Glycolytic Activities in the Larval Digestive Tract of Trypoxylus dichotomus (Coleoptera: Scarabaeidae)

Abstract
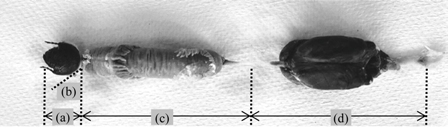
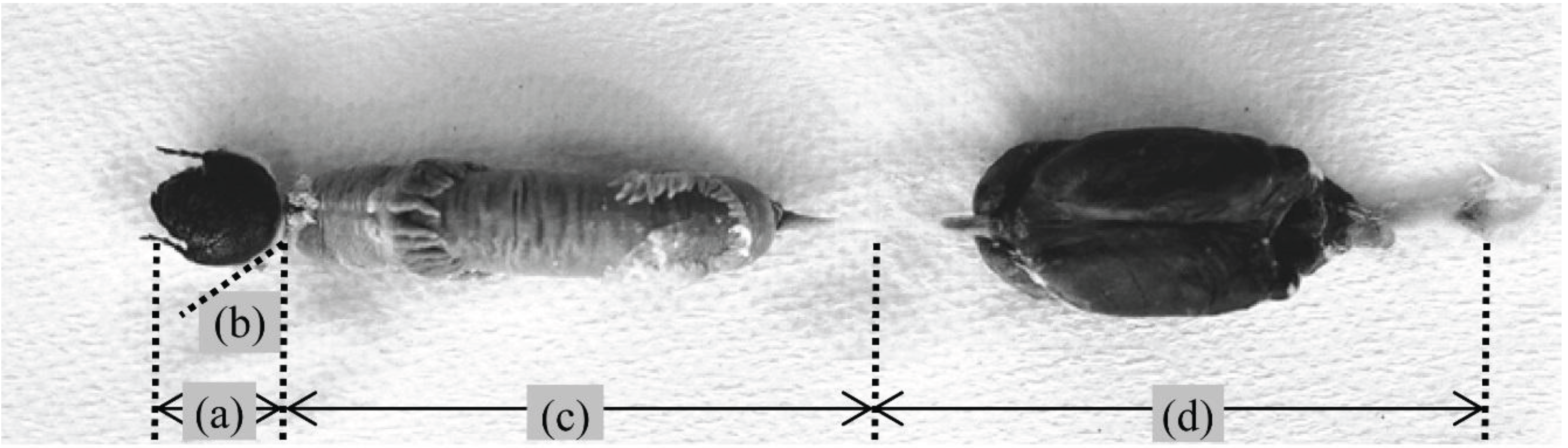
:
1. Introduction
2. Experimental Section
2.1. Insects
2.2. Chemicals
2.3. Gut Extracts

2.4. Glycanase Assays
2.5. Glycosidase Assays
2.6. Analysis of the Neutral Sugar Composition of Larval Food
3. Results and Discussion
3.1. pH of the Gut
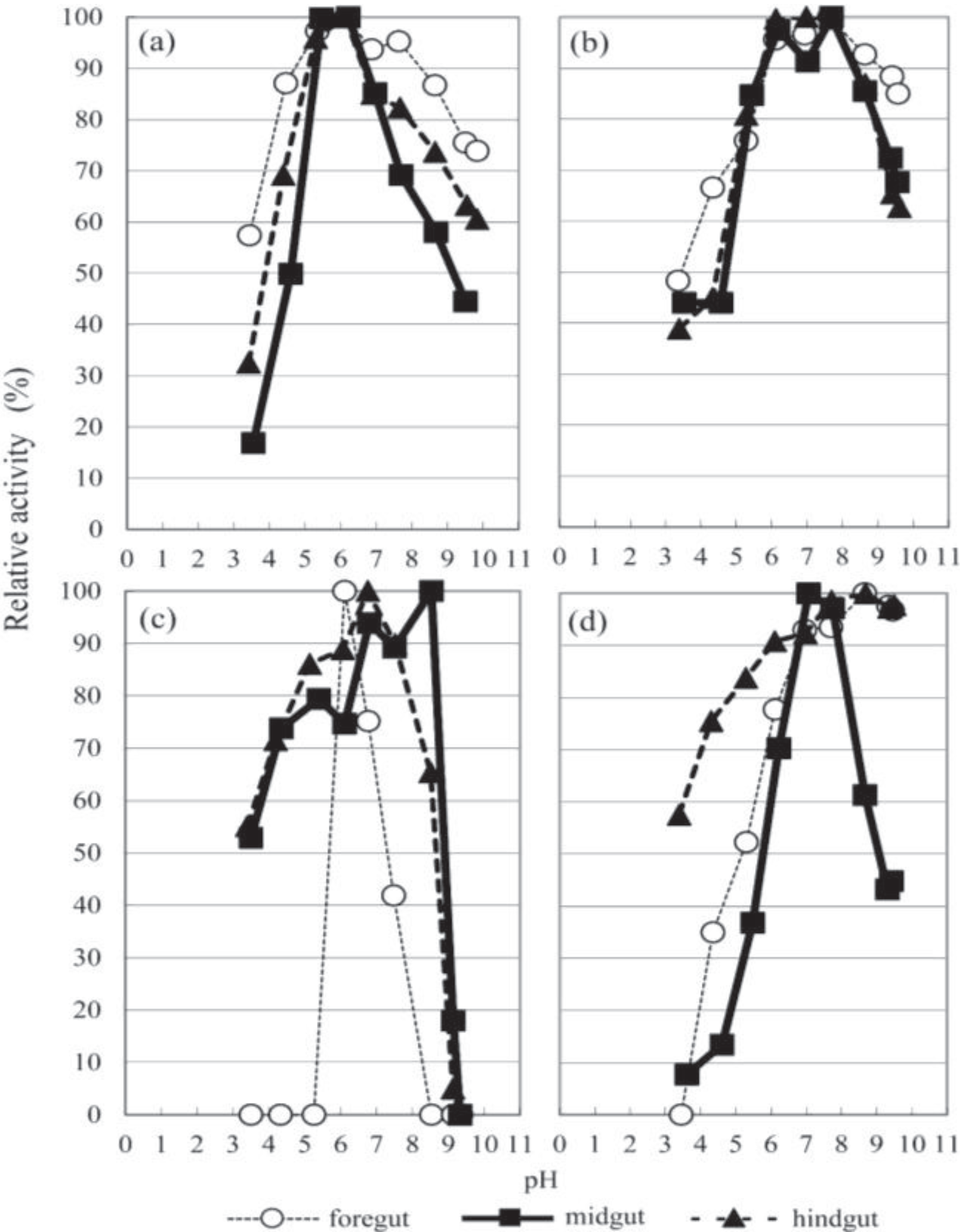
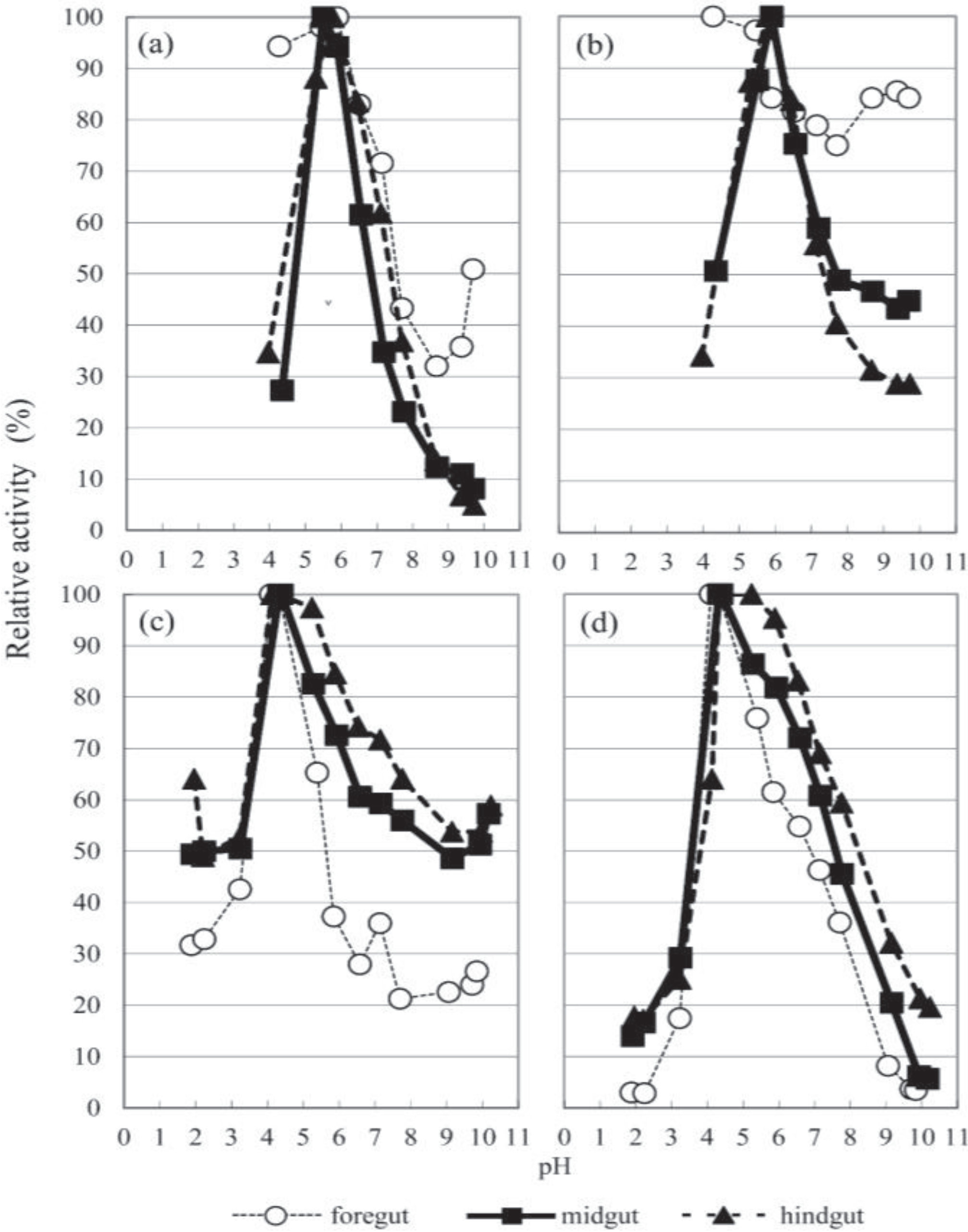
3.2. The Optimal pH
3.3. Glycanase Activities



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | Substrate | Optimum pH | Total Activity (mU) | Specific Activity | |
|---|---|---|---|---|---|
| (mU/mg Protein) | (mU/g Organ) | ||||
| Foregut | CM-cellulose | ND a | 188 b ± 25 c | 4.79 ± 0.55 | 164 ± 23 |
| β-1,3-glucan | 6.15 | 194 ± 24 | 4.95 ± 0.69 | 169 ± 26 | |
| β-1,4-xylan | 7.63 | 1733 ± 415 | 44.25 ± 11.37 | 1514 ± 9 | |
| Pectin | 6.11 | 2540 ± 360 | 64.86 ± 8.20 | 2219 ± 8 | |
| Soluble starch | 8.66 | 127 ± 26 | 3.23 ± 0.63 | 111 ± 19 | |
| Midgut | CM-cellulose | ND a | 319 ± 28 | 8.11 ± 0.77 | 52 ± 6 |
| β-1,3-glucan | 6.22 | 3066 ± 220 | 77.95 ± 4.15 | 502 ± 36 | |
| β-1,4-xylan | 6.18, 7.69 | 7094 ± 543 | 180.36 ± 16.14 | 1162 ± 3 | |
| Pectin | 8.53 | 2982 ± 261 | 75.80 ± 7.18 | 488 ± 1 | |
| Soluble starch | 7.07 | 13,520 ± 569 | 343.74 ± 13.19 | 2214 ± 85 | |
| Hindgut | CM-cellulose | ND a | 80 ± 14 | 5.85 ± 1.02 | 16 ± 4 |
| β-1,3-glucan | 6.14 | 856 ± 87 | 62.63 ± 6.94 | 170 ± 20 | |
| β-1,4-xylan | 6.98 | 2039 ± 311 | 149.19 ± 27.82 | 404 ± 8 | |
| Pectin | 6.76 | 2836 ± 105 | 207.51 ± 13.08 | 562 ± 4 | |
| Soluble starch | 8.69 | 145 ± 36 | 10.61 ± 3.02 | 29 ± 8 | |
| Organ | Substrate | Optimum pH | Total Activity (mU) | Specific Activity | |
|---|---|---|---|---|---|
| (mU/mg Protein) | (mU/g Organ) | ||||
| Foregut | PNP-β-glucoside | 5.89 | 5.09 b ± 0.83 c | 0.13 ± 0.02 | 4.4 ± 0.83 |
| PNP-β-xyloside | N.D. a | 3.10 ± 0.38 | 0.08 ± 0.01 | 2.7 ± 0.41 | |
| PNP-β-mannoside | 4.10 | 34.40 ± 4.25 | 1.75 ± 0.61 | 24.4 ± 3.61 | |
| PNP-β-N-acetylglucosaminide | 4.10 | 1397.83 ± 127.30 | 71.08 ± 7.56 | 992.5 ± 99.64 | |
| Midgut | PNP-β-glucoside | 5.50 | 94.70 ± 3.21 | 2.41 ± 0.13 | 15.5 ± 0.81 |
| PNP-β-xyloside | 5.89 | 25.91 ± 4.23 | 0.66 ± 0.09 | 4.2 ± 0.63 | |
| PNP-β-mannoside | 4.33 | 301.72 ± 50.75 | 18.03 ± 8.35 | 43.1 ± 8.91 | |
| PNP-β-N-acetylglucosaminide | 4.33 | 702.00 ± 51.74 | 41.95 ± 4.12 | 100.3 ± 14.39 | |
| Hindgut | PNP-β-glucoside | 5.77 | 59.04 ± 3.80 | 4.32 ± 0.38 | 11.7 ± 0.54 |
| PNP-β-xyloside | 5.77 | 17.20 ± 2.59 | 1.26 ± 0.18 | 3.4 ± 0.48 | |
| PNP-β-mannoside | 4.13 | 91.18 ± 11.74 | 8.29 ± 1.18 | 15.2 ± 3.60 | |
| PNP-β-N-acetylglucosaminide | 5.24 | 198.57 ± 45.04 | 18.05 ± 4.09 | 33.2 ± 13.76 | |
3.4. Glycosidase Activities
3.5. Neutral Sugar Composition of Larvae Food
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Martin, M.M. Cellulose digestion in insects. Comp. Biochem. Physiol. A 1983, 75, 313–324. [Google Scholar] [CrossRef]
- Watanabe, H.; Tokuda, G. Cellulolytic systems in insects. Annu. Rev. Entomol. 2010, 55, 609–632. [Google Scholar] [CrossRef]
- Terra, W.R.; Ferreira, C. Insect digestive enzymes: Properties, compartmentalization and function. Comp. Biochem. Physiol. B 1994, 109, 1–62. [Google Scholar]
- Ni, J.; Tokuda, G. Lignocellulose-degrading enzymes from termites and their symbiotic microbiota. Biotechnol. Adv. 2013, 31, 838–850. [Google Scholar] [CrossRef]
- Debris, M.M.; Chararas, C.; Courtois, J.E. Répartition des enzymes hydrolysant les polyssacharides chez quelques insectes parasites des peupliers et un xylophage du cèdre. C. R. Séances Soc. Biol. Filiales. 1964, 158, 1241–1243. [Google Scholar]
- Soo Hoo, C.F.; Dudzinski, A. Digestion by the larva of the pruinose scarab, Sericesthis geminate. Ent. Exp. Appl. 1967, 10, 7–15. [Google Scholar] [CrossRef]
- Biggs, D.R.; McGregor, P.G. Gut pH and amylase and protease activity in larvae of the New Zealand grass grub (Costelytra zealandica; Coleoptera: Scarabaeidae) as a basis for selecting inhibitors. Insect Biochem. Mol. Biol. 1996, 26, 69–75. [Google Scholar] [CrossRef]
- Mishra, S.C.; Sen-Sarma, P.K. Carbohydrases in xylophagous coleopterous larvae (Cerambycidae and Scarabaeidae) and their evolutionary significance. Mater. Org. 1985, 20, 221–230. [Google Scholar]
- Rössler, M.E. Ernährungsphysiologische Untersuchungen an Scarabaeidenlarven (Oryctes nasicornis L., Melolontha melolontha L.). J. Inst. Physiol. 1961, 6, 62–80. [Google Scholar] [CrossRef]
- Yamane, A.; Nitto, M.; Shibamoto, T. Food habit of forest insects (V). Comparison of carbohydrate-hydrolyzing enzymes from the larvae of wood-boring beetles, Japanese horned scarabaeid, and pine eggar (in Japanese). Trans. 76th Meet. Jpn. For. Soc. 1965, 1, 393–395. [Google Scholar]
- Bayon, C. Volatile fatty acids and methane production in relation to anaerobic carbohydrate fermentation in Oryctes nasicornis larvae (Coleoptera: Scarabaeidae). J. Insect Physiol. 1980, 26, 819–828. [Google Scholar] [CrossRef]
- Schlottke, E. Über die Verdauungsfermente im Holz fressender Käferlarven. Zool. Jahrb. Abt. Allg. Zool. Physiol. Tiere. 1945, 61, 88–140. [Google Scholar]
- Cazemier, A.E.; Op den Camp, H.J.M.; Hackstein, J.H.P.; Vogels, G.D. Fibre digestion in arthropods. Comp. Biochem. Physiol. Physiol. A 1997, 118, 101–109. [Google Scholar]
- Raw, F. Arthropoda (except Acari and Collembola). In Soil Biology; Burges, A., Raw, F., Eds.; Academic Press: London, UK, 1967; pp. 342–350. [Google Scholar]
- Crowson, R.A. The Biology of the Coleoptera, 1st ed.; Academic Press: London, UK, 1981. [Google Scholar]
- Ritcher, P.O. Biology of Scarabaeidae. Annu. Rev. Entomol. 1958, 3, 311–334. [Google Scholar] [CrossRef]
- Hurpin, B.; Fresneau, M. Élevage de deux dynastides, Oryctes nasicornis L., Phyllognathus silenus F (Coleopt: Scarabaeidae). Rev. Pathol. Vég. d’Entomol. Agric. Fr. 1964, 43, 75–96. [Google Scholar]
- Koyama, M.; Iwata, R.; Yamane, A. Comparison of larval growth of two scarabaeids fed with mixture of cow dung and wood chip. Jpn. J. Environ. Entomol. Zool. 2003, 14, 1–7. [Google Scholar]
- Koyama, M.; Iwata, R.; Yamane, A.; Katase, T.; Ueda, S. Nutrient intake in the third instar larvae of Anomala cuprea and Protaetia orientalis submarmorea (Coleoptera: Scarabaeidae) from a mixture of cow dung and wood chips: Results from stable isotope analyses of nitrogen and carbon. Appl. Entomol. Zool. 2003, 38, 305–311. [Google Scholar] [CrossRef]
- Green, J.M. Drying reactivity of cellulose. In Methods in Carbohydrate Chemistry, 1st ed.; Whistler, R.L., Ed.; Academic Press: New York, NY, USA, 1963; Volume 3, pp. 95–103. [Google Scholar]
- Cirelli, A.F.; de Lederkremer, R.M. A gel-forming (1→3)-β-D-glucan isolated from Cyttaria harioti Fischer. Carbohydr. Res. 1976, 48, 217–223. [Google Scholar] [CrossRef]
- Iriki, Y.; Miwa, T. Chemical nature of the cell wall of the green algae, Codium, Acetabularia and Halicoryne. Nature 1960, 185, 178–179. [Google Scholar] [CrossRef]
- Kamimiya, S.; Nishiya, T.; Izaki, K.; Takahashi, H. Purification and properties of pectin trans-eliminase in Erwinia aroideae formed in the presence of nalidixic acid. Agr. Biol. Chem. 1974, 38, 1071–1078. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for the determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar]
- Nelson, N. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem. 1944, 153, 375–380. [Google Scholar]
- Somogyi, M. Notes on sugar determination. J. Biol. Chem. 1952, 195, 19–23. [Google Scholar]
- Anzai, H.; Uchida, N.; Nishide, E. Interfering effects of the buffer solutions on the method of Somogyi and Nelson (in Japanese with English abstract). Bull. Coll. Agr. Vet. Med. Nihon Univ. 1988, 45, 23–27. [Google Scholar]
- Anzai, H.; Enami, Y.; Chida, T.; Okoshi, A.; Omuro, T.; Uchida, N.; Nishide, E. Polysaccharide digestive enzymes from midgut gland of abalone (in Japanese with English abstract). Bull. Coll. Agr. Vet. Med. Nihon Univ. 1991, 48, 119–127. [Google Scholar]
- Unno, K. Dynastine Beetles(in Japanese), 1st ed.; Data House: Tokyo, Japan, 1993; pp. 54–80. [Google Scholar]
- Borchardt, L.G.; Piper, C.V. A gas chromatographic method for carbohydrates as alditol-acetates. Tappi 1970, 53, 257–260. [Google Scholar]
- Swingle, M.C. Anatomy and physiology of the digestive tract of the Japanese beetle. J. Agric. Res. 1930, 41, 181–196. [Google Scholar]
- Staudenmayer, T.; Stellwaag, F. Über die Wasserstoffionenkonzentration und das Pufferungsvermögen des Darmes von Clysia ambiguella, Polychrosis botrana und einigen anderen Insekten sowie ihres Futters. Z. Angew. Entomol. 1940, 6, 589–607. [Google Scholar]
- Wildbolz, T. Beitrag zur Anatomie, Histologie und Physiologie des Darmkanals der Larve von Mololontha melolontha L. Mitt. Schweiz.Entomol. Ges. 1954, 27, 193–239. [Google Scholar]
- Grayson, J.M.D. Digestive tract pH of six species of Coleoptera. Ann. Entomol. Soc. Am. 1958, 51, 403–405. [Google Scholar]
- Broadway, R.M.; Villani, M.G. Does host range influence susceptibility of herbivorous insects to non-host plant proteinase inhibitors? Entomol. Exp. Appl. 1995, 76, 303–312. [Google Scholar] [CrossRef]
- McGhie, T.K.; Christeller, J.T.; Ford, R.; Allsopp, P.G. Characterization of midgut proteinase activities of white grubs: Lepidiota noxia, Lepidiota negatoria, and Antitrogus consanguineus (Scarabaeidae, Melolonthini). Arch. Insect Biochem. Physiol. 1995, 28, 351–363. [Google Scholar] [CrossRef]
- Johnson, K.S.; Rabosky, D. Phylogenetic distribution of cysteine proteinases in beetles: Evidence for an evolutionary shift to an alkaline digestive strategy in Cerambycidae. Comp. Biochem. Physiol. B 2000, 126, 609–619. [Google Scholar] [CrossRef]
- Lemke, T.; Stingl, U.; Egert, M.; Friedrich, M.W.; Brune, A. Physicochemical conditions and microbial activities in the highly alkaline gut of the humus-feeding larva of Pachonda ephippiata (Coleoptera: Scarabaeidae). Appl. Environ. Microbiol. 2003, 69, 6650–6658. [Google Scholar] [CrossRef]
- Ricou, M.G. Les diastases du tube digestif de Melolontha melolontha L. Rev. Pathol. Vég. d’Entomol. Agric. Fr. 1958, 37, 249–253. [Google Scholar]
- Yamane, A.; Nitto, M.; Shibamoto, T. (Food habit of forest insects (II). The carbohydrate-hydrolyzing enzymes from the larvae of Shirahoshizo rufescens Roelofs (in Japanese). Trans. 74th Meet. Jpn. For. Soc. 1963, 1, 345–348. [Google Scholar]
- Courtois, J.E.; Petek, F.; Kolahi-Zanouzi, M.A. Étude des osidases du hanneton. C. R. Séances Soc. Biol. Filiales. 1962, 156, 565–566. [Google Scholar]
- Wiedemann, J.F. Die Zelluloseverdauung bei Lamellicornierlarven. Z. Morph. Ökol. Tiere. 1930, 19, 228–258. [Google Scholar]
- Courtois, J.E.; Chararas, C. Les enzymes hydrolysant les glucides (hydrates de carbone) chez les insectes xylophages parasites des conifères et de quelques autres arbres forestiers. Beih. Mater. Org. 1966, 1, 127–150. [Google Scholar]
- Holter, P. Food utilization of dung-eating Aphodius larvae (Scarabaeidae). Oikos 1974, 25, 71–79. [Google Scholar] [CrossRef]
- Bauchop, T.; Clarke, R.T.J. Gut microbiology and carbohydrate digestion in the larva of Costelytra zealandica (Coleoptera: Scarabaeidae). N. Z. J. Zool. 1975, 2, 237–243. [Google Scholar] [CrossRef]
- Kawai, M. Glycanase. In Chemistry and Biochemistry of the Mushroom, 1st ed.; Mizuno, T., Kawai, M., Eds.; Gakkai Syuppan Center Co. Ltd.: Tokyo, Japan, 1992; pp. 175–181. [Google Scholar]
- Takeishi, H.; Anzai, H.; Urai, M.; Aizawa, T.; Wada, N.; Iwabuchi, N.; Sunairi, M.; Nakajima, M. Xylanolytic and alkaliphilic Dietzia sp. isolated from larvae of the Japanese horned beetle, Trypoxylus dichotomas. Actinomycetologica 2006, 20, 49–54. [Google Scholar]
- Tribolium Genome Sequencing Consortium. The genome of the model beetle and pest Tribolium castaneum. Nature 2008, 452, 949–955. [Google Scholar] [CrossRef]
- Keeling, C.I.; Yuen, M.M.; Liao, N.Y.; Roderick Docking, T.; Chan, S.K.; Taylor, G.A.; Palmquist, D.L.; Jackman, S.D.; Nguyen, A.; Li, M.; et al. Draft genome of the mountain pine beetle, Dendroctonus ponderosae Hopkins, a major forest pest. Genome Biol. 2013, 14, R27. [Google Scholar]
- Pauchet, Y.; Kirsch, R.; Giraud, S.; Vogel, H.; Heckel, D.G. Identification and characterization of plant cell wall degrading enzymes from three glycoside hydrolase families in the cerambycid beetle Apriona japonica. Insect Biochem. Mol. Biol. 2014, 49, 1–13. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wada, N.; Sunairi, M.; Anzai, H.; Iwata, R.; Yamane, A.; Nakajima, M. Glycolytic Activities in the Larval Digestive Tract of Trypoxylus dichotomus (Coleoptera: Scarabaeidae). Insects 2014, 5, 351-363. https://doi.org/10.3390/insects5020351
Wada N, Sunairi M, Anzai H, Iwata R, Yamane A, Nakajima M. Glycolytic Activities in the Larval Digestive Tract of Trypoxylus dichotomus (Coleoptera: Scarabaeidae). Insects. 2014; 5(2):351-363. https://doi.org/10.3390/insects5020351
Chicago/Turabian StyleWada, Noriko, Michio Sunairi, Hirosi Anzai, Ryûtarô Iwata, Akiomi Yamane, and Mutsuyasu Nakajima. 2014. "Glycolytic Activities in the Larval Digestive Tract of Trypoxylus dichotomus (Coleoptera: Scarabaeidae)" Insects 5, no. 2: 351-363. https://doi.org/10.3390/insects5020351