
Study of Pathogenesis Using Fluorescent Strain of Cordyceps farinosa Revealed Infection of Thitarodes armoricanus Larvae via Digestive Tract


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
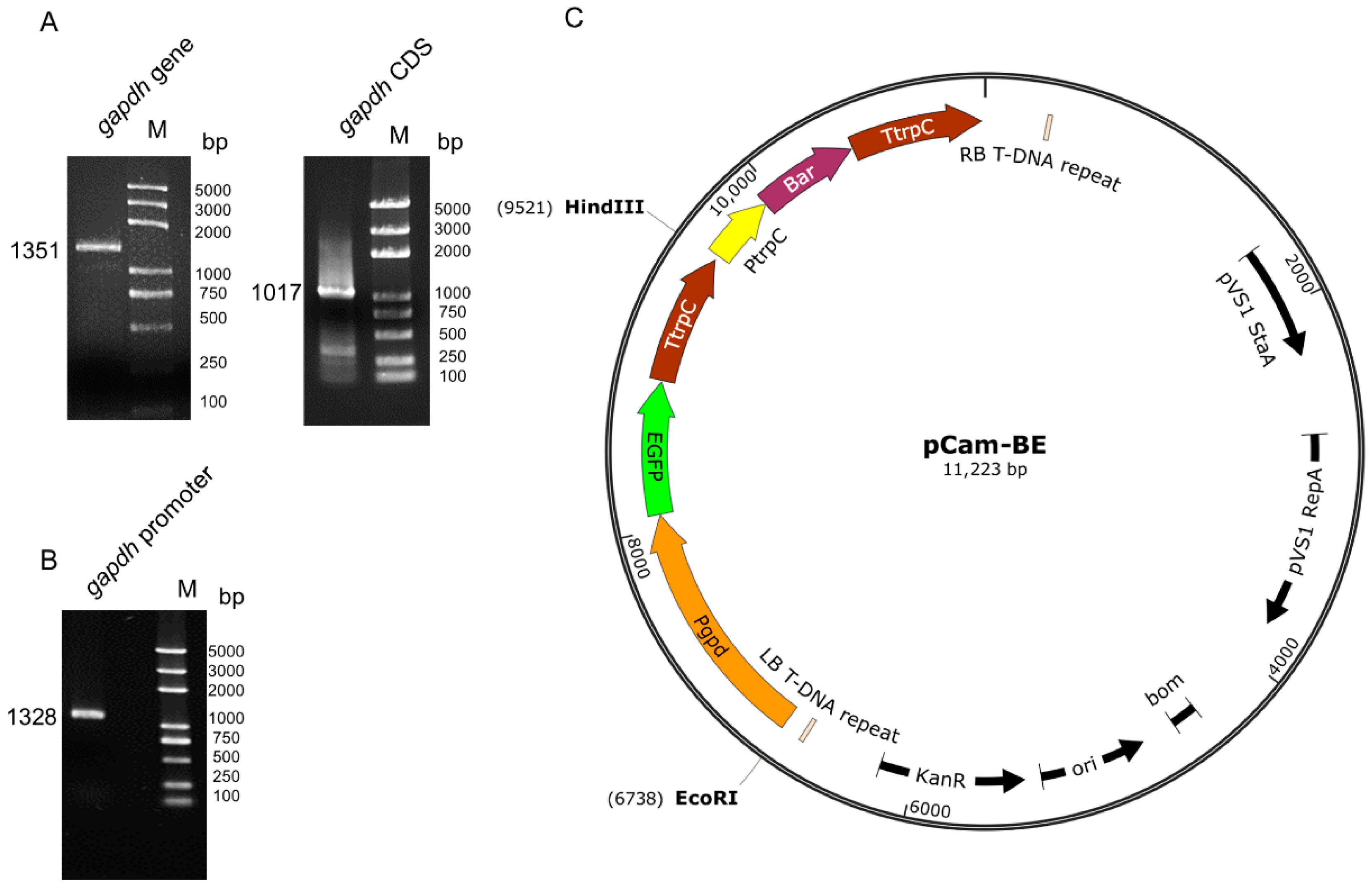
2.2. Cloning of the Glyceraldehyde 3-Phosphate Dehydrogenase (Gapdh) Gene and Promoter of C. farinosa
2.3. Construction of Binary Vectors
2.4. Agrobacterium-Mediated Transformation of C. farinosa
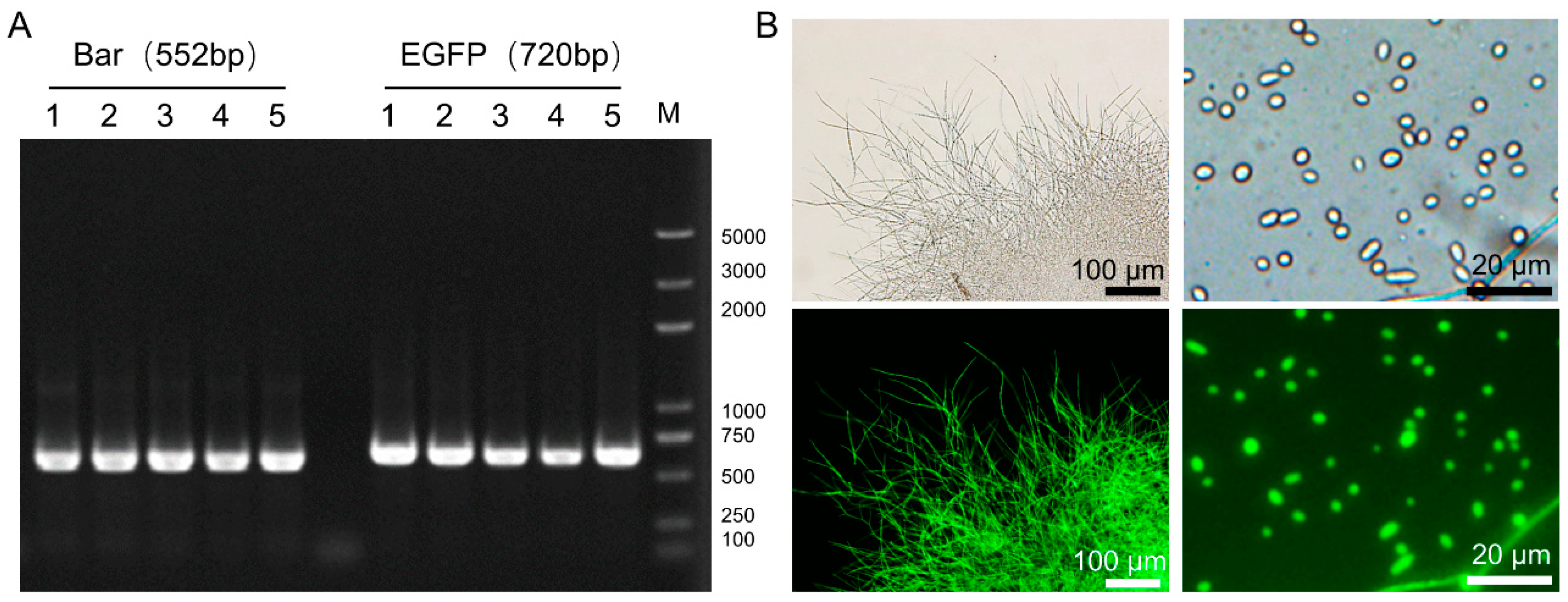
2.5. Genetic Stability and Fluorescence Analysis of the Transformants
2.6. Inoculation of T. armoricanus Larvae with C. farinosa
2.7. Collection and Observation of Infected Larvae Samples
2.8. Statistical Analysis
3. Results
3.1. pCam-BE Vector Construction
3.2. Fluorescent Strain of C. farinosa
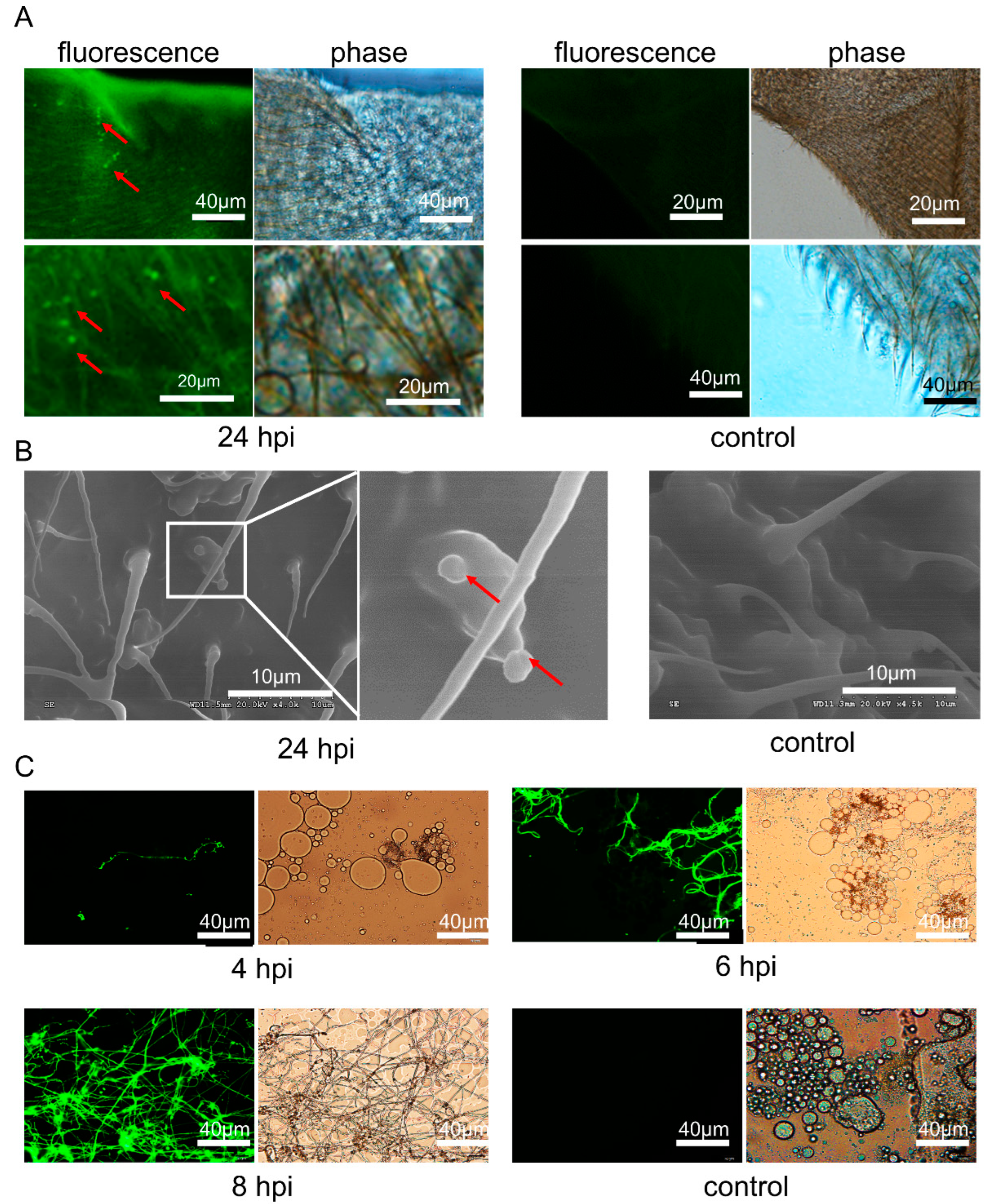
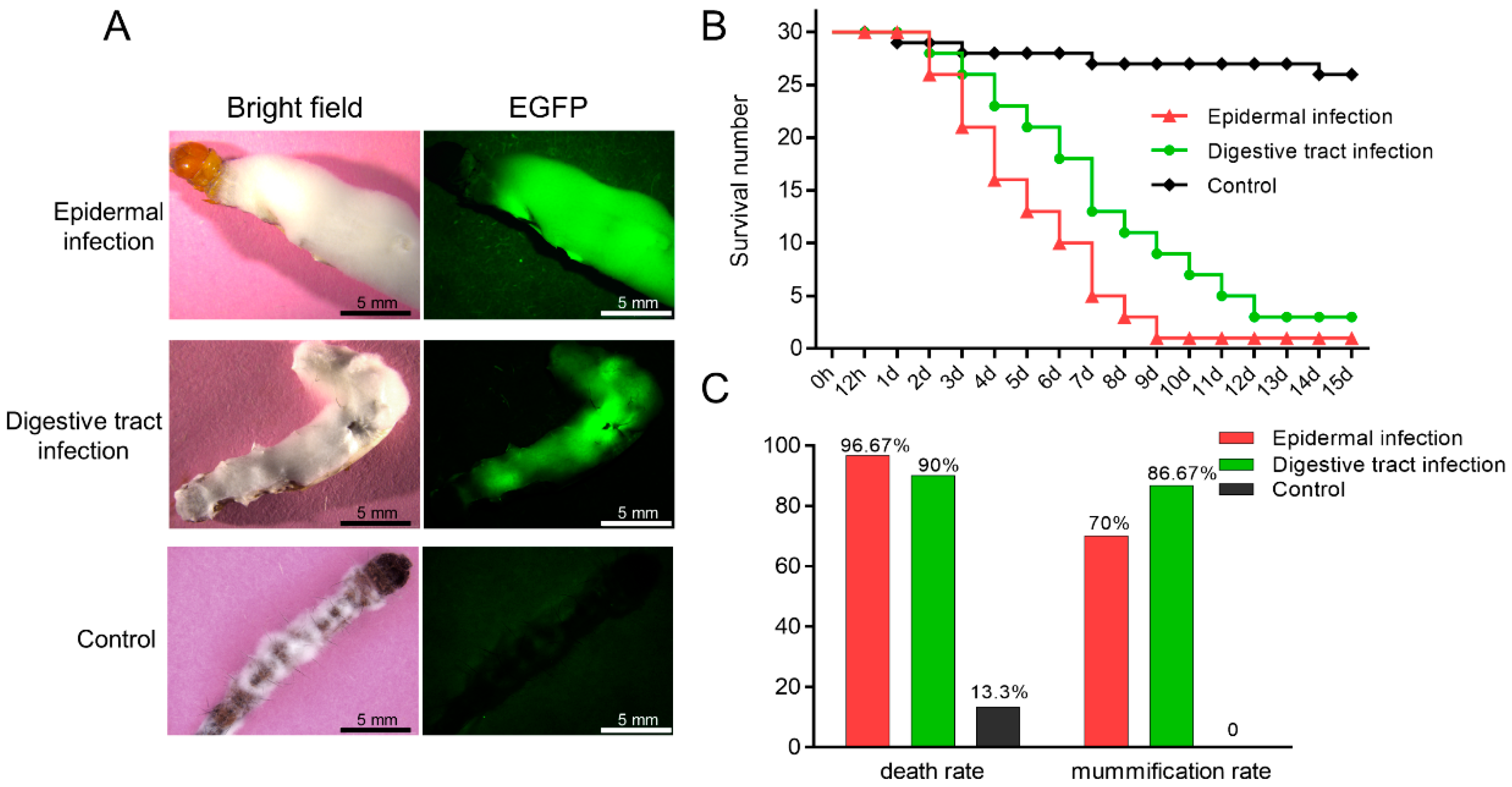
3.3. C. farinosa Infection through Host Body Surface
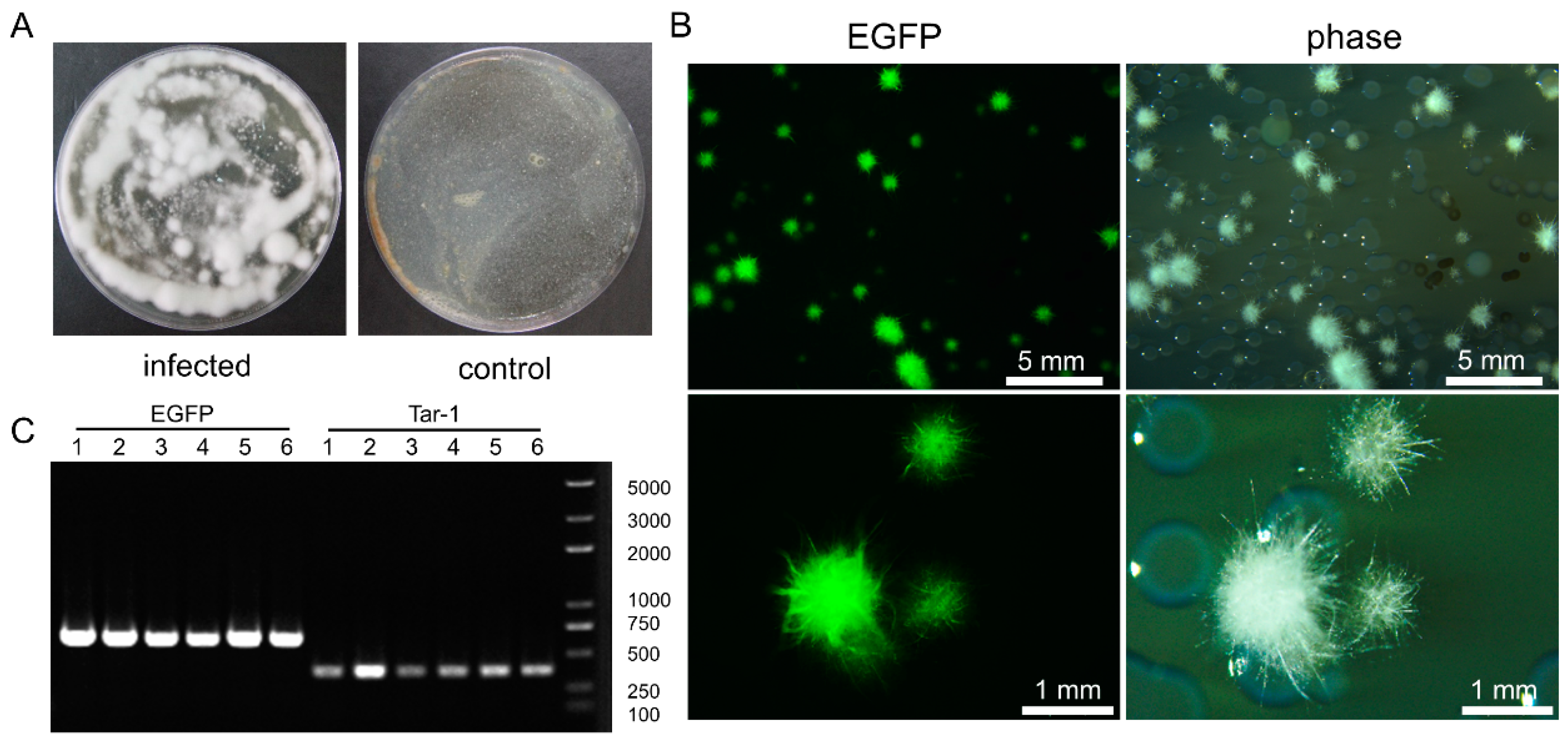
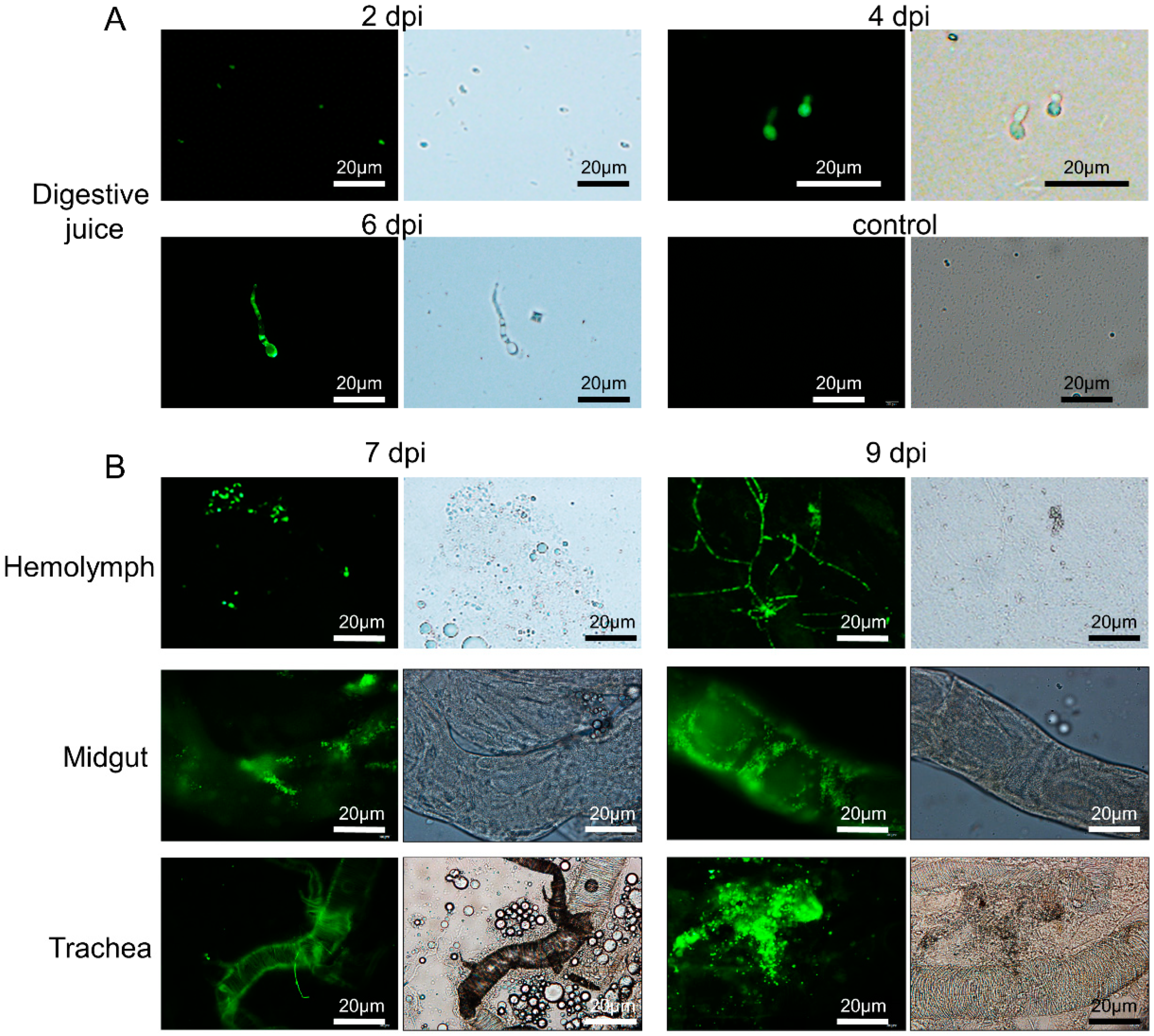
3.4. C. farinosa Infection via Host Digestive Tract
3.5. Comparison and Statistical Analysis of the Two Infection Methods
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hsu, T.H.; Shiao, L.H.; Hsieh, C.Y.; Chang, D.M. A Comparison of the Chemical Composition and Bioactive Ingredients of the Chinese Medicinal Mushroom DongChongXiaCao, Its Counterfeit and Mimic, and Fermented Mycelium of Cordyceps Sinensis. Food Chem. 2002, 78, 463–469. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.; Guo, Q.; Zheng, Q.; Zhang, W. Metabolomic Comparison between Wild Ophiocordyceps Sinensis and Artificial Cultured Cordyceps militaris. Biomed. Chromatogr. 2018, 32, e4279. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, L.; Tian, E. Advances in Research of the Artificial Cultivation of Ophiocordyceps Sinensis in China. Crit. Rev. Biotechnol. 2014, 34, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Yue, K.; Ye, M.; Lin, X.; Zhou, Z. The Artificial Cultivation of Medicinal Caterpillar Fungus, Ophiocordyceps Sinensis (Ascomycetes): A Review. Int. J. Med. Mushrooms 2013, 15, 425–434. [Google Scholar] [CrossRef]
- Dong, C.; Guo, S.; Wang, W.; Liu, X. Cordyceps Industry in China. Mycology 2015, 6, 121–129. [Google Scholar] [CrossRef]
- Liu, F.; Wu, X.; Liu, Y.; Chen, D.; Zhang, D.; Yang, D. Progress on molecular biology of Isaria farinosa, pathogen of host of Ophiocordyceps sinensis during the artificial culture. China J. Chin. Mater. Medica 2016, 41, 403–409. [Google Scholar] [CrossRef]
- He, Y.; You, H.; Lu, Z.; Chen, S. Dynamic Changes of Hepialus larva’s Entomopathogenic Fungus Isaria farinose During Culture of Ophiocordyceps sinensis. Southwest China J. Agric. Sci. 2020, 33, 2840–2845. [Google Scholar] [CrossRef]
- Ishii, M.; Kanuka, H.; Badolo, A.; Sagnon, N.; Guelbeogo, W.M.; Koike, M.; Aiuchi, D. Proboscis Infection Route of Beauveria Bassiana Triggers Early Death of Anopheles mosquito. Sci. Rep. 2017, 7, 3476. [Google Scholar] [CrossRef]
- Alkhaibari, A.M.; Carolino, A.T.; Yavasoglu, S.I.; Maffeis, T.; Mattoso, T.C.; Bull, J.C.; Samuels, R.I.; Butt, T.M. Metarhizium Brunneum Blastospore Pathogenesis in Aedes Aegypti Larvae: Attack on Several Fronts Accelerates Mortality. PLoS Pathog. 2016, 12, e1005715. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Q.; Xie, Y.; Zhu, Y.; Xue, J.; Li, J.; Fan, R. Morphological and Ultrastructural Characterization of Carposina Sasakii Larvae (Lepidoptera: Carposinidae) Infected by Beauveria Bassiana (Ascomycota: Hypocreales: Clavicipitaceae). Micron Oxf. Engl. 2013, 44, 303–311. [Google Scholar] [CrossRef]
- He, M.; Su, J.; Xu, Y.; Chen, J.; Chern, M.; Lei, M.; Qi, T.; Wang, Z.; Ryder, L.S.; Tang, B.; et al. Discovery of Broad-Spectrum Fungicides That Block Septin-Dependent Infection Processes of Pathogenic Fungi. Nat. Microbiol. 2020, 5, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Shapiro-Ilan, D.I.; Cottrell, T.E.; Jackson, M.A.; Wood, B.W. Virulence of Hypocreales Fungi to Pecan Aphids (Hemiptera: Aphididae) in the Laboratory. J. Invertebr. Pathol. 2008, 99, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Barelli, L.; Padilla-Guerrero, I.E.; Bidochka, M.J. Differential Expression of Insect and Plant Specific Adhesin Genes, Mad1 and Mad2, in Metarhizium robertsii. Fungal Biol. 2011, 115, 1174–1185. [Google Scholar] [CrossRef] [PubMed]
- Tawidian, P.; Rhodes, V.L.; Michel, K. Mosquito-Fungus Interactions and Antifungal Immunity. Insect Biochem. Mol. Biol. 2019, 111, 103182. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Shang, Y.; Tang, G.; Wang, C. Identification of a Key G-Protein Coupled Receptor in Mediating Appressorium Formation and Fungal Virulence against Insects. Sci. China Life Sci. 2021, 64, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Cheung, P.Y.K.; Grula, E.A. In Vivo Events Associated with Entomopathology of Beauveria Bassiana for the Corn Earworm (Heliothis Zea). J. Invertebr. Pathol. 1982, 39, 303–313. [Google Scholar] [CrossRef]
- Holder, D.J.; Keyhani, N.O. Adhesion of the Entomopathogenic Fungus Beauveria (Cordyceps) Bassiana to Substrata. Appl. Environ. Microbiol. 2005, 71, 5260–5266. [Google Scholar] [CrossRef] [Green Version]
- Niermans, K.; Meyer, A.M.; den Hil, E.F.H.; van Loon, J.J.A.; van der Fels-Klerx, H.J. A Systematic Literature Review on the Effects of Mycotoxin Exposure on Insects and on Mycotoxin Accumulation and Biotransformation. Mycotoxin Res. 2021, 37, 279–295. [Google Scholar] [CrossRef]
- Adachi, H.; Doi, H.; Kasahara, Y.; Sawa, R.; Nakajima, K.; Kubota, Y.; Hosokawa, N.; Tateishi, K.; Nomoto, A. Asteltoxins from the Entomopathogenic Fungus Pochonia Bulbillosa 8-H-28. J. Nat. Prod. 2015, 78, 1730–1734. [Google Scholar] [CrossRef]
- Cao, W.; Wang, G.; Zheng, W.; Wang, R.; Du, L.; Song, J.; Wang, J.; Feng, S. Comparison of toxicity of Beauveria bassiana and histopathological changes of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) larvae infected in different inoculation ways. Acta Entomol. Sin. 2011, 54, 409–415. [Google Scholar] [CrossRef]
- Lyu, Y.; Xia, J.; Zhang, Z.; Li, Q.; Dong, C.; Li, W. Symptoms, infection and histopathology of Hepialus sp. larvae parasitized by Isaria farinosa. Mycosystema 2018, 37, 314–324. [Google Scholar] [CrossRef]
- Li, W.; Dong, C.; Liu, X.; Li, Q.; Xia, J.; Liang, L. Research Advances in Artificial Cultivation of Chinese Cordyceps. Mycosystema 2016, 35, 375–387. [Google Scholar] [CrossRef]
- Steentjes, M.B.F.; Tonn, S.; Coolman, H.; Langebeeke, S.; Scholten, O.E.; van Kan, J.A.L. Visualization of Three Sclerotiniaceae Species Pathogenic on Onion Reveals Distinct Biology and Infection Strategies. Int. J. Mol. Sci. 2021, 22, 1865. [Google Scholar] [CrossRef]
- Lozoya-Pérez, N.E.; Casas-Flores, S.; Martínez-Álvarez, J.A.; López-Ramírez, L.A.; Lopes-Bezerra, L.M.; Franco, B.; Mora-Montes, H.M. Generation of Sporothrix Schenckii Mutants Expressing the Green Fluorescent Protein Suitable for the Study of Host-Fungus Interactions. Fungal Biol. 2018, 122, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.S.; Ploetz, R.C.; Rollins, J.A. Comparing Avocado, Swamp Bay, and Camphortree as Hosts of Raffaelea Lauricola Using a Green Fluorescent Protein (GFP)-Labeled Strain of the Pathogen. Phytopathology 2017, 107, 70–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooykaas, P.J.J.; van Heusden, G.P.H.; Niu, X.; Reza Roushan, M.; Soltani, J.; Zhang, X.; van der Zaal, B.J. Agrobacterium-Mediated Transformation of Yeast and Fungi. Curr. Top. Microbiol. Immunol. 2018, 418, 349–374. [Google Scholar] [CrossRef]
- Li, D.; Tang, Y.; Lin, J.; Cai, W. Methods for Genetic Transformation of Filamentous Fungi. Microb. Cell Factories 2017, 16, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Ma, Y.; Liu, Y.; Tong, S.; Zhu, S.; Jin, D.; Pei, Y.; Fan, Y. A Polyketide Synthase, BbpksP, Contributes to Conidial Cell Wall Structure and UV Tolerance in Beauveria bassiana. J. Invertebr. Pathol. 2020, 169, 107280. [Google Scholar] [CrossRef]
- Wu, P.; Qin, Q.; Zhang, J.; Zhang, H.; Li, X.; Wang, H.; Meng, Q. The Invasion Process of the Entomopathogenic Fungus Ophiocordyceps Sinensis into the Larvae of Ghost Moths (Thitarodes Xiaojinensis) Using a GFP-Labeled Strain. Front. Microbiol. 2022, 13, 974323. [Google Scholar] [CrossRef]
- Wamiti, L.G.; Khamis, F.M.; Abd-Alla, A.M.M.; Ombura, F.L.O.; Akutse, K.S.; Subramanian, S.; Odiwuor, S.O.; Ochieng, S.J.; Ekesi, S.; Maniania, N.K. Metarhizium Anisopliae Infection Reduces Trypanosoma Congolense Reproduction in Glossina Fuscipes Fuscipes and Its Ability to Acquire or Transmit the Parasite. BMC Microbiol. 2018, 18, 142. [Google Scholar] [CrossRef]
- Mauchline, N.; Hallett, I.; Hill, G.; Casonato, S. Process of Infection of Armored Scale Insects (Diaspididae) by an Entomopathogenic Cosmospora Sp. J. Invertebr. Pathol. 2011, 108, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Islam, W.; Adnan, M.; Shabbir, A.; Naveed, H.; Abubakar, Y.S.; Qasim, M.; Tayyab, M.; Noman, A.; Nisar, M.S.; Khan, K.A.; et al. Insect-Fungal-Interactions: A Detailed Review on Entomopathogenic Fungi Pathogenicity to Combat Insect Pests. Microb. Pathog. 2021, 159, 105122. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.-L.; Hou, J.; Feng, M.-G.; Ying, S.-H. Transcriptomic Analyses Reveal Comprehensive Responses of Insect Hemocytes to Mycopathogen Beauveria Bassiana, and Fungal Virulence-Related Cell Wall Protein Assists Pathogen to Evade Host Cellular Defense. Virulence 2020, 11, 1352–1365. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Cao, L.; Zhang, Y.; Ye, Y.; Han, R. Laboratory Rearing of Thitarodes Armoricanus and Thitarodes Jianchuanensis (Lepidoptera: Hepialidae), Hosts of the Chinese Medicinal Fungus Ophiocordyceps Sinensis (Hypocreales: Ophiocordycipitaceae). J. Econ. Entomol. 2016, 109, 176–181. [Google Scholar] [CrossRef]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the Surface: Entomopathogenic Fungi versus the Insect Cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Deng, J.; Wang, H.; Zhao, X.; Luo, Z.; Yu, C.; Zhang, Y. Multifunctional Role of a Fungal Pathogen-Secreted Laccase 2 in Evasion of Insect Immune Defense. Environ. Microbiol. 2021, 23, 1256–1274. [Google Scholar] [CrossRef]
- Agrawal, Y.; Khatri, I.; Subramanian, S.; Shenoy, B.D. Genome Sequence, Comparative Analysis, and Evolutionary Insights into Chitinases of Entomopathogenic Fungus Hirsutella thompsonii. Genome Biol. Evol. 2015, 7, 916–930. [Google Scholar] [CrossRef]
- Feng, P.; Shang, Y.; Cen, K.; Wang, C. Fungal Biosynthesis of the Bibenzoquinone Oosporein to Evade Insect Immunity. Proc. Natl. Acad. Sci. USA 2015, 112, 11365–11370. [Google Scholar] [CrossRef] [Green Version]
- Yin, F.; Xiao, M.; Berestetskiy, A.; Hu, Q. The Metarhizium Anisopliae Toxin, Destruxin A, Interacts with the SEC23A and TEME214 Proteins of Bombyx mori. J. Fungi 2021, 7, 460. [Google Scholar] [CrossRef]
- Sharififard, M.; Mossadegh, M.; Vazirianzadeh, B.; Zarei-Mahmoudabadi, A. Interactions between Entomopathogenic Fungus, Metarhizium Anisopliae and Sublethal Doses of Spinosad for Control of House Fly, Musca domestica. Iran. J. Arthropod-Borne Dis. 2011, 5, 28–36. [Google Scholar]
- Zimmermann, G. The Entomopathogenic Fungi Isaria Farinosa (Formerly Paecilomyces Farinosus) and the Isaria Fumosorosea Species Complex (Formerly Paecilomyces Fumosoroseus): Biology, Ecology and Use in Biological Control. Biocontrol Sci. Technol. 2008, 18, 865–901. [Google Scholar] [CrossRef]
- Liu, F.; Wu, X.; He, Z.Y.; Xiang, M.; He, Y.; Liu, X.; Chen, S.; Zeng, W. Nutritional Effects on the Mycelial Growth and Enzymatic Activity of Isaria Farinosa, and Hepialus Larvae Growth. J. Appl. Microbiol. 2016, 120, 1605–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, Q.; Zhang, X.; Chen, W.; Hu, Q. Secondary Metabolites and the Risks of Isaria Fumosorosea and Isaria farinosa. Molecules 2019, 24, 664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, Y.; Hu, J.; Wang, F.; Chen, R.; Chen, S.; Li, Y.; Qin, S. Biological characteristics and miticide activity of Paecilomyces farinosus to Panonychus citri. J. Hunan Agric. Univ. Sci. 2017, 43, 411–416. [Google Scholar] [CrossRef]
- Li, X.; Liu, Q.; Li, W.; Li, Q.; Qian, Z.; Liu, X.; Dong, C. A Breakthrough in the Artificial Cultivation of Chinese Cordyceps on a Large-Scale and Its Impact on Science, the Economy, and Industry. Crit. Rev. Biotechnol. 2019, 39, 181–191. [Google Scholar] [CrossRef]
- Cao, L.; Ye, Y.; Han, R. Fruiting Body Production of the Medicinal Chinese Caterpillar Mushroom, Ophiocordyceps Sinensis (Ascomycetes), in Artificial Medium. Int. J. Med. Mushrooms 2015, 17, 1107–1112. [Google Scholar] [CrossRef]
- He, Z.; Zhang, D.; Li, L.; Xing, K.; Chen, S. The infection research on the Ophiocordyceps sinensis filamentous mycelium to Hepialus moths larvae. J. Environ. Entomol. 2017, 39, 673–678. [Google Scholar]
- Lei, W.; Zhang, G.; Peng, Q.; Liu, X. Development of Ophiocordyceps Sinensis through Plant-Mediated Interkingdom Host Colonization. Int. J. Mol. Sci. 2015, 16, 17482–17493. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, C.; Wei, J.; Pan, G.; Li, C.; Zhou, Z. Study of Pathogenesis Using Fluorescent Strain of Cordyceps farinosa Revealed Infection of Thitarodes armoricanus Larvae via Digestive Tract. Insects 2022, 13, 1039. https://doi.org/10.3390/insects13111039
Tong C, Wei J, Pan G, Li C, Zhou Z. Study of Pathogenesis Using Fluorescent Strain of Cordyceps farinosa Revealed Infection of Thitarodes armoricanus Larvae via Digestive Tract. Insects. 2022; 13(11):1039. https://doi.org/10.3390/insects13111039
Chicago/Turabian StyleTong, Chaoqun, Junhong Wei, Guoqing Pan, Chunfeng Li, and Zeyang Zhou. 2022. "Study of Pathogenesis Using Fluorescent Strain of Cordyceps farinosa Revealed Infection of Thitarodes armoricanus Larvae via Digestive Tract" Insects 13, no. 11: 1039. https://doi.org/10.3390/insects13111039