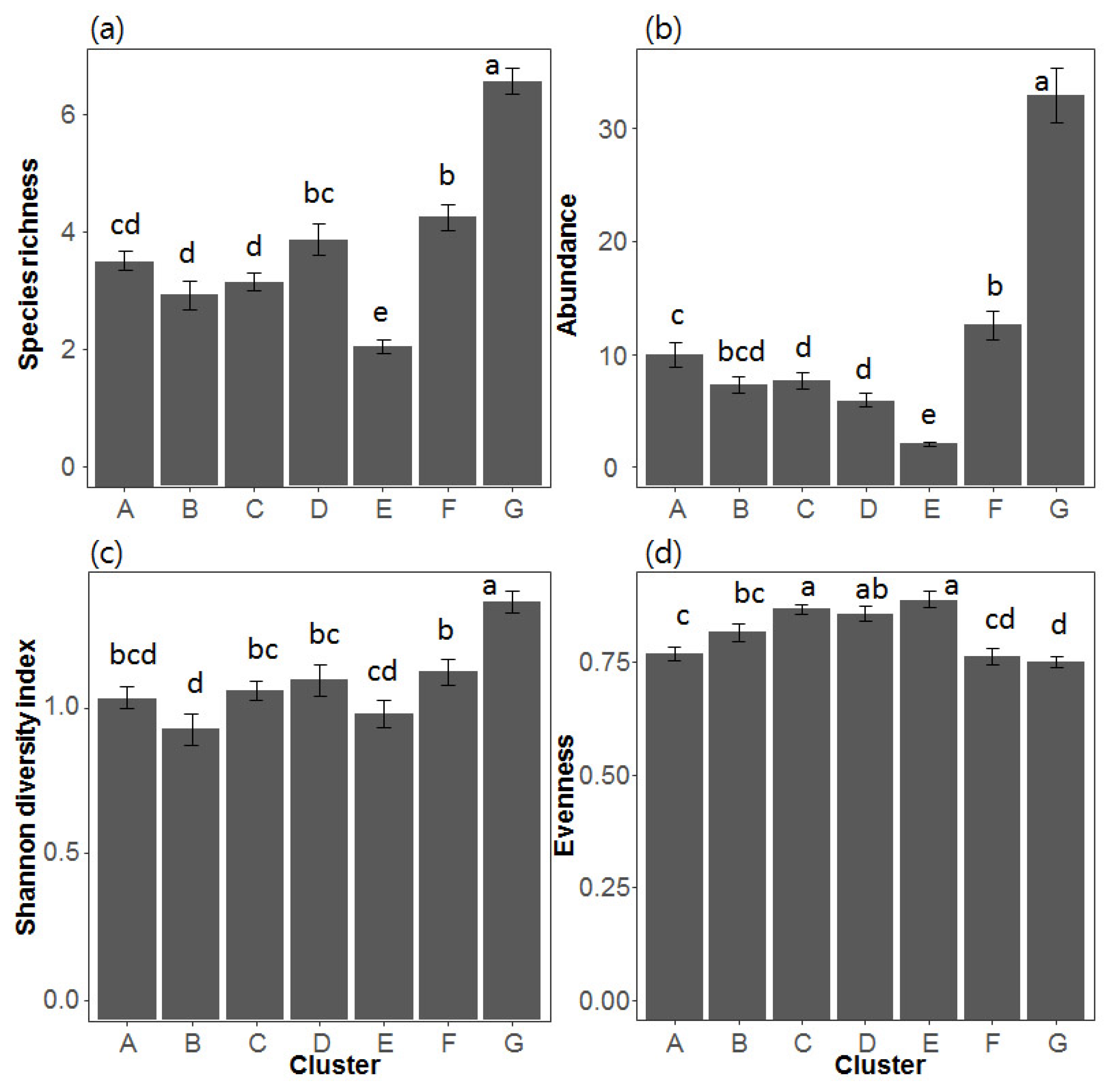
3.3. Differences in Environmental Variables among Different Assemblage Patterns
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Park, Y.-S.; Céréghino, R.; Compin, A.; Lek, S. Applications of artificial neural networks for patterning and predicting aquatic insect species richness in running waters. Ecol. Model. 2003, 160, 265–280. [Google Scholar] [CrossRef]
- Goertzen, D.; Suhling, F. Promoting dragonfly diversity in cities: Major determinants and implications for urban pond design. J. Insect Conserv. 2013, 17, 399–409. [Google Scholar] [CrossRef]
- Corbet, P.S. Dragonflies: Behavior and Ecology of Odonata; Comstock Pub. Associates: Ithaca, NY, USA, 1999; ISBN 978-0-8014-2592-9. [Google Scholar]
- Kalkman, V.J.; Clausnitzer, V.; Dijkstra, K.-D.B.; Orr, A.G.; Paulson, D.R.; van Tol, J. Global diversity of dragonflies (Odonata) in freshwater. In Freshwater Animal Diversity Assessment; Balian, E.V., Lévêque, C., Segers, H., Martens, K., Eds.; Developments in Hydrobiology, Springer Netherlands: Dordrecht, The Netherlands, 2008; pp. 351–363. ISBN 978-1-4020-8259-7. [Google Scholar]
- Osborn, R.; Samways, M.J. Determinants of adult dragonfly assemblage patterns at new ponds in South Africa. Odonatologica 1996, 25, 49–58. [Google Scholar]
- Kutcher, T.E.; Bried, J.T. Adult Odonata conservatism as an indicator of freshwater wetland condition. Ecol. Indic. 2014, 38, 31–39. [Google Scholar] [CrossRef]
- Hassall, C. Odonata as candidate macroecological barometers for global climate change. Freshw. Sci. 2015, 34, 1040–1049. [Google Scholar] [CrossRef]
- Jonsson, M.; Johansson, F.; Karlsson, C.; Brodin, T. Intermediate predator impact on consumers weakens with increasing predator diversity in the presence of a top-predator. Acta Oecol. Int. J. Ecol. 2007, 31, 79–85. [Google Scholar] [CrossRef]
- Brooks, S. Field Guide to the Dragonflies and Damselfies of Great Britain and Ireland; Bloomsbury Publishing Inc.: New York, NY, USA, 2018; ISBN 978-1-4729-6453-3. [Google Scholar]
- Foote, A.L.; Hornung, C.L.R. Odonates as biological indicators of grazing effects on Canadian prairie wetlands. Ecol. Entomol. 2005, 30, 273–283. [Google Scholar] [CrossRef]
- Villalobos-Jimenez, G.; Dunn, A.; Hassall, C. Dragonflies and damselflies (Odonata) in urban ecosystems: A review. Eur. J. Entomol. 2016, 113, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Renner, S.; Périco, E.; Dalzochio, M.S.; Sahlén, G. Water body type and land cover shape the dragonfly communities (Odonata) in the Pampa biome, Rio Grande do Sul, Brazil. J. Insect Conserv. 2018, 22, 113–125. [Google Scholar] [CrossRef]
- Hofmann, T.A.; Mason, C.F. Habitat characteristics and the distribution of Odonata in a lowland river catchment in eastern England. Hydrobiologia 2005, 539, 137–147. [Google Scholar] [CrossRef]
- Sahlén, G.; Ekestubbe, K. Identification of dragonflies (Odonata) as indicators of general species richness in boreal forest lakes. Biodivers. Conserv. 2001, 10, 673–690. [Google Scholar] [CrossRef]
- Ruggiero, A.; Céréghino, R.; Figuerola, J.; Marty, P.; Angélibert, S. Farm ponds make a contribution to the biodiversity of aquatic insects in a French agricultural landscape. C. R. Biol. 2008, 331, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Jung, K. Odonata Larvae of Korea; Nature & Ecology: Seoul, Korea, 2012; ISBN 978-89-962995-6-1. (In Korean) [Google Scholar]
- Park, Y.-S.; Song, M.-Y.; Park, Y.-C.; Oh, K.-H.; Cho, E.; Chon, T.-S. Community patterns of benthic macroinvertebrates collected on the national scale in Korea. Ecol. Model. 2007, 203, 26–33. [Google Scholar] [CrossRef]
- Li, F.; Chung, N.; Bae, M.-J.; Kwon, Y.-S.; Park, Y.-S. Relationships between stream macroinvertebrates and environmental variables at multiple spatial scales. Freshw. Biol. 2012, 57, 2107–2124. [Google Scholar] [CrossRef]
- Ministry of Environment (MOE), National Institute of Environmental Research (NIER). Survey and Evaluation Method for River and Stream Ecosystem Health Assessment; MOE, NIER: Incheon, Korea, 2015. (In Korean) [Google Scholar]
- Bae, M.-J.; Kwon, Y.; Hwang, S.-J.; Chon, T.-S.; Yang, H.-J.; Kwak, I.-S.; Park, J.-H.; Ham, S.-A.; Park, Y.-S. Relationships between three major stream assemblages and their environmental factors in multiple spatial scales. Ann. Limnol. Int. J. Limnol. 2011, 47, S91–S105. [Google Scholar] [CrossRef] [Green Version]
- Ministry of Land, Infrastructure and Transport (MOLIT), Han River Flood Control Office (HRFCO). List of Rivers of South Korea; MOLIT: Sejong, Korea, 2013. (In Korean) [Google Scholar]
- Jeju Development Institute. The Hydrologic and Water Quality Characteristics Analysis for the Streamwater Application in Jeju Island; Jeju Development Institute: Jeju, Korea, 2009. (In Korean) [Google Scholar]
- Environmental Systems Research Incorporated (ESRI). ArcGIS 10.1; Environmental Systems Research Incorporated: Redlands, CA, USA, 2013. [Google Scholar]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater; American Public Health Association/American Water Works Association/Water Environment Federation: Washington, DC, USA, 2005. [Google Scholar]
- Cummins, K.W.; Lauff, G.H. The influence of substrate particle size on the microdistribution of stream macrobenthos. Hydrobiologia 1969, 34, 145–181. [Google Scholar] [CrossRef]
- Kohonen, T. Self-organized formation of topologically correct feature maps. Biol. Cybern. 1982, 43, 59–69. [Google Scholar] [CrossRef]
- Park, Y.-S.; Kwon, Y.-S.; Hwang, S.-J.; Park, S. Characterizing effects of landscape and morphometric factors on water quality of reservoirs using a self-organizing map. Environ. Model. Softw. 2014, 55, 214–221. [Google Scholar] [CrossRef]
- Céréghino, R.; Park, Y.-S. Review of the Self-Organizing Map (SOM) approach in water resources: Commentary. Environ. Model. Softw. 2009, 24, 945–947. [Google Scholar] [CrossRef]
- Vesanto, J. Neural Network Tool for Data Mining: SOM Toolbox. In Proceedings of the TOOLMET 2000—3rd International Symbosium on Tool Environments and Development Methods for Intelligent Systems, Oulu, Finland, 13–14 April 2000; pp. 152–169. [Google Scholar]
- Park, Y.-S.; Tison, J.; Lek, S.; Giraudel, J.-L.; Coste, M.; Delmas, F. Application of a self-organizing map to select representative species in multivariate analysis: A case study determining diatom distribution patterns across France. Ecol. Inform. 2006, 1, 247–257. [Google Scholar] [CrossRef]
- Dufrene, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.D.; Jansen, F. Indicspecies: Relationship between Species and Groups of Sites. R Package Version 1.7.6. Available online: https://cran.r-project.org/web/packages/indicspecies/ (accessed on 24 October 2018).
- Shepard, R.N. The analysis of proximities: Multidimensional scaling with an unknown distance function. II. Psychometrika 1962, 27, 219–246. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011; Available online: http://www.R-project.org (accessed on 6 September 2017).
- Pohlert, T. PMCMR: Calculate Pairwise Multiple Comparisons of Mean Rank Sums. R Package Version 4.3. Available online: https://CRAN.R-project.org/package=PMCMR (accessed on 24 October 2018).
- Wehrens, R.; Kruisselbrink, J. kohonen: Supervised and Unsupervised Self-Organising Maps. R Package Version 3.0.6. Available online: https://CRAN.R-project.org/package=kohonen (accessed on 24 October 2018).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-2. Available online: https://CRAN.R-project.org/package=vegan (accessed on 24 October 2018).
- Clausnitzer, V.; Kalkman, V.J.; Ram, M.; Collen, B.; Baillie, J.E.M.; Bedjanič, M.; Darwall, W.R.T.; Dijkstra, K.-D.B.; Dow, R.; Hawking, J.; et al. Odonata enter the biodiversity crisis debate: The first global assessment of an insect group. Biol. Conserv. 2009, 142, 1864–1869. [Google Scholar] [CrossRef]
- Bae, M.-J.; Chun, J.; Chon, T.-S.; Park, Y.-S.; Bae, M.-J.; Chun, J.H.; Chon, T.-S.; Park, Y.-S. Spatio-Temporal Variability in Benthic Macroinvertebrate Communities in Headwater Streams in South Korea. Water 2016, 8, 99. [Google Scholar] [CrossRef]
- Lee, D.-Y.; Bae, M.-J.; Kwon, Y.-S.; Park, C.-W.; Yang, H.M.; Shin, Y.; Kwon, T.-S.; Park, Y.-S. Characteristics of Spatiotemporal Patterns in Benthic Macroinvertebrate Communities in Two Adjacent Headwater Streams. Korean J. Ecol. Environ. 2018, 51, 192–203. [Google Scholar] [CrossRef]
- Suh, A.N.; Samways, M.J. Significance of temporal changes when designing a reservoir for conservation of dragonfly diversity. Biodivers. Conserv. 2005, 14, 165–178. [Google Scholar] [CrossRef]
- Van de Meutter, F.; Meester, L.D.; Stoks, R. Water turbidity affects predator–prey interactions in a fish–damselfly system. Oecologia 2005, 144, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, P. Predation by Enallagma nymphs (Odonata, Zygoptera) under different conditions of spatial heterogeneity. Hydrobiologia 1997, 356, 1–9. [Google Scholar] [CrossRef]
- Jung, K. Odonata of Korea; Ilgongyuksa: Seoul, Korea, 2007; ISBN 978-89-958060-3-6. (In Korean) [Google Scholar]
- Ward, L.; Mill, P.J. Substrate selection in larval Calopteryx splendens (Harris) (Zygoptera: Calopterygidae). Odonatologica 2008, 37, 69–77. [Google Scholar]
- Samejima, Y.; Tsubaki, Y. Body temperature and body size affect flight performance in a damselfly. Behav. Ecol. Sociobiol. 2010, 64, 685–692. [Google Scholar] [CrossRef]
- Tsubaki, Y.; Samejima, Y.; Siva-Jothy, M.T. Damselfly females prefer hot males: Higher courtship success in males in sunspots. Behav. Ecol. Sociobiol. 2010, 64, 1547–1554. [Google Scholar] [CrossRef]
- Kalcounis-Rueppell, M.C.; Payne, V.H.; Huff, S.R.; Boyko, A.L. Effects of wastewater treatment plant effluent on bat foraging ecology in an urban stream system. Biol. Conserv. 2007, 138, 120–130. [Google Scholar] [CrossRef]
- Harabis, F.; Dolny, A. Ecological factors determining the density-distribution of Central European dragonflies (Odonata). Eur. J. Entomol. 2010, 107, 571–577. [Google Scholar] [CrossRef] [Green Version]
- Sahlén, G.; Haase, S.; Suhling, F. Morphology of dragonfly larvae along a habitat gradient: Interactions with feeding behaviour and growth (Odonata: Libellulidae). Int. J. Odonatol. 2008, 11, 225–240. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Variable (Unit) | Abbreviation | Mean | SE * | Range |
|---|---|---|---|---|---|
| Geography | Stream order | Sord | 5.0 | 0.05 | 1.0–9.0 |
| Altitude (m) | 109.8 | 3.67 | 0.0–680.4 | ||
| Slope (°) | 1.3 | 0.14 | 0.0–38.8 | ||
| Meteorology | Total annual precipitation (mm) | Precipitation | 1242.4 | 3.64 | 955.7–1773.9 |
| Annual average temperature (°C) | Tavg | 11.1 | 0.05 | 5.7–15.4 | |
| Maximum temperature in July (°C) | Tmax | 27.5 | 0.04 | 23.1–29.1 | |
| Minimum temperature in January (°C) | Tmin | –8.2 | 0.09 | –14.3–3.1 | |
| Land use | Urban (%) | 12.2 | 0.56 | 0.0–93.4 | |
| Forest (%) | 40.8 | 0.91 | 0.0–100.0 | ||
| Agricultural land (%) | Agriculture | 32.2 | 0.71 | 0.0–94.9 | |
| Grassland (%) | 4.1 | 0.15 | 0.0–36.2 | ||
| Wetland (%) | 2.7 | 0.10 | 0.0–28.1 | ||
| Bareland (%) | 3.3 | 0.15 | 0.0–50.5 | ||
| Waterside (%) | 4.8 | 0.23 | 0.0–99.5 | ||
| Substrate composition ** | Silt (%) | 11.3 | 0.60 | 0.0–100.0 | |
| Fine sand (%) | Fsand | 23.1 | 0.55 | 0.0–91.9 | |
| Coarse sand (%) | Csand | 18.9 | 0.26 | 0.0–65.0 | |
| Gravel (%) | 21.2 | 0.32 | 0.0–65.0 | ||
| Cobble (%) | 17.8 | 0.35 | 0.0–50.0 | ||
| Boulder (%) | 7.6 | 0.31 | 0.0–60.0 | ||
| Hydrology | Water width (m) | Width | 57.0 | 3.03 | 1.0–1400.0 |
| Water depth (cm) | Depth | 30.7 | 0.45 | 9.9–149.2 | |
| Current velocity (cm/s) | Velocity | 32.0 | 0.73 | 0.0–106.6 | |
| Percentage of riffle (%) | Riffle | 19.5 | 0.62 | 0.0–100.0 | |
| Percentage of run (%) | Run | 58.8 | 0.98 | 0.0–100.0 | |
| Percentage of pool (%) | Pool | 21.8 | 0.93 | 0.0–100.0 | |
| Physiochemistry | Dissolve oxygen (mg/L) | DO | 8.8 | 0.04 | 3.2–12.9 |
| (Water quality) | Biochemical oxygen demand (mg/L) | BOD | 1.9 | 0.04 | 0.6–10.8 |
| Total Nitrogen (mg/L) | TN | 2.6 | 0.04 | 0.7–10.8 | |
| Total Phosphate (mg/L) | TP | 0.1 | 0.00 | 0.0–1.1 | |
| Chlorophyll-a (μg/m2) | Chl-a | 3.6 | 0.17 | 0.6–90.1 | |
| pH | 7.8 | 0.01 | 6.3–9.2 | ||
| Electric conductivity (μS/cm) | Conductivity | 224.6 | 6.74 | 22.0–2626.0 | |
| Turbidity (NTU) | 12.0 | 0.45 | 0.0–94.3 |
| Suborder | Family | Species | Frequency | Cluster | Stat * | p-Value |
|---|---|---|---|---|---|---|
| Anisoptera | Gomphidae | Lamelligomphus ringens | 332 | A | 0.789 | <0.001 |
| Anisoptera | Gomphidae | Ophiogomphus obscura | 64 | A | 0.332 | <0.001 |
| Anisoptera | Gomphidae | Sieboldius albardae | 271 | B | 0.791 | <0.001 |
| Anisoptera | Corduliidae | Macromia amphigena | 54 | B | 0.246 | 0.003 |
| Anisoptera | Gomphidae | Davidius lunatus | 361 | C | 0.692 | <0.001 |
| Zygoptera | Coenagrionidae | Paracercion hieroglyphicum | 33 | D | 0.259 | <0.001 |
| Anisoptera | Corduliidae | Macromia manchuria | 43 | D | 0.219 | 0.003 |
| Zygoptera | Lestidae | Lestes sponsa | 6 | D | 0.135 | 0.044 |
| Anisoptera | Libellulidae | Orthetrum albistylum | 202 | F | 0.446 | <0.001 |
| Anisoptera | Libellulidae | Pantala flavescens | 23 | F | 0.224 | 0.003 |
| Anisoptera | Libellulidae | Sympetrum parvulum | 6 | F | 0.198 | <0.001 |
| Anisoptera | Libellulidae | Sympetrum kunckeli | 11 | F | 0.139 | 0.046 |
| Zygoptera | Coenagrionidae | Paracercion calamorum | 298 | G | 0.684 | <0.001 |
| Zygoptera | Coenagrionidae | Ischnura asiatica | 334 | G | 0.661 | <0.001 |
| Zygoptera | Calopterygidae | Calopteryx japonica | 263 | G | 0.548 | <0.001 |
| Zygoptera | Platycnemididae | Platycnemis phillopoda | 201 | G | 0.452 | <0.001 |
| Anisoptera | Aeshnidae | Anax parthenope | 79 | G | 0.431 | <0.001 |
| Zygoptera | Platycnemididae | Copera annulata | 95 | G | 0.387 | <0.001 |
| Anisoptera | Libellulidae | Deielia phaon | 49 | G | 0.344 | <0.001 |
| Anisoptera | Libellulidae | Crocothemis servilia | 90 | G | 0.32 | <0.001 |
| Anisoptera | Libellulidae | Libellula quadrimaculata | 28 | G | 0.261 | <0.001 |
| Zygoptera | Coenagrionidae | Enallagma cyathigerum | 33 | G | 0.241 | 0.002 |
| Zygoptera | Calopterygidae | Atrocalopteryx atrata | 96 | G | 0.219 | 0.029 |
| Anisoptera | Aeshnidae | Anax nigrofasciatus | 15 | G | 0.210 | 0.003 |
| Anisoptera | Gomphidae | Shaogomphus postocularis | 32 | G | 0.200 | 0.009 |
| Anisoptera | Libellulidae | Orthetrum lineostigma | 39 | G | 0.180 | 0.024 |
| Anisoptera | Corduliidae | Epitheca marginata | 21 | G | 0.165 | 0.021 |
| Cluster | Species | Environmental Variables | |||||
|---|---|---|---|---|---|---|---|
| Altitude (m) | Temperature (°C) * | Forest (%) | Cobble (%) | Riffle (%) | BOD (mg/L) | ||
| A | Lamelligomphus ringens | 110.0 (4.3) b | −8.9 (0.1) c | 44.2 (1.2) b | 21.0 (0.5) b | 20.6 (0.8) a | 1.5 (0.0) c |
| B | Sieboldius albardae | 147.2 (6.4) a | −9.3 (0.2) c | 58.8 (1.5) a | 23.4 (0.5) a | 24.6 (1.2) a | 1.2 (0.0) e |
| C | Davidius lunatus | 150.8 (7.4) b | −9.0 (0.1) c | 55.4 (1.4) a | 21.7 (0.5) b | 25.2 (1.0) a | 1.4 (0.0) d |
| F | Orthetrum albistylum | 50.9 (3.2) d | −6.9 (0.2) a | 31.0 (1.7) d,e | 11.7 (0.7) d | 11.6 (1.1) c | 2.4 (0.1) a |
| G | Paracercion calamorum | 61.9 (3.9) d | −6.9 (0.1) a | 32.7 (1.4) d | 12.7 (0.6) d | 11.4 (0.9) c | 2.2 (0.1) b |
| G | Ischnura asiatica | 54.0 (3.0) d | −6.7 (0.1) a | 28.0 (1.3) e | 12.0 (0.5) d | 12.9 (1.0) c | 2.4 (0.1) a |
| G | Calopteryx japonica | 90.9 (4.8) c | −7.7 (0.2) b | 38.9 (1.5) c | 17.3 (0.6) c | 18.3 (1.0) b | 1.6 (0.1) c |
| G | Platycnemis phillopoda | 62.2 (4.6) d | −7.2 (0.1) a | 33.1 (1.7) d,e | 12.2 (0.7) d | 11.4 (1.1) c | 2.3 (0.1) a,b |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.-Y.; Lee, D.-S.; Bae, M.-J.; Hwang, S.-J.; Noh, S.-Y.; Moon, J.-S.; Park, Y.-S. Distribution Patterns of Odonate Assemblages in Relation to Environmental Variables in Streams of South Korea. Insects 2018, 9, 152. https://doi.org/10.3390/insects9040152
Lee D-Y, Lee D-S, Bae M-J, Hwang S-J, Noh S-Y, Moon J-S, Park Y-S. Distribution Patterns of Odonate Assemblages in Relation to Environmental Variables in Streams of South Korea. Insects. 2018; 9(4):152. https://doi.org/10.3390/insects9040152
Chicago/Turabian StyleLee, Da-Yeong, Dae-Seong Lee, Mi-Jung Bae, Soon-Jin Hwang, Seong-Yu Noh, Jeong-Suk Moon, and Young-Seuk Park. 2018. "Distribution Patterns of Odonate Assemblages in Relation to Environmental Variables in Streams of South Korea" Insects 9, no. 4: 152. https://doi.org/10.3390/insects9040152