
‘Candidatus Phytoplasma ziziphi’ Changes the Metabolite Composition of Jujube Tree Leaves and Affects the Feeding Behavior of Its Insect Vector Hishimonus hamatus Kuoh


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. H. hamatus Collection and Rearing
2.2. Plant Rearing
2.3. Electropenetrography
2.4. EPG Statistic Analysis
2.5. Widely Targeted Metabolome Analysis
3. Results
3.1. Feeding Behavior of H. hamatus
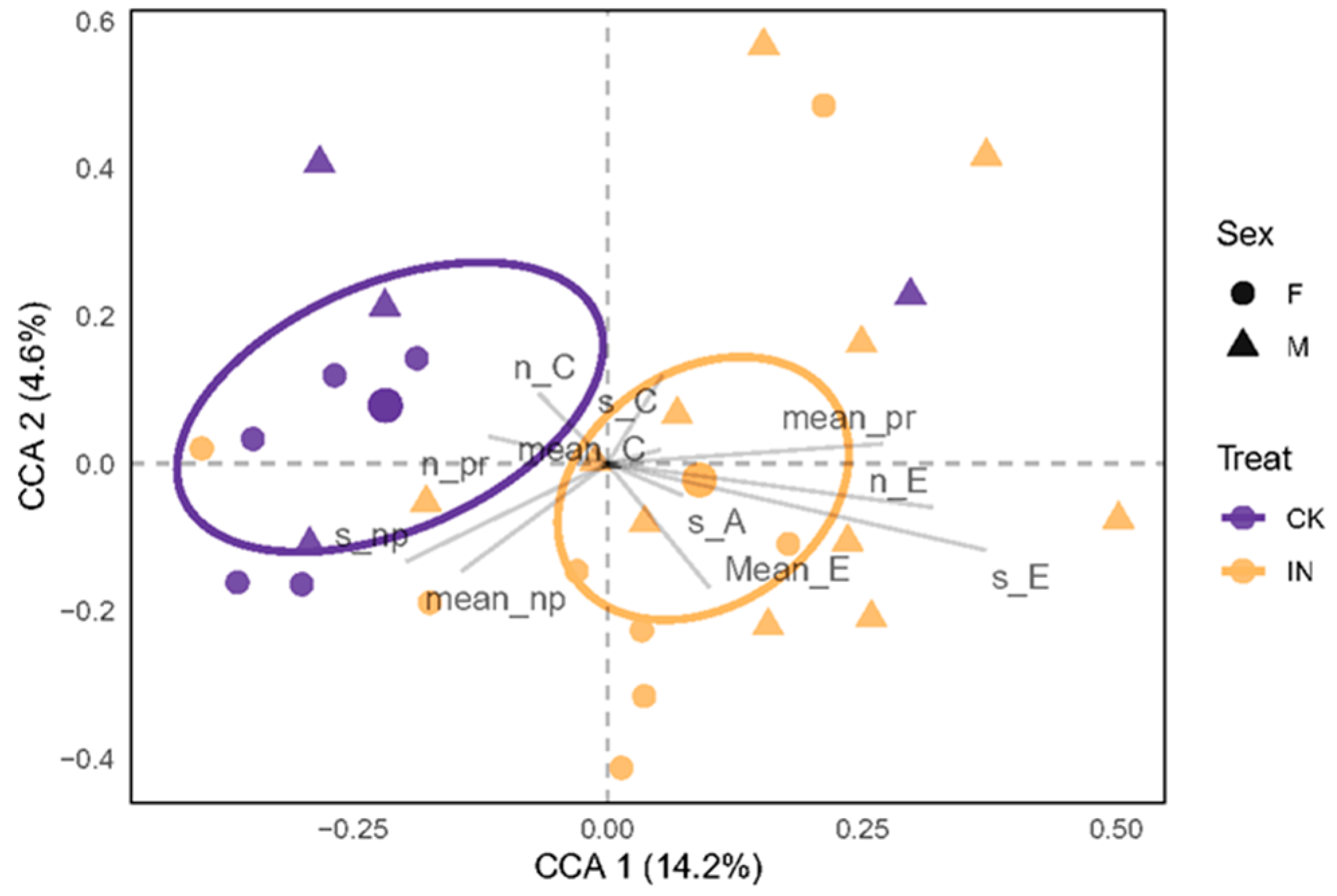
3.2. Metabolomic Analysis of Ziziphus jujuba Leaves Infected by Phytoplasma
4. Discussion
4.1. JWB Infection Affected the Feeding Behavior of Hishimonus hamatus
4.2. JWB Infection Changed the Metabolite Composition of Chinese Jujube Leaves
4.2.1. Changes of Carbohydrates
4.2.2. Changes of Lipids
4.2.3. Changes of Amino Acids
4.2.4. Changes of Lignans and Coumarins
4.2.5. Changes of Triterpenoids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pagliari, L.; Musetti, R. Phytoplasmas: An Introduction. In Phytoplasmas; Musetti, R., Pagliari, L., Eds.; Springer: New York, NY, USA, 2019; pp. 1–6. ISBN 978-1-4939-8836-5. [Google Scholar]
- Hogenhout, S.A.; Oshima, K.; Ammar, E.; Kakizawa, S.; Kingdom, H.N.; Namba, S. Phytoplasmas: Bacteria That Manipulate Plants and Insects. Mol. Plant Pathol. 2008, 9, 403–423. [Google Scholar] [CrossRef] [PubMed]
- Weintraub, P.G.; Trivellone, V.; Krüger, K. The Biology and Ecology of Leafhopper Transmission of Phytoplasmas. In Phytoplasmas: Plant Pathogenic Bacteria—II; Bertaccini, A., Weintraub, P.G., Rao, G.P., Mori, N., Eds.; Springer: Singapore, 2019; pp. 27–51. ISBN 9789811328312. [Google Scholar]
- Lee, I.M.; Davis, R.E.; Gundersen-Rindal, D.E. Phytoplasma: Phytopathogenic Mollicutes. Annu. Rev. Microbiol. 2000, 54, 221–255. [Google Scholar] [CrossRef]
- Marcone, C.; Valiunas, D.; Mondal, S.; Sundararaj, R. On Some Significant Phytoplasma Diseases of Forest Trees: An Update. Forests 2021, 12, 408. [Google Scholar] [CrossRef]
- Sugio, A.; MacLean, A.M.; Kingdom, H.N.; Grieve, V.M.; Manimekalai, R.; Hogenhout, S.A. Diverse Targets of Phytoplasma Effectors: From Plant Development to Defense Against Insects. Annu. Rev. Phytopathol. 2011, 49, 175–195. [Google Scholar] [CrossRef]
- Tomkins, M.; Kliot, A.; Marée, A.F.; Hogenhout, S.A. A Multi-Layered Mechanistic Modelling Approach to Understand How Effector Genes Extend beyond Phytoplasma to Modulate Plant Hosts, Insect Vectors and the Environment. Curr. Opin. Plant Bio. 2018, 44, 39–48. [Google Scholar] [CrossRef]
- Liu, M.; Wang, J.; Wang, L.; Liu, P.; Zhao, J.; Zhao, Z.; Yao, S.; Stănică, F.; Liu, Z.; Wang, L.; et al. The Historical and Current Research Progress on Jujube–a Superfruit for the Future. Hortic. Res. 2020, 7, 119. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Zhang, Z.; Li, S.; Lian, Q.; Fu, P.; He, Y.; Qiao, J.; Xu, K.; Liu, L.; Wu, M.; et al. Genomic Analyses of Diverse Wild and Cultivated Accessions Provide Insights into the Evolutionary History of Jujube. Plant Biotechnol. J. 2021, 19, 517–531. [Google Scholar] [CrossRef] [PubMed]
- Rashwan, A.K.; Karim, N.; Shishir, M.R.I.; Bao, T.; Lu, Y.; Chen, W. Jujube Fruit: A Potential Nutritious Fruit for the Development of Functional Food Products. J. Funct. Foods 2020, 75, 104205. [Google Scholar] [CrossRef]
- Ma, Y.; Han, Y.; Feng, X.; Gao, H.; Cao, B.; Song, L. Genome-Wide Identification of BAM (β-Amylase) Gene Family in Jujube (Ziziphus Jujuba Mill.) and Expression in Response to Abiotic Stress. BMC Genom. 2022, 23, 438. [Google Scholar] [CrossRef]
- Shi, Q.; Han, G.; Liu, Y.; Jiang, J.; Jia, Y.; Li, X. Nutrient Composition and Quality Traits of Dried Jujube Fruits in Seven Producing Areas Based on Metabolomics Analysis. Food Chem. 2022, 385, 132627. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, L.; Chen, P.; Li, Q.; Yang, Q.; Zhang, Y.; Tan, B.; Ye, X.; Zheng, X.; Feng, J. Genome-Wide Identification and Expression of the Lipoxygenase Gene Family in Jujube (Ziziphus jujuba) in Response to Phytoplasma Infection. J. Plant Biochem. Biotechnol. 2022, 31, 139–153. [Google Scholar] [CrossRef]
- Wang, J.; Song, L.; Jiao, Q.; Yang, S.; Gao, R.; Lu, X.; Zhou, G. Comparative Genome Analysis of Jujube Witches’-Broom Phytoplasma, an Obligate Pathogen That Causes Jujube Witches’-Broom Disease. BMC Genom. 2018, 19, 689. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Liu, Z.; Dai, L.; Bu, J.; Liu, M.; Zhao, Z.; Jiang, Z.; Gao, W.; Zhao, J. Changing Host Photosynthetic, Carbohydrate, and Energy Metabolisms Play Important Roles in Phytoplasma Infection. Phytopathology 2018, 108, 1067–1077. [Google Scholar] [CrossRef]
- Jung, H.Y.; Sawayanagi, T.; Kakizawa, S.; Nishigawa, H.; Wei, W.; Oshima, K.; Miyata, S.; Ugaki, M.; Hibi, T.; Namba, S. ‘Candidatus Phytoplasma Ziziphi’, a Novel Phytoplasma Taxon Associated with Jujube Witches’-Broom Disease. Int. J. Syst. Evol. Micr. 2003, 53, 1037–1041. [Google Scholar] [CrossRef]
- Deng, M.; Ma, F.; Zhang, X.; Huang, J.; Yang, J.; Chen, M.; Zhou, J.; Sun, Q.; Sun, J. Genome-Wide Identification of Jujube Witches’ Broom Phytoplasma Effectors Revealed the Role of SJP3 in Inducing Phyllody. Sci. Hortic. 2021, 290, 110548. [Google Scholar] [CrossRef]
- Zhou, J.; Ma, F.; Yao, Y.; Deng, M.; Chen, M.; Zhang, S.; Li, Y.; Yang, J.; Zhang, N.; Huang, J.; et al. Jujube Witches’ Broom Phytoplasma Effectors SJP1 and SJP2 Induce Lateral Bud Outgrowth by Repressing the ZJBRC1 -controlled Auxin Efflux Channel. Plant Cell Environ. 2021, 44, 3257–3272. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Chen, L.; Ye, X.; Tan, B.; Zheng, X.; Cheng, J.; Wang, W.; Yang, Q.; Zhang, Y.; Li, J.; et al. Phytoplasma Effector Zaofeng6 Induces Shoot Proliferation by Decreasing the Expression of ZjTCP7 in Ziziphus Jujuba. Hortic. Res. 2022, 9, uhab032. [Google Scholar] [CrossRef]
- Weintraub, P.G.; Beanland, L. Insect Vectors of Phytoplasmas. Annu. Rev. Entomol. 2006, 51, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Marcone, C. Molecular Biology and Pathogenicity of Phytoplasmas. Ann. Appl. Biol. 2014, 165, 199–221. [Google Scholar] [CrossRef]
- European Food Safety Authority Scientific Opinion on Pest Categorisation of Grapevine Flavescence Dorée. EFSA J. 2014, 12, 3851. [CrossRef]
- Ripamonti, M.; Pegoraro, M.; Rossi, M.; Bodino, N.; Beal, D.; Panero, L.; Marzachì, C.; Bosco, D. Prevalence of Flavescence Dorée Phytoplasma-Infected Scaphoideus titanus in Different Vineyard Agroecosystems of Northwestern Italy. Insects 2020, 11, 301. [Google Scholar] [CrossRef]
- Götsch, D.; Strauss, G.; Blümel, S. Laboratory Trials to Reduce Egg Hatching of the American Grapevine Leafhopper (Scaphoideus titanus) with Selected Insecticides. Bul. Insectol. 2020, 73, 53–58. [Google Scholar]
- Ding, L.; Lu, W.; Yang, Y.; Zhong, Q.; Zhou, T.; Wang, G.; Song, C.; Ma, W.; Chen, W.; Wu, Y. Immunodominant Membrane Protein (Imp) Promotes the Transmission of Wheat Blue Dwarf (WBD) Phytoplasma by Directly Interacting with α-Tubulin in Leafhoppers. Eur. J. Plant Pathol. 2022, 162, 357–367. [Google Scholar] [CrossRef]
- Wang, C.; Yu, B.W.; Zhou, P.Z.; Jiang, X.Y.; Shen, J.Y.; Chen, Z.Y. A Study of the Insect Vector (Hishimonides chinensis Aufrjv) Transmiting the Jujube Witches’ Broome Disease (I). Acta Phytopathol. Sin. 1981, 11, 27–31. [Google Scholar] [CrossRef]
- Sun, S.M.; Zhang, F.W.; Tian, X.D.; Wan, X. The Observation of the Biological Characteristics of Hishimonoides fascialis Kuoh, the Vector of Jujube Witches’ Broom Diseas. Chin. Fruits 1985, 9, 42–45. [Google Scholar] [CrossRef]
- Cai, P.; Cui, S.Y.; Kuoh, C.L. A new Species of Hishimonus injurious to Ziziphus jujuba (Homoptera: Cicadelloidea, Euscelidae). Acta Entomol. Sin. 1995, 38, 217–219. [Google Scholar] [CrossRef]
- Guo, J.L.; Wang, J.X.; He, G.X.; Qiaerban, H.; Dai, L.Z.; Huang, L.B.; Zhang, Z.Y.; Zhang, T.Q.; Ren, Z.G.; Wang, J.Z. Infection of Hishimonus lamellatus by Wolbachia (Rickettsiales: Alphaproteobacteria) and Jujube Witches’-Broom Phytoplasma, and phylogenetic analysis of these pathogens. Chin. J. Appl. Entomol. 2021, 58, 949–958. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, C.A.; Zhou, P.Z.; Liu, X.F. Investigations on the Distribution of the Insect Vector Transmitting the Jujube Witches’ Broom Disease. Acta Phytopathol. Sin. 1986, 13, 174. [Google Scholar]
- Chen, Y.X.; Zhang, Y.X.; Liu, R.C.; Li, L.L.; Li, C.Q.; Li, B.L.; Li, G.W. Investigation on Occurrence and Damage of Jujube Witches’-Broom, Jujube Fruit-Cracking and Carposina sasakii in jujube area of Northern Shaanxi. Plant Protect. 2023, 49, 317–327+345. [Google Scholar] [CrossRef]
- Backus, E.A.; Guedes, R.N.C.; Reif, K.E. AC–DC Electropenetrography: Fundamentals, Controversies, and Perspectives for Arthropod Pest Management. Pest Manag. Sci. 2021, 77, 1132–1149. [Google Scholar] [CrossRef]
- Ripamonti, M.; Maron, F.; Cornara, D.; Marzachì, C.; Fereres, A.; Bosco, D. Leafhopper Feeding Behaviour on Three Grapevine Cultivars with Different Susceptibilities to Flavescence Dorée. J. Insect Physiol. 2022, 137, 104366. [Google Scholar] [CrossRef] [PubMed]
- Ripamonti, M.; Galetto, L.; Maron, F.; Marzachì, C.; Bosco, D. Scaphoideus titanus Fitness on Grapevine Varieties with Different Susceptibility to Flavescence Dorée Phytoplasma. J. Appl. Entomol. 2022, 146, 1260–1271. [Google Scholar] [CrossRef]
- Görg, L.M.; Gross, J. Influence of Ontogenetic and Migration Stage on Feeding Behavior of Cacopsylla picta on ‘Candidatus Phytoplasma Mali’ Infected and Non-Infected Apple Plants. J. Insect Physiol. 2021, 131, 104229. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. 2022. Available online: https://github.com/vegandevs/vegan (accessed on 11 October 2022).
- Quensen, J. Ggordiplots: Make Ggplot Versions of Vegans Ordiplots. 2023. Available online: https://github.com/jfq3/ggordiplots (accessed on 23 July 2023).
- Naimi, B.; Hamm, N.; Groen, T.A.; Skidmore, A.K.; Toxopeus, A.G. Where is positional uncertainty a problem for species distribution modelling. Ecography 2014, 37, 191–203. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; Balakrishnan, N., Colton, T., Everitt, B., Piegorsch, W., Ruggeri, F., Teugels, J.L., Eds.; Wiley: Hoboken, NJ, USA, 2017; pp. 1–15. ISBN 978-1-118-44511-2. [Google Scholar]
- Sandanayaka, W.R.M.; Jia, Y.; Charles, J.G. EPG Technique as a Tool to Reveal Host Plant Acceptance by Xylem Sap-Feeding Insects. J. Appl. Entomol. 2013, 137, 519–529. [Google Scholar] [CrossRef]
- Backus, E.A.; Cervantes, F.A.; Guedes, R.N.C.; Li, A.Y.; Wayadande, A.C. AC–DC Electropenetrography for In-Depth Studies of Feeding and Oviposition Behaviors. Ann. Entomol. Soc. Am. 2019, 112, 236–248. [Google Scholar] [CrossRef]
- Roddee, J.; Backus, E.A.; Wangkeeree, J.; Hanboonsong, Y. Alteration in the Stylet Probing Behavior and Host Preference of the Vector Matsumuratettix hiroglyphicus (Hemiptera: Cicadellidae) After Infection with Sugarcane White Leaf Phytoplasma. J. Econ. Entomol. 2021, 114, 1081–1090. [Google Scholar] [CrossRef]
- Mauck, K.; Bosque-Pérez, N.A.; Eigenbrode, S.D.; De Moraes, C.M.; Mescher, M.C. Transmission Mechanisms Shape Pathogen Effects on Host-Vector Interactions: Evidence from Plant Viruses. Funct. Ecol. 2012, 26, 1162–1175. [Google Scholar] [CrossRef]
- Kube, M.; Schneider, B.; Kuhl, H.; Dandekar, T.; Heitmann, K.; Migdoll, A.M.; Reinhardt, R.; Seemüller, E. The Linear Chromosome of the Plant-Pathogenic Mycoplasma ‘Candidatus Phytoplasma Mali’. BMC Genom. 2008, 9, 306. [Google Scholar] [CrossRef]
- Tan, Y.; Li, Q.L.; Zhao, Y.; Wei, H.; Wang, J.; Baker, C.J.; Liu, Q.; Wei, W. Integration of Metabolomics and Existing Omics Data Reveals a New Insights into Phytoplasma-Induced Metabolic Reprogramming in Host Plants. PLoS ONE 2021, 16, e0246203. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal Triacylglycerols as Feedstocks for Biofuel Production: Perspectives and Advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef] [PubMed]
- Oshima, K.; Maejima, K.; Namba, S. Genomic and Evolutionary Aspects of Phytoplasmas. Front. Microbiol. 2013, 4, 230. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Cho, S.T.; Lin, Y.C.; Tan, C.M.; Chiu, Y.C.; Yang, J.Y.; Kuo, C.H. Comparative Genome Analysis of ‘Candidatus Phytoplasma luffae’ Reveals the Influential Roles of Potential Mobile Units in Phytoplasma Evolution. Front. Microbiol. 2022, 13, 773608. [Google Scholar] [CrossRef] [PubMed]
- Fischer, H.D.; Goggin, F.L. Impact of loss-of-function of Fatty Acid Desaturase 7 in Arabidopsis thaliana on the green peach aphid, Myzus persicae, and the cabbage aphid, Brevicoryne brassicae. Entomol. Exp. Appl. 2022, 170, 235–244. [Google Scholar] [CrossRef]
- Hildebrandt, T.M.; Nunes Nesi, A.; Araújo, W.L.; Braun, H.P. Amino Acid Catabolism in Plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef] [PubMed]
- Brodbeck, B.V.; Mizell, R.F.; French, W.J.; Andersen, P.C.; Aldrich, J.H. Amino Acids as Determinants of Host Preference for the Xylem Feeding Leafhopper, Homalodisca coagulata (Homoptera: Cicadellidae). Oecologia 1990, 83, 338–345. [Google Scholar] [CrossRef]
- Görg, L.M.; Gallinger, J.; Gross, J. The Phytopathogen ‘Candidatus Phytoplasma Mali’ Alters Apple Tree Phloem Composition and Affects Oviposition Behavior of Its Vector Cacopsylla picta. Chemoecology 2021, 31, 31–45. [Google Scholar] [CrossRef]
- Umezawa, T. Diversity in lignan biosynthesis. Phytochem. Rev. 2003, 2, 371–390. [Google Scholar] [CrossRef]
- Harmatha, J.; Nawrot, J. Insect feeding deterrent activity of lignans and related phenylpropanoids with a methylenedioxyphenyl (piperonyl) structure moiety. In Proceedings of the 11th International Symposium on Insect-Plant Relationships; Nielsen, J.K., Kjaer, C., Schoonhoven, L.M., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 51–60. [Google Scholar] [CrossRef]
- Harmatha, J.; Dinan, L. Biological activities of lignans and stilbenoids associated with plant-insect chemical interactions. Phytochem. Rev. 2003, 2, 321–330. [Google Scholar] [CrossRef]
- Saguez, J.; Attoumbré, J.; Giordanengo, P.; Baltora-Rosset, S. Biological activities of lignans and neolignans on the aphid Myzus persicae (Sulzer). Arthropod-Plant Inte. 2013, 7, 225–233. [Google Scholar] [CrossRef]
- Li, Y.K.; Wei, J.Q.; Fang, J.M.; Lv, W.B.; Ji, Y.F.; Aioub, A.A.A.; Zhang, J.W.; Hu, Z.N. Insecticidal activity of four lignans isolated from Phryma leptostachya. Molecules 2019, 24, 1976. [Google Scholar] [CrossRef]
- Pavela, R.; Maggi, F.; Benelli, G. Coumarin (2H-1-benzopyran-2-one): A novel and eco-friendly aphicide. Nat. Prod. Res. 2021, 35, 1566–1571. [Google Scholar] [CrossRef] [PubMed]
- Thimmappa, R.; Geisler, K.; Louveau, T.; O’Maille, P.; Osbourn, A. Triterpene biosynthesis in plants. Ann. Rev. Plant Biol. 2014, 65, 225–257. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.H.; Yang, S.F.; Huang, J.G.; Zhou, L.J. Insecticidal triterpenes in Meliaceae: Plant species, molecules and activities: Part I (Aphanamixis-Chukrasia). Int. J. Mol. Sci. 2021, 22, 13262. [Google Scholar] [CrossRef]
- Sanjayan, K.P.; Partho, P.D. Influence of asiatic acid, a triterpenoid from Shores robusta, on the feeding and enzyme activity of Oxya fuscovittata (Insecta, Orthoptera). J. Appl. Entomol. 1993, 115, 506–510. [Google Scholar] [CrossRef]
- Fernandes, C.P.; Xavier, A.; Pacheco, J.P.F.; Santos, M.G.; Mexas, R.; Ratcliffe, N.A.; Gonzalez, M.S.; Mello, C.B.; Rocha, L.; Feder, D. Laboratory evaluation of the effects of Manilkara subsericea (Mart.) Dubard extracts and triterpenes on the development of Dysdercus peruvianus and Oncopeltus fasciatus. Pest Manag. Sci. 2013, 69, 292–301. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treat | Healthy | JWB-Infected | ||
|---|---|---|---|---|
| Sex | Female | Male | Female | Male |
| Number of insects | 11 | 12 | 12 | 16 |
| Number of non-probing periods * | 154.0 ± 17.8 a | 130.0 ± 34.0 a | 137.5 ± 29.4 a | 123.0 ± 37.5 a |
| Total duration of non-probing periods [min] ** | 225.3 ± 21.1 a | 266.8 ± 27.6 a | 218.0 ± 28.8 ab | 113.1 ± 16.8 b |
| Number of probes * | 153.0 ± 17.8 a | 129.5 ± 34.1 a | 137.0 ± 29.4 a | 123.0 ± 37.5 a |
| Total probing time [min] * | 134.7 ± 21.1 b | 92.7 ± 27.7 b | 142.0 ± 28.8 a | 245.8 ± 16.8 a |
| Total duration of pathway phase [min] ** | 51.4 ± 18.8 b | 22.3 ± 6.5 b | 70.9 ± 16.6 b | 119.1 ± 19.2 a |
| Number of active xylem ingestion phases * | 106.0 ± 27.2 a | 88.5 ± 39.5 a | 93.5 ± 21.1 a | 100.5 ± 20.7 a |
| Total duration of active xylem ingestion [min]* | 76.0 ± 8.4 a | 52.5 ± 25.8 a | 73.0 ± 24.2 a | 118.0 ± 27.8 a |
| Mean duration of a single event of pathway phase [min] * | 0.8 ± 0.2 a | 0.9 ± 0.1 a | 0.8 ± 0.2 a | 1.1 ± 0.3 a |
| Number of phloem ingestions * | 0 ± 0.72 b | 0 ± 5.62 b | 8.5 ± 7.8 a | 17.5 ± 12.6 a |
| Total duration of phloem ingestions [min] * | 0 ± 0.12 b | 0 ± 3.78 b | 7.8 ± 6.7 a | 3.70 ± 8.67 a |
| Mean duration of a single event of phloem ingestion [min] * | 0.1 ± 0.1 a | 0.4 ± 0.1 a | 0.8 ± 0.6 a | 0.8 ± 0.3 a |
| Percentage of non-probing time [%] # | 52.0 ± 10.1 a | 25.4 ± 10.0 b | 47.3 ± 8.9 a | 20.7 ± 3.9 b |
| Percentage of probing time spent in pathway-phase [%] # | 34.7 ± 5.6 a | 60.4 ± 10.8 b | 30.7 ± 6.9 a | 49.5 ± 6.3 b |
| Percentage of probing time spent in active xylem ingestion [%] * | 13.4 ± 4.7 a | 13.8 ± 3.4 a | 16.0 ± 3.3 a | 20.4 ± 3.8 a |
| Percentage of probing time spent in passive phloem ingestion [%] * | 0.1 ± 0.1 b | 0.5 ± 2.7 b | 4.1 ± 1.7 a | 5.3 ± 2.1 a |
| Time to 1st probe [min] * | 42.0 ± 32.1 a | 65.7 ± 13.5 a | 67.6 ± 32.1 a | 5.0 ± 19.5 a |
| Time to 1st phloem ingestion [min] * | 38.7 ± 40.2 a | 92.3 ± 69.9 a | 16.9 ± 7.3 a | 10.0 ± 32.6 a |
| Number of probing times before 1st phloem ingestion * | 9.0 ± 45.8 a | 3.0 ± 85.8 a | 3.5 ± 3.8 b | 0.0 ± 0.1 b |
| Df | Sum of Sqs | R2 | F | Pr (>F) | Signif | |
|---|---|---|---|---|---|---|
| Treat | 1 | 0.1954 | 0.1243 | 3.8094 | 0.0099 | ** |
| Sex | 1 | 0.1313 | 0.0836 | 2.5608 | 0.0560 | |
| Treat × Sex | 1 | 0.0136 | 0.0087 | 0.2654 | 0.8751 | |
| Residual | 24 | 1.2308 | 0.7834 | NA | NA | |
| Total | 27 | 1.5711 | 1 | NA | NA |
| Main Class | Number of Metabolites | ||
|---|---|---|---|
| Identified | Down-Regulated | Up-Regulated | |
| Alkaloids | 82 | 13 | 34 |
| Alkaloids | 30 | 2 | 9 |
| Aporphine alkaloids | 6 | 0 | 3 |
| Benzylphenylethylamine | 1 | 0 | 1 |
| Phenolamine | 26 | 6 | 15 |
| Piperidine alkaloids | 1 | 0 | 1 |
| Plumerane alkaloids | 13 | 2 | 7 |
| Pyridine alkaloids | 2 | 1 | 0 |
| Quinoline alkaloids | 2 | 1 | 0 |
| Tropan alkaloids | 1 | 1 | 0 |
| Amino acids and derivatives | 95 | 7 | 51 |
| Flavonoids | 266 | 50 | 74 |
| Anthocyanidins | 10 | 2 | 4 |
| Chalcones | 15 | 2 | 8 |
| Flavanols | 15 | 4 | 1 |
| Flavanones | 29 | 7 | 11 |
| Flavanonols | 8 | 3 | 3 |
| Flavones | 68 | 13 | 20 |
| Flavonols | 114 | 18 | 25 |
| Isoflavones | 6 | 1 | 2 |
| Lignans and Coumarins | 43 | 27 | 4 |
| Coumarins | 19 | 11 | 1 |
| Lignans | 24 | 16 | 3 |
| Lipids | 139 | 5 | 80 |
| Free fatty acids | 64 | 1 | 17 |
| Glycerol ester | 19 | 2 | 11 |
| Lysophosphatidyl choline | 29 | 0 | 28 |
| Lysophosphatidyl ethanolamine | 23 | 2 | 21 |
| Phosphatidyl choline | 1 | 0 | 1 |
| Spshingolipids | 3 | 0 | 2 |
| Nucleotides and derivatives | 52 | 4 | 28 |
| Organic acids | 64 | 5 | 24 |
| Others | 27 | 10 | 5 |
| Aldehyde compounds | 2 | 1 | 0 |
| Chromone | 4 | 1 | 0 |
| Ketone compounds | 6 | 2 | 3 |
| Others | 15 | 6 | 2 |
| Saccharides | 66 | 3 | 12 |
| Vitamines and derivatives | 19 | 7 | 2 |
| Quinones | 8 | 1 | 1 |
| Tannins | 8 | 0 | 1 |
| Phenolic acids | 183 | 41 | 64 |
| Terpenoids | 128 | 80 | 13 |
| Triterpene | 96 | 78 | 3 |
| Triterpene Saponin | 32 | 2 | 10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, R.-C.; Li, B.-L.; Chen, X.-L.; Liu, J.-J.; Luo, K.; Li, G.-W. ‘Candidatus Phytoplasma ziziphi’ Changes the Metabolite Composition of Jujube Tree Leaves and Affects the Feeding Behavior of Its Insect Vector Hishimonus hamatus Kuoh. Insects 2023, 14, 750. https://doi.org/10.3390/insects14090750
Liu R-C, Li B-L, Chen X-L, Liu J-J, Luo K, Li G-W. ‘Candidatus Phytoplasma ziziphi’ Changes the Metabolite Composition of Jujube Tree Leaves and Affects the Feeding Behavior of Its Insect Vector Hishimonus hamatus Kuoh. Insects. 2023; 14(9):750. https://doi.org/10.3390/insects14090750
Chicago/Turabian StyleLiu, Rui-Chang, Bo-Liao Li, Xiu-Lin Chen, Jing-Jing Liu, Kun Luo, and Guang-Wei Li. 2023. "‘Candidatus Phytoplasma ziziphi’ Changes the Metabolite Composition of Jujube Tree Leaves and Affects the Feeding Behavior of Its Insect Vector Hishimonus hamatus Kuoh" Insects 14, no. 9: 750. https://doi.org/10.3390/insects14090750