The Effects of Alternative Foods on Life History and Cannibalism of Amblyseius herbicolus (Acari: Phytoseiidae)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Materials
2.2. Experimental Apparatus
2.3. Effects of Different Alternative Foods on Life Table Parameters
2.4. Effects of Different Alternative Foods on Cannibalism
2.5. Statistical Analyses
3. Results
3.1. Developmental Durations and Reproduction
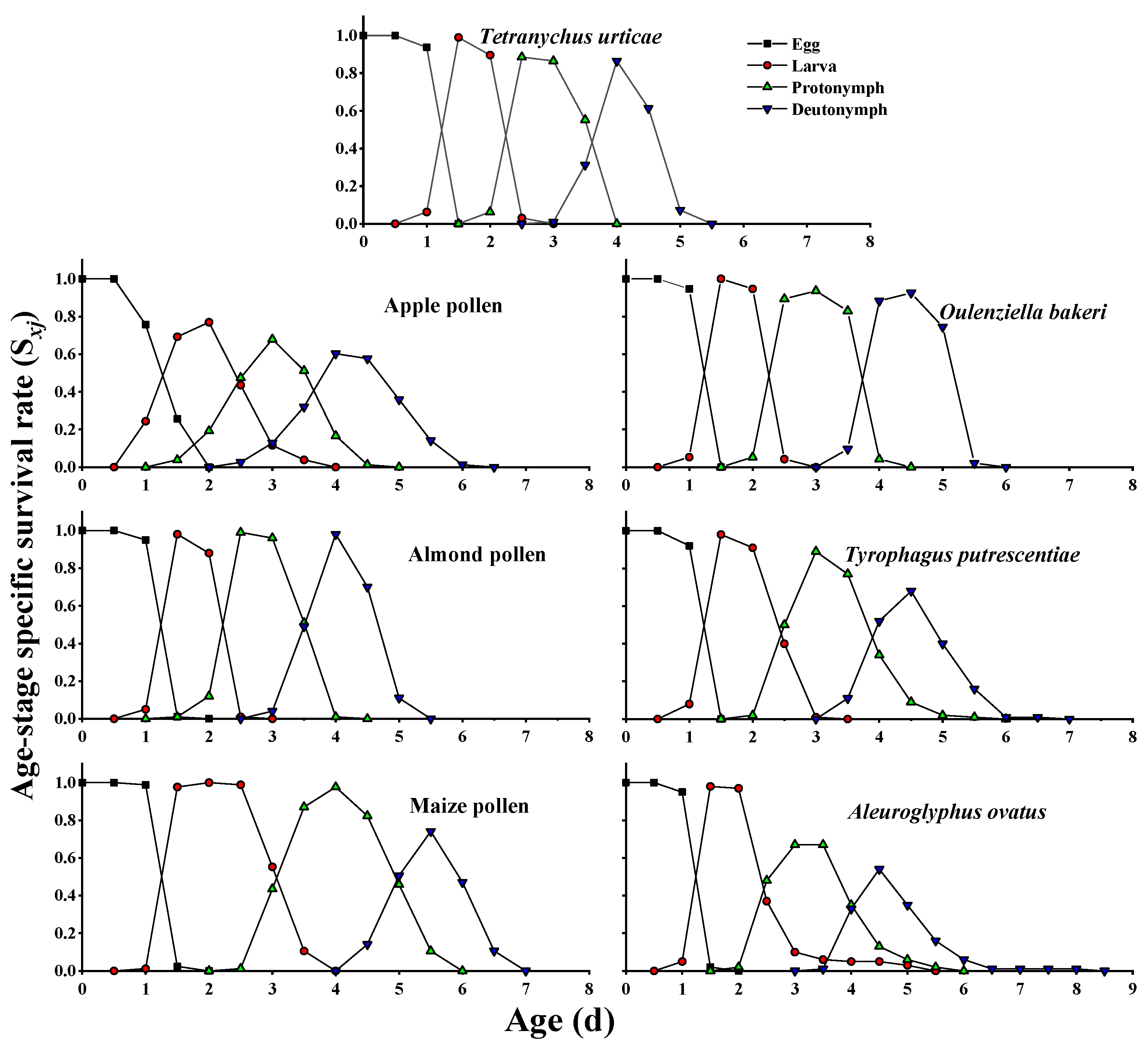
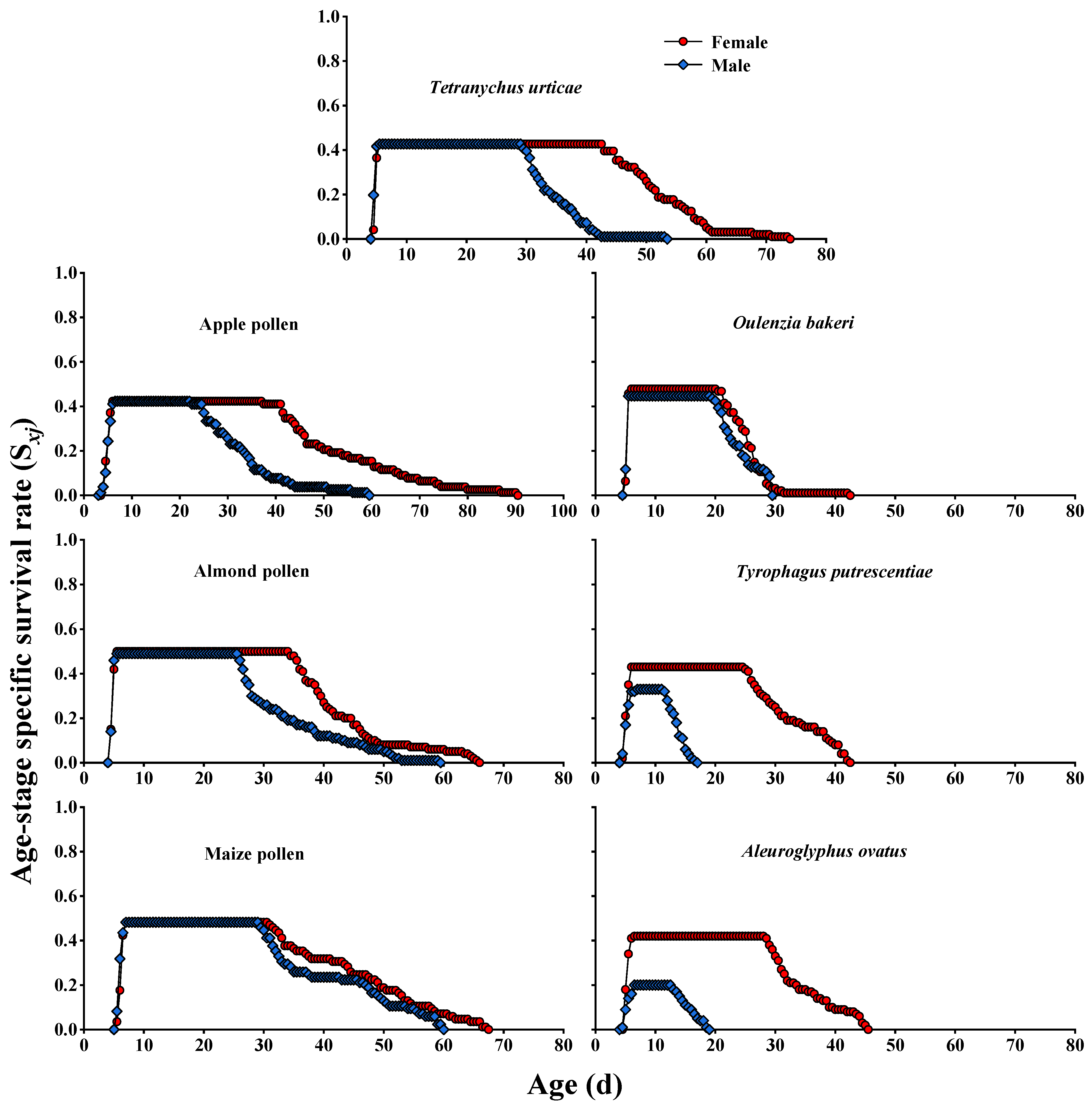
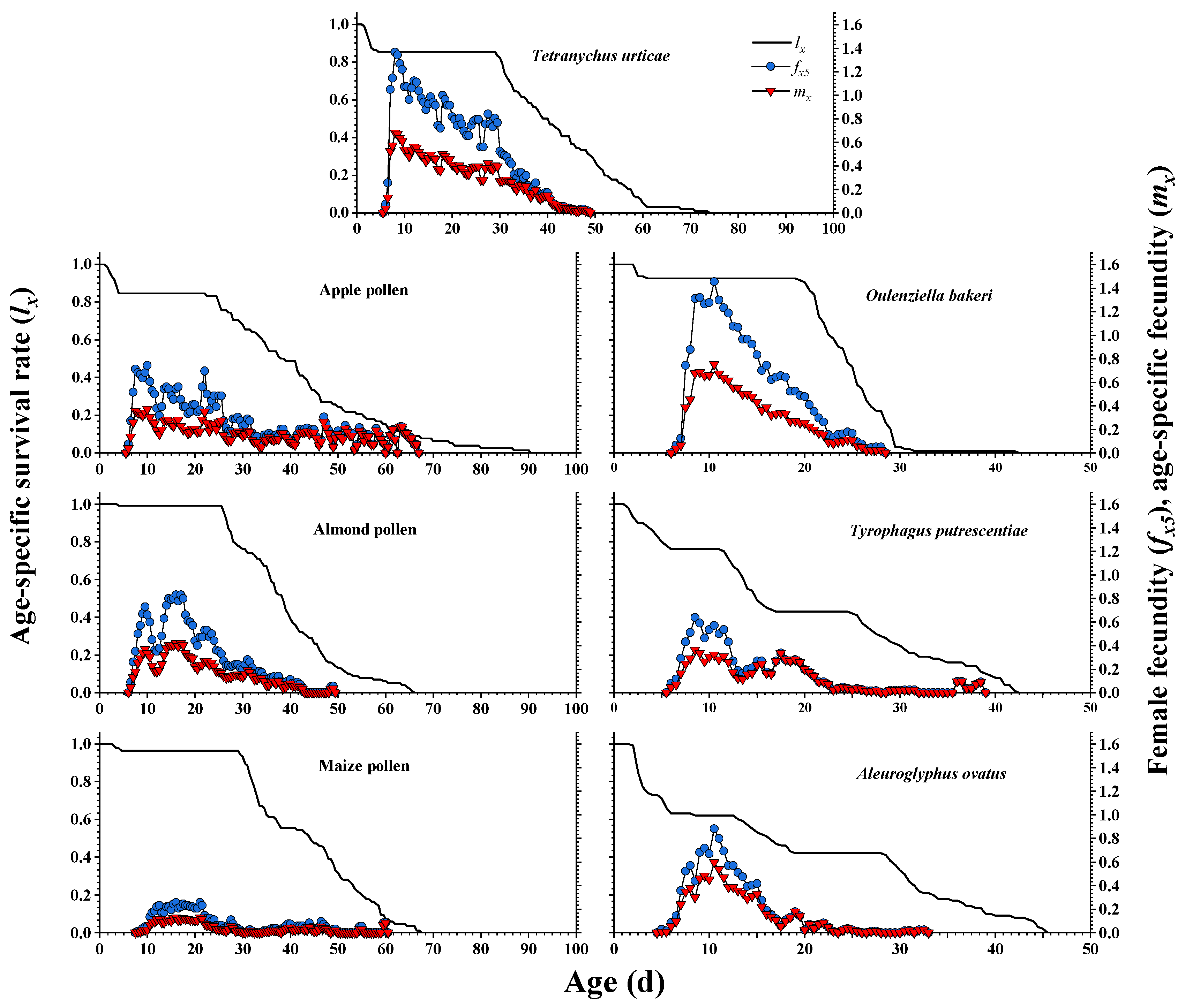
3.2. Survival Rate and Fecundity Curves
3.3. Population Parameters
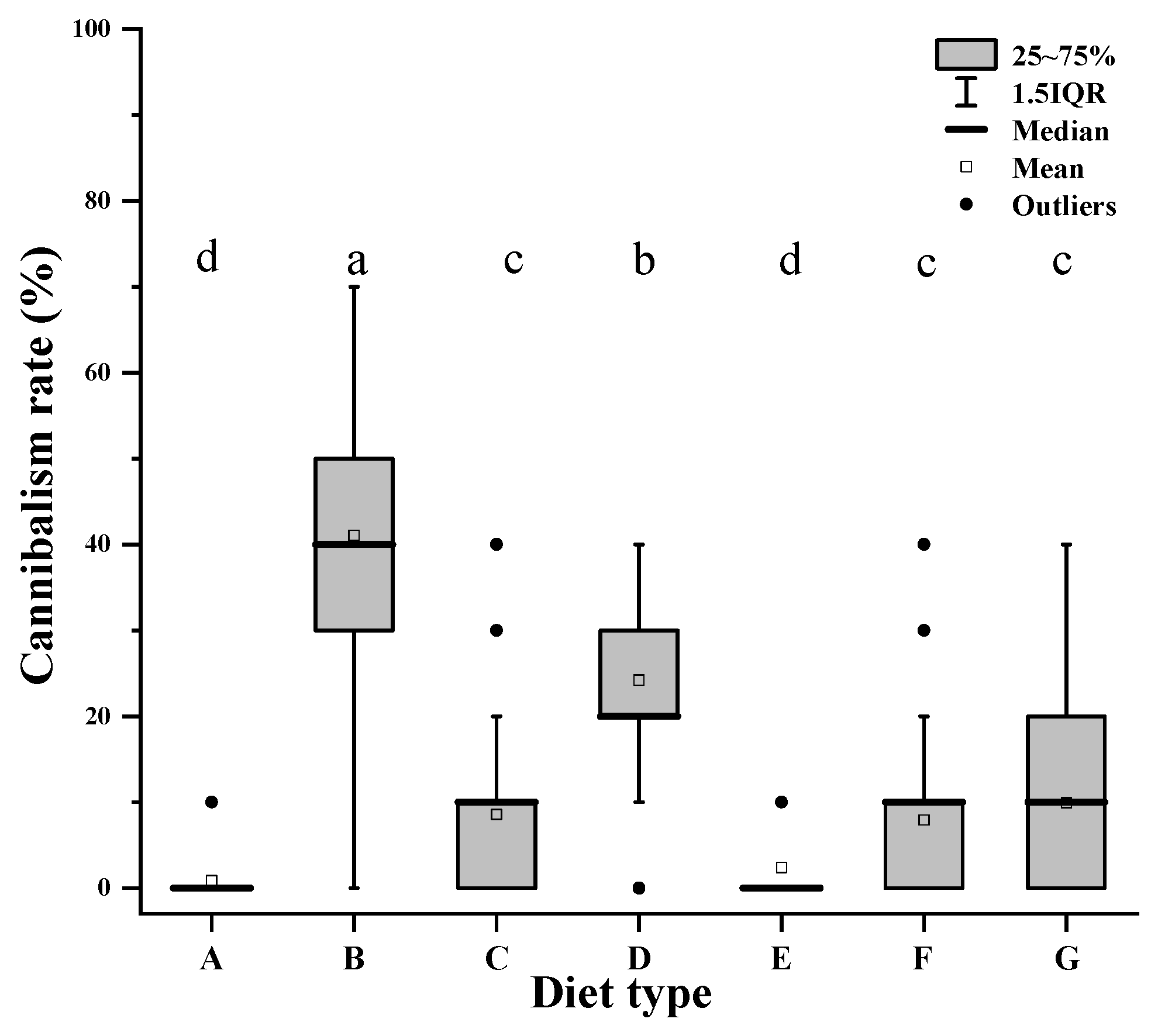
3.4. Effects of Different Alternative Foods on Cannibalism
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Lenteren, J.C. The state of commercial augmentative biological control: Plenty of natural enemies, but a frustrating lack of uptake. BioControl 2012, 57, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Bernardo, A.M.G.; De Oliveira, C.M.; Oliveira, R.A.; Vacacela, H.E.; Venzon, M.; Pallini, A.; Janssen, A. Performance of Orius insidiosus on alternative foods. J. Appl. Entomol. 2017, 141, 702–707. [Google Scholar] [CrossRef]
- Knapp, M.; Van Houten, Y.; Van Baal, E.; Groot, T. Use of predatory mites in commercial biocontrol: Current status and future prospects. Acarologia 2018, 58, 72–82. [Google Scholar] [CrossRef]
- Portillo, N.; Alomar, O.; Wäckers, F. Nectarivory by the planttissue feeding predator Macrolophus pygmaeus Rambur (Heteroptera: Miridae): Nutritional redundancy or nutritional beneft? J. Insect Physiol. 2012, 58, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Pumariño, L.; Alomar, O. The role of omnivory in the conservation of predators: Orius majusculus (Heteroptera: Anthocoridae) on sweet alyssum. Biol. Control 2012, 62, 24–28. [Google Scholar] [CrossRef]
- Marcossi, T.; Fonseca, M.M.; Carbajal, P.; Cardoso, A.; Janssen, A. High–quality alternative food reduces cannibalism in the predatory mite Amblyseius herbicolus (Acari: Phytoseiidae). Exp. Appl. Acarol. 2020, 81, 189–200. [Google Scholar] [CrossRef]
- Vandekerkhove, B.; De Clercq, P. Pollen as an alternative or supplementary food for the mirid predator Macrolophus pygmaeus. Biol. Control 2010, 53, 238–242. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Wu, H.; Pang, H.; De Clercq, P. An artificial diet containing plant pollen for the mealybug predator Cryptolaemus montrouzieri. Pest Manag. Sci. 2017, 73, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Owashi, Y.; Hayashi, M.; Abe, J.; Miura, K. Effects of an alternative diet of Artemia cysts on the development and reproduction of Nesidiocoris tenuis (Hemiptera: Miridae). Appl. Entomol. Zool. 2019, 55, 121–127. [Google Scholar] [CrossRef]
- Vangansbeke, D.; Nguyen, D.T.; Audenaert, J.; Verhoeven, R.; Deforce, K.; Gobin, B.; Tirry, L.; De Clercq, P. Diet–dependent cannibalism in the omnivorous phytoseiid mite Amblydromalus limonicus. Biol. Control 2014, 74, 30–35. [Google Scholar] [CrossRef]
- Cavalcante, A.C.C.; Borges, L.R.; Lourenção, A.L.; De Moraes, G.J. Potential of two populations of Amblyseius swirskii (Acari: Phytoseiidae) for the control of Bemisia tabaci biotype B (Hemiptera: Aleyrodidae) in Brazil. Exp. Appl. Acarol. 2015, 67, 523–533. [Google Scholar] [CrossRef]
- Bouagga, S.; Urbaneja, A.; Pérez–Hedo, M. Combined use of predatory mirids with Amblyseius swirskii (Acari: Phytoseiidae) to enhance pest management in sweet pepper. J. Econ. Entomol. 2018, 111, 1112–1120. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, M.; Saqib, S.M. Rearing of predatory seven spotted ladybird beetle Coccinella septempunctata L. (Coleoptera: Coccinellidae) on natural and artificial diets under laboratory conditions. Pak. J. Zool. 2010, 42, 47–51. [Google Scholar]
- Ricupero, M.; Dai, C.; Siscaro, G.; Russo, A.; Biondi, A.; Zappalà, L. Potential diet regimens for laboratory rearing of the harlequin ladybird. BioControl 2020, 65, 583–592. [Google Scholar] [CrossRef]
- Hauge, M.S.; Nielsen, F.H.; Toft, S. Weak responses to dietary enrichment in a specialized aphid predator. Physiol. Entomol. 2011, 36, 360–367. [Google Scholar] [CrossRef]
- Sun, Y.X.; Hao, Y.N.; Riddick, E.W.; Liu, T.X. Factitious prey and artificial diets for predatory lady beetles: Current situation, obstacles, and approaches for improvement: A review. Biocontrol Sci. Technol. 2017, 27, 601–619. [Google Scholar] [CrossRef]
- Chen, M.L.; Huang, Y.H.; Qiu, B.Y.; Chen, P.T.; Du, X.Y.; Li, H.S.; Pang, H. Changes in life history traits and transcriptional regulation of Coccinellini ladybirds in using alternative prey. BMC Genom. 2020, 21, 44. [Google Scholar] [CrossRef]
- Schausberger, P. Cannibalism among phytoseiid mites: A review. Exp. Appl. Acarol. 2003, 29, 173–191. [Google Scholar] [CrossRef] [PubMed]
- Farazmand, A.; Fathipour, Y.; Kamali, K. Cannibalism in Scolothrips longicornis (Thysanoptera: Thripidae), Neoseiulus californicus and Typhlodromus bagdasarjani (Acari: Phytoseiidae). Syst. Appl. Acarol. 2014, 19, 471–480. [Google Scholar]
- Liu, M.X.; Chu, W.Q.; Xu, C.; Zheng, Q.M.; Song, W.B.; Li, Y.Y.; Liu, H. Extraguild prey availability reduced cannibalism and reciprocal intraguild predation of Neoseiulus barkeri (Acari: Phytoseiidae) and Scolothrips takahashii (Thysanoptera: Thripidae). Syst. Appl. Acarol. 2020, 25, 775–786. [Google Scholar]
- Calabuig, A.; Pekas, A.; Wäckers, F.L. The quality of nonprey food affects cannibalism, intraguild predation, and hyperpredation in two species of phytoseiid mites. J. Econ. Entomol. 2018, 111, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.T.; Vangansbeke, D.; Clercq, P. Artificial and factitious foods support the development and reproduction of the predatory mite Amblyseius swirskii. Exp. Appl. Acarol. 2013, 62, 181–194. [Google Scholar] [CrossRef]
- Duarte, M.V.A.; Venzon, M.; Bittencourt, M.C.S.; Rodríguez-Cruz, F.A.; Pallini, A.; Janssen, A. Alternative food promotes broad mite control on Chilli pepper plants. BioControl 2015, 60, 817–825. [Google Scholar] [CrossRef]
- Samaras, K.; Pappas, M.L.; Fytas, E.; Broufas, G.D. Pollen suitability for the development and reproduction of Amblydromalus limonicus (Acari: Phytoseiidae). BioControl 2015, 60, 773–782. [Google Scholar] [CrossRef]
- Kalile, M.O.; Cardoso, A.C.; Pallini, A.; Fonseca, M.M.; Elliot, S.L.; Fialho, V.S.; Carvalho, T.S.; Janssen, A. A predatory mite as potential biological control agent of Diaphorina citri. BioControl 2021, 66, 237–248. [Google Scholar] [CrossRef]
- Hajizadeh, J.; Nazari, M. A checklist and key for the phytoseiid mites (Acari: Phytoseiidae) of citrus orchards in Iran, with a new record for Iranian phytoseiid mites. Syst. Appl. Acarol. 2012, 17, 388–396. [Google Scholar] [CrossRef]
- Rodríguez-Cruz, F.A.; Janssen, A.; Pallini, A.; Duarte, M.V.A.; Pinto, C.M.F.; Venzon, M. Two predatory mite species as potential control agents of broad mites. BioControl 2017, 62, 505–513. [Google Scholar] [CrossRef]
- Berton, L.H.C.; De Carvalho Mineiro, J.L.; Sato, M.E.; Joaquim, A.D.A.F.; Raga, A. Mite fauna of a coffee agroecosystem (Coffea arabica L.) in the municipality of Monte Alegre do Sul, São Paulo State, Brazil. Part I. Acarologia 2019, 59, 542–550. [Google Scholar] [CrossRef]
- Franco, R.A.; Reis, P.R.; Zacarias, M.S.; Oliveira, D.C. Influência da teia de Oligonychus ilicis (McGregor) (Acari: Tetranychidae) sobre os fitoseídeos predadores associados. Neotrop. Entomol. 2010, 39, 97–100. [Google Scholar] [CrossRef] [Green Version]
- Reis, P.R.; Teodoro, A.V.; Pedro Neto, M.; Silva, E.A. Life history of Amblyseius herbicolus (Chant) (Acari: Phytoseiidae) on coffee plants. Neotrop. Entomol. 2007, 36, 282–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, W.; Paynter, Q.; Zhang, Z.Q. Predation, prey preference and reproduction of predatory mites Amblydromalus limonicus (Garman), Amblyseius herbicolus (Chant) and Neoseiulus cucumeris (Oudemans) (Mesostigmata: Phytoseiidae) on immature Sericothrips staphylinus Haliday (Thysanoptera: Thripidae), a biocontrol agent of gorse. Syst. Appl. Acarol. 2019, 24, 508–519. [Google Scholar]
- Lam, W.; Paynter, Q.; Zhang, Z.Q. Functional response of Amblyseius herbicolus (Acari: Phytoseiidae) on Sericothrips staphylinus (Thysanoptera: Thripidae), an ineffective biocontrol agent of gorse. Biol. Control 2021, 152, 104468. [Google Scholar] [CrossRef]
- Cavalcante, A.C.C.; Dos Santos, V.L.V.; Rossi, L.C.; Moraes, G.J.D. Potential of five Brazilian populations of Phytoseiidae (Acari) for the biological control of Bemisia tabaci (Insecta: Hemiptera). J. Econ. Entomol. 2015, 108, 29–33. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Z.Q. The dried fruit mite Carpoglyphus lactis (Acari: Carpoglyphidae) is a suitable alternative prey for Amblyseius herbicolus (Acari: Phytoseiidae). Syst. Appl. Acarol. 2021, 26, 2167–2176. [Google Scholar] [CrossRef]
- Chi, H.S.I.N.; Liu, H.S.I. Two new methods for the study of insect population ecology. Boll. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-able analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, T. MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. 2021. Available online: http://140.120.197.173/Ecology/prod02.htm (accessed on 8 May 2021).
- Palevsky, E. Pollen provisioning for the promotion of biological control by omnivorous phytoseiids in organic greenhouses. Acta Hortic. 2017, 1164, 383–390. [Google Scholar] [CrossRef]
- Pirayeshfar, F.; Safavi, S.A.; Moayeri, H.R.S.; Messelink, G.J. Provision of astigmatid mites as supplementary food increases the density of the predatory mite Amblyseius swirskii in greenhouse crops, but does not support the omnivorous pest, western flower thrips. BioControl 2021, 66, 511–522. [Google Scholar] [CrossRef]
- Papadopoulos, G.D.; Papadoulis, G.T. Effect of seven different pollens on bio-ecological parameters of the predatory mite Typhlodromus foenilis (Acari: Phytoseiidae). Environ. Entomol. 2008, 37, 340–347. [Google Scholar] [CrossRef]
- Riahi, E.; Fathipour, Y.; Talebi, A.A.; Mehrabadi, M. Pollen quality and predator viability: Life table of Typhlodromus bagdasarjani on seven different plant pollens and two-spotted spider mite. Syst. Appl. Acarol. 2016, 21, 1399–1412. [Google Scholar] [CrossRef]
- Ranabhat, N.B.; Goleva, I.; Zebitz, C.P.W. Life tables of Neoseiulus cucumeris exclusively fed with seven different pollens. BioControl 2014, 59, 195–203. [Google Scholar] [CrossRef]
- Delisle, J.F.; Brodeur, J.; Shipp, L. Evaluation of various types of supplemental food for two species of predatory mites, Amblyseius swirskii and Neoseiulus cucumeris (Acari: Phytoseiidae). Exp. Appl Acarol. 2015, 65, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.T.; Jonckheere, W.; Nguyen, V.H.; Van Leeuwen, T.; De Clercq, P. Life tables and feeding habits of Proprioseiopsis lenis (Acari: Phytoseiidae) and implications for its biological control potential in Southeast Asia. Syst. Appl. Acarol. 2019, 24, 857–865. [Google Scholar]
- Asgari, F.; Moayeri, H.R.S.; Kavousi, A.; Enkegaard, A.; Chi, H. Demography and mass rearing of Amblyseius swirskii (Acari: Phytoseiidae) fed on two species of stored-product mites and their mixture. J. Econ. Entomol. 2020, 113, 2604–2612. [Google Scholar] [CrossRef]
- Li, Y.Y.; Yuan, J.G.; Liu, M.X.; Zhang, Z.H.; Zhou, H.; Liu, H. Evaluation of four artificial diets on demography parameters of Neoseiulus barkeri. BioControl 2021, 66, 789–802. [Google Scholar] [CrossRef]
- Eini, N.; Jafari, S.; Fathipour, Y.; Zalucki, M.P. How pollen grains of 23 plant species affect performance of the predatory mite Neoseiulus californicus. BioControl 2022, 67, 173–187. [Google Scholar] [CrossRef]
- Gotoh, T.; Tsuchiya, A. Effect of multiple mating on reproduction and longevity of the phytoseiid mite Neoseiulus californicus. Exp. Appl. Acarol. 2008, 44, 185–197. [Google Scholar] [CrossRef]
- Wiernasz, D.C.; Hines, J.; Parker, D.G.; Cole, B.J. Mating for variety increases foraging activity in the harvester ant, Pogonomyrmex occidentalis. Mol. Ecol. 2008, 17, 1137–1144. [Google Scholar] [CrossRef]
- Khanamani, M.; Fathipour, Y.; Talebi, A.A.; Mehrabadi, M. Quantitative analysis of long-term mass rearing of Neoseiulus californicus (Acari: Phytoseiidae) on almond pollen. J. Econ. Entomol. 2017, 110, 1442–1450. [Google Scholar] [CrossRef]
- Ghasemzadeh, S.; Leman, A.; Messelink, G.J. Biological control of Echinothrips americanus by phytoseiid predatory mites and the effect of pollen as supplemental food. Exp. Appl. Acarol. 2017, 73, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Nemati, A.; Riahi, E. Does feeding on pollen grains affect the performance of Amblyseius swirskii (Acari: Phytoseiidae) during subsequent generations? Bull. Entomol. Res. 2020, 110, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Yazdanpanah, S.; Fathipour, Y.; Riahi, E.; Zalucki, M.P. Mass production of Neoseiulus cucumeris (Acari: Phytoseiidae): An assessment of 50 generations reared on almond pollen. J. Econ. Entomol. 2021, 114, 2255–2263. [Google Scholar] [CrossRef]
- Rodríguez-Cruz, F.A.; Venzon, M.; Pinto, C.M.F. Performance of Amblyseius herbicolus on broad mites and on castor bean and sunnhemp pollen. Exp. Appl. Acarol. 2013, 60, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Zou, D.Y.; Wu, H.H.; Coudron, T.A.; Zhang, L.S.; Wang, M.Q.; Liu, C.X.; Chen, H.Y. A meridic diet for continuous rearing of Arma chinensis (Hemiptera: Pentatomidae: Asopinae). Biol. Control 2013, 67, 491–497. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Zeng, W.; Deng, W.; Li, M.; Hu, W.; Cai, H.; Li, Y.; Xie, P.; Tan, L.; Zhou, Z. Egg cannibalism varies with sex, reproductive status, and egg and nymph ages in Arma custos (Hemiptera: Asopinae). Front. Ecol. Evol. 2021, 9, 570. [Google Scholar] [CrossRef]
- Roberge, C.; Fréchette, B.; Labrie, G.; Dumont, F.; Lucas, E. Gregarious pupation act as a defensive mechanism against cannibalism and intraguild predation. Insect Sci. 2016, 23, 612–620. [Google Scholar] [CrossRef]
- Ye, J.; Xu, Q.; Li, Z.; Lu, X.; Han, S. Effect of cannibalism on the growth and development of Mallada basalis (Neuroptera: Chrysopidae). Fla. Entomol. 2014, 97, 1075–1080. [Google Scholar] [CrossRef]
- Mendes, J.A.; Barros, M.E.N.; Lima, D.B.; Melo, J.W.S. Cannibalism in Amblyseius largoensis (Muma) (Acari: Phytoseiidae), an important natural enemy of coconut mite pests. Int. J. Acarol. 2017, 43, 387–392. [Google Scholar] [CrossRef]
- Prasad, K.D.; Prasad, B.S.R.V. Biological pest control using cannibalistic predators and with provision of additional food: A theoretical study. Theor. Ecol. 2018, 11, 191–211. [Google Scholar] [CrossRef]
- Zhu, R.; Guo, J.J.; Yi, T.C.; Xiao, R.; Jin, D.C. Functional and numerical responses of Neoseiulus californicus (Mcgregor) to eggs and nymphs of Oulenziella bakeri and Tetranychus urticae. Syst. Appl. Acarol. 2019, 24, 1225–1235. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Natural Prey | Alternative Food | |||||
|---|---|---|---|---|---|---|---|
| T. urticae | Apple Pollen | Almond Pollen | Maize Pollen | O. bakeri | T. putrescentiae | A. ovatus | |
| Egg (d) | 1.47 ± 0.01 (96) a | 1.51 ± 0.04 (78) a | 1.48 ± 0.01 (100) a | 1.51 ± 0.01 (85) a | 1.47 ± 0.01 (94) a | 1.46 ± 0.01 (100) a | 1.49 ± 0.01 (100) a |
| Larva (d) | 0.99 ± 0.01 (85) cd | 1.08 ± 0.03 (66) c | 0.96 ± 0.01 (100) d | 1.83 ± 0.04 (83) a | 1.02 ± 0.01 (88) cd | 1.25 ± 0.03 (90) b | 1.21 ± 0.04 (75) b |
| Protonymph (d) | 1.35 ± 0.03 (84) d | 1.23 ± 0.04 (66) e | 1.30 ± 0.03 (100) de | 1.90 ± 0.04 (82) a | 1.48 ± 0.01 (87) c | 1.46 ± 0.04 (81) c | 1.61 ± 0.04 (65) b |
| Deutonymph (d) | 1.08 ± 0.02 (82) de | 1.28 ± 0.03 (66) b | 1.16 ± 0.02 (99) cd | 1.02 ± 0.01 (82) e | 1.44 ± 0.02 (87) a | 1.18 ± 0.03 (76) c | 1.10 ± 0.03 (62) cde |
| Pre-adult (d) | 4.90 ± 0.03 (82) d | 5.10 ± 0.01 (66) c | 4.91 ± 0.03 (99) d | 6.24 ± 0.05 (82) a | 5.41 ± 0.02 (87) b | 5.32 ± 0.05 (76) b | 5.43 ± 0.06 (63) b |
| Pre-adult survival rate (Sa) § | 0.85 ± 0.04 (96) bc | 0.85 ± 0.04 (78) bc | 0.99 ± 0.01 (100) a | 0.96 ± 0.02 (85) ab | 0.93 ± 0.03 (94) bc | 0.76 ± 0.04 (100) c | 0.62 ± 0.05 (100) d |
| Female longevity (d) | 48.20 ± 1.15 (41) a | 50.36 ± 2.51 (33) a | 39.26 ± 1.26 (50) b | 40.46 ± 1.76 (41) b | 20.38 ± 0.55 (45) d | 27.72 ± 0.92 (43) c | 29.90 ± 0.89 (42) c |
| Male longevity (d) | 30.41 ± 0.77 (41) b | 28.88 ± 1.57 (33) b | 29.98 ± 1.31 (49) b | 36.12 ± 1.68 (41) a | 19.12 ± 0.52 (42) c | 8.61 ± 0.25 (33) d | 10.35 ± 0.42 (20) d |
| Total lifespan (d) | 44.21 ± 1.22 (82) a | 44.72 ± 1.96 (66) a | 39.58 ± 1.02 (99) b | 44.53 ± 1.24 (82) a | 25.18 ± 0.39 (87) c | 24.74 ± 1.21 (76) c | 29.02 ± 1.31 (62) c |
| Total preoviposition period (TPOP) (d) | 7.02 ± 0.14 (41) c | 7.44 ± 0.19 (33) b | 7.99 ± 0.25 (50) b | 11.69 ± 0.37 (37) a | 7.63 ± 0.10 (45) b | 7.77 ± 0.16 (43) b | 7.72 ± 0.17 (41) b |
| Adult preoviposition period (APOP) (d) | 2.00 ± 0.13 (41) c | 2.39 ± 0.11 (33) bc | 3.06 ± 0.25 (50) b | 5.32 ± 0.36 (37) a | 2.18 ± 0.10 (45) c | 2.44 ± 0.15 (43) bc | 2.34 ± 0.15 (41) bc |
| Oviposition period (d) | 25.80 ± 0.55 (41) a | 19.61 ± 1.10 (33) b | 18.14 ± 0.45 (50) b | 7.54 ± 0.31 (37) d | 12.57 ± 0.27 (45) c | 7.67 ± 0.30 (43) d | 6.83 ± 0.32 (41) d |
| Fecundity (eggs/female) | 48.95 ± 0.97 (41) a | 28.55 ± 2.37 (33) b | 26.06 ± 0.70 (50) b | 7.84 ± 0.34 (37) e | 26.02 ± 0.69 (45) b | 10.51 ± 0.46 (43) c | 12.07 ± 0.45 (41) c |
| Parameters | Natural Prey | Alternative Food | |||||
|---|---|---|---|---|---|---|---|
| T. urticae | Apple Pollen | Almond Pollen | Maize Pollen | O. bakeri | T. putrescentiae | A. ovatus | |
| r (day−1) | 0.210 ± 0.011 a | 0.155 ± 0.012 b | 0.163 ± 0.008 b | 0.064 ± 0.008 d | 0.202 ± 0.010 a | 0.120 ± 0.011 c | 0.137 ± 0.012 bc |
| λ (day−1) | 1.233 ± 0.014 a | 1.168 ± 0.014 b | 1.177 ± 0.010 b | 1.066 ± 0.008 d | 1.224 ± 0.012 a | 1.128 ± 0.012 c | 1.147 ± 0.014 bc |
| R0 (offspring/individual) | 20.906 ± 2.494 a | 12.077 ± 1.694 b | 13.030 ± 1.348 b | 3.412 ± 0.448 d | 12.457 ± 1.384 b | 4.520 ± 0.555 cd | 4.950 ± 0.620 c |
| T (d) | 14.509 ± 0.261 c | 16.078 ± 0.558 b | 15.708 ± 0.293 b | 19.081 ± 0.620 a | 12.488 ± 0.142 d | 12.566 ± 0.339 d | 11.693 ± 0.270 e |
| Cannibalism Rate | df | Sum sq | H | p Value |
|---|---|---|---|---|
| Diet type | 6 | 18662687 | 455.80 | 0.00000 |
| Predation days | 6 | 350979 | 8.57 | 0.19912 |
| Diet type: predation days | 36 | 1034180 | 25.26 | 0.90970 |
| Residuals | 686 | 10005694 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, F.; Ni, Z.-H.; Zou, M.-T.; Zhu, R.; Yi, T.-C.; Guo, J.-J.; Jin, D.-C. The Effects of Alternative Foods on Life History and Cannibalism of Amblyseius herbicolus (Acari: Phytoseiidae). Insects 2022, 13, 1036. https://doi.org/10.3390/insects13111036
Hou F, Ni Z-H, Zou M-T, Zhu R, Yi T-C, Guo J-J, Jin D-C. The Effects of Alternative Foods on Life History and Cannibalism of Amblyseius herbicolus (Acari: Phytoseiidae). Insects. 2022; 13(11):1036. https://doi.org/10.3390/insects13111036
Chicago/Turabian StyleHou, Fei, Zhao-Hong Ni, Meng-Ting Zou, Rui Zhu, Tian-Ci Yi, Jian-Jun Guo, and Dao-Chao Jin. 2022. "The Effects of Alternative Foods on Life History and Cannibalism of Amblyseius herbicolus (Acari: Phytoseiidae)" Insects 13, no. 11: 1036. https://doi.org/10.3390/insects13111036