
DNA Barcoding Identifies Unknown Females and Larvae of Fannia R.-D. (Diptera: Fanniidae) from Carrion Succession Experiment and Case Report

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. DNA Extraction, Amplification and Sequencing
2.3. Sequence Alignment and Data Analysis
3. Case Report
4. Results
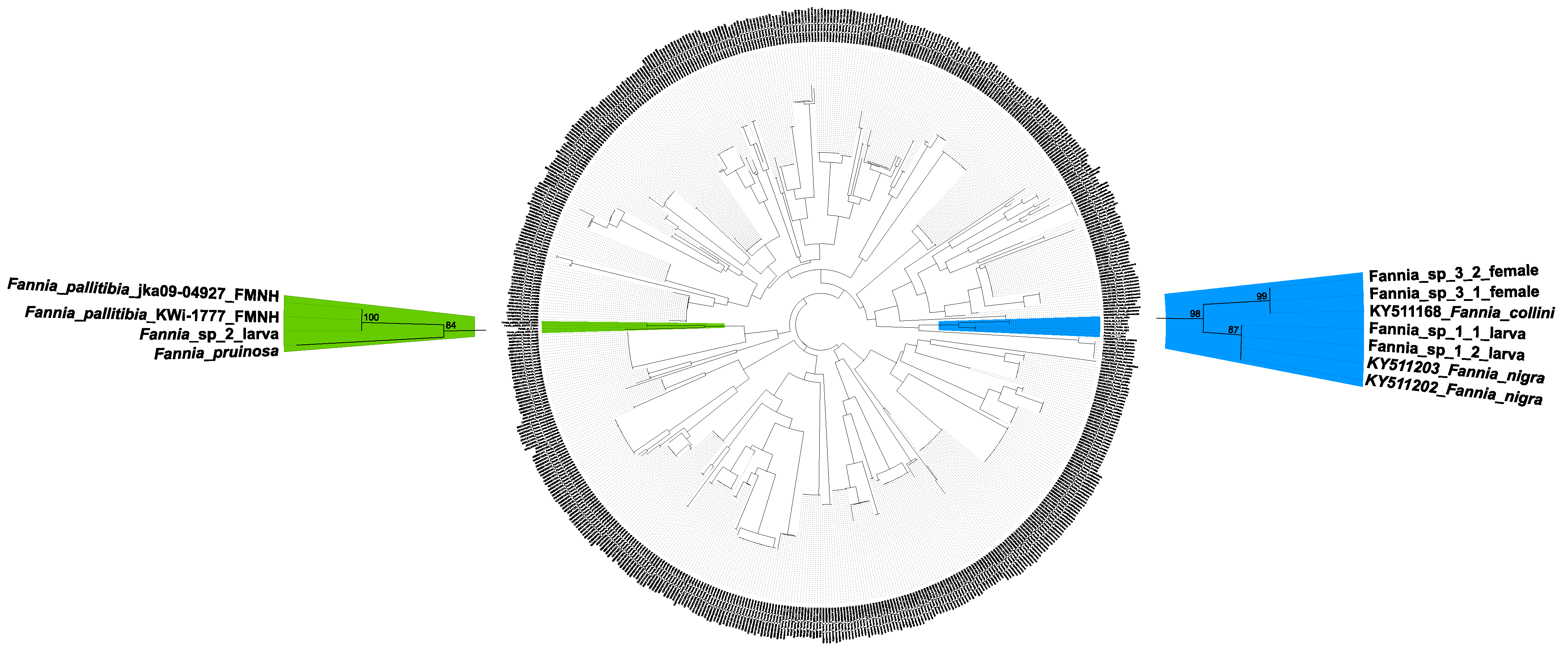
4.1. Molecular Data and Identification
4.2. Fannia nigra Malloch, 1910
4.3. Fannia pallitibia (Rondani, 1866)
4.4. Fannia collini d’Assis-Fonseca, 1966
5. Discussion
5.1. Taxonomy
- 49.
- Two prealar setae, anterior prealar seta longer than half length of posterior notopleural seta and inserted nearer to suture than to supraalar seta…49a
- 49a.
- Hind tibia with 8–10 short anterodorsal setae in addition with 1 strong seta; lower orbital setae in the middle of fronto-orbital plates… F. immutica Collin
- ‒
- Hind tibia with 1 strong anterodorsal seta, without additional setae above; lower orbital setae inserted nearer to outer margin of fronto-orbital plates… F. collini d’Assis-Fonseca
- ‒
- Only one weak prealar seta inserted nearer to supra-alar seta…50
5.2. Forensic Importance
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grzywacz, A.; Jarmusz, M.; Szpila, K. New and noteworthy records of carrion-visiting Fannia Robineau-Desvoidy (Diptera: Fanniidae) of Poland. Entomol. Fenn. 2018, 29, 169–174. [Google Scholar] [CrossRef]
- Grzywacz, A.; Castro, C.P. New records of Fannia Robineau-Desvoidy (Diptera: Fanniidae) collected on pig carrion in Portugal with additional data on the distribution of F. conspecta Rudzinski. Entomol. Fenn. 2012, 23, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Mądra, A.; Frątczak, K.; Grzywacz, A.; Matuszewski, S. Long-term study of pig carrion entomofauna. Forensic Sci. Int. 2015, 252, 1–10. [Google Scholar] [CrossRef]
- Velásquez, Y.; Martínez-Sánchez, A.; Rojo, S. First record of Fannia leucosticta (Meigen) (Diptera: Fanniidae) breeding in human corpses. Forensic Sci. Int. 2013, 229, 13–15. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, S.D.; Costa, D.L.; Oliveira, D.L. Entomological evidence in a case of a suicide victim by hanging: First collaboration between entomologists and forensic police in north-eastern Brazil. Aust. J. Forensic Sci. 2019, 51, 231–239. [Google Scholar] [CrossRef]
- Bourel, B.; Tournel, G.; Hédouin, V.; Gosset, D. Entomofauna of buried bodies in northern France. Int. J. Legal Med. 2004, 118, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Mariani, R.; García-Mancuso, R.; Varela, G.L.; Inda, A.M. Entomofauna of a buried body: Study of the exhumation of a human cadaver in Buenos Aires, Argentina. Forensic Sci. Int. 2014, 237, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Benecke, M.; Lessig, R. Child neglect and forensic entomology. Forensic Sci. Int. 2001, 120, 155–159. [Google Scholar] [CrossRef]
- Benecke, M.; Josephi, E.; Zweihoff, R. Neglect of the elderly: Forensic entomology cases and considerations. Forensic Sci. Int. 2004, 146S, S195–S199. [Google Scholar] [CrossRef]
- Grzywacz, A.; Wyborska, D.; Piwczyński, M. DNA barcoding allows identification of European Fanniidae (Diptera) of forensic interest. Forensic Sci. Int. 2017, 278, 106–114. [Google Scholar] [CrossRef]
- Holloway, B.A. Larvae of New Zealand Fanniidae (Diptera: Calyptrata). N. Z. J. Entomol. 1984, 11, 239–258. [Google Scholar] [CrossRef]
- Lyneborg, L. Taxonomy of European Fannia larvae (Diptera, Fanniidae). Staatl. Mus. Nat. 1970, 215, 1–28. [Google Scholar]
- Szpila, K.; Grzywacz, A. Larvae of the North American Calyptratae flies of forensic importance. In Forensic Entomology: The Utility of Arthropods in Legal Investigations; Byrd, J.H., Tomberlin, J.K., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 531–545. [Google Scholar] [CrossRef]
- Grisales, D.; Lecheta, M.C.; Aballay, F.H.; de Carvalho, C.J.B. A key and checklist to the Neotropical forensically important “Little House Flies” (Diptera: Fanniidae). Zoologia 2016, 33, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Fiedler, A.; Halbach, M.; Sinclair, B.; Benecke, M. What is the edge of a forest? A diversity analysis of adult Diptera found on decomposing piglets inside and on the edge of a western German woodland inspired by a courtroom question. Entomol. Heute 2008, 20, 173–191. [Google Scholar]
- Jarmusz, M.; Grzywacz, A.; Bajerlein, D. A comparative study of the entomofauna (Coleoptera, Diptera) associated with hanging and ground pig carcasses in a forest habitat of Poland. Forensic Sci. Int. 2020, 309, 110212. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, N.I.; Domínguez, M.C. A new species of the genus Fannia Robineau-Desvoidy (Diptera: Fanniidae) belonging to the canicularis species group, collected on pig carrion in the Yungas of the province of Jujuy, Argentina. Stud. Neotrop. Fauna Environ. 2010, 45, 95–100. [Google Scholar] [CrossRef]
- Faria, L.S.; Paseto, M.L.; Franco, F.T.; Perdigão, V.C.; Capel, G.; Mendes, J. Insects breeding in pig carrion in two environments of a rural area of the state of minas gerais, Brazil. Neotrop. Entomol. 2013, 42, 216–222. [Google Scholar] [CrossRef]
- Bohart, G.E.; Gressitt, J.L. Filth-inhabiting flies of Guam. Bernice Pauahi Bish. Mus. Bull. 1951, 204, 1–151. [Google Scholar]
- Grzywacz, A.; Pape, T.; Szpila, K. Larval morphology of the lesser housefly, Fannia canicularis. Med. Vet. Entomol. 2012, 26, 70–82. [Google Scholar] [CrossRef]
- Carmo, R.F.R.; Oliveira, D.L.; Barbosa, T.M.; Soares, T.F.; Souza, J.R.B.; Vasconcelos, S.D. Visitors versus colonizers: An empirical study on the use of vertebrate carcasses by necrophagous Diptera in a rainforest fragment. Ann. Entomol. Soc. Am. 2017, 110, 492–500. [Google Scholar] [CrossRef]
- Pont, A.C. A Revision of Australian Fanniidae (Diptera: Calyptrata). Aust. J. Zool. Suppl. Ser. 1977, 51, 1–60. [Google Scholar] [CrossRef]
- Smith, K.G.V. A Manual of Forensic Entomology; British Museum (Natural History): London, UK; Cornell University Press: Ithaca, NY, USA, 1986. [Google Scholar]
- Domínguez, M.C.; Pont, A.C. Fanniidae (Insecta: Diptera). Fauna N. Z. 2014, 71, 1–91. [Google Scholar]
- Huckett, H.C. The Muscidae of California: Exclusive of subfamilies Muscinae and Stomoxyinae. Bull. Calif. Insect Surv. 1975, 18, 1–148. [Google Scholar]
- de Carvalho, C.J.B.; Mello-Patiu, C.A. de Key to the adults of the most common forensic species of Diptera in South America. Rev. Bras. Entomol. 2008, 52, 390–406. [Google Scholar] [CrossRef] [Green Version]
- Rozkošný, R.; Gregor, F.; Pont, A.C. The European Fanniidae (Diptera). Acta Sci. Nat. Brno 1997, 31, 1–80. [Google Scholar]
- Chillcott, J.G. A revision of the Nearctic species of Fanniinae (Diptera: Muscidae). Can. Entomol. 1960, 92, 5–295. [Google Scholar] [CrossRef]
- Jarmusz, M.; Bajerlein, D. Decomposition of hanging pig carcasses in a forest habitat of Poland. Forensic Sci. Int. 2019, 300, 32–42. [Google Scholar] [CrossRef]
- Courtney, G.W.; Sinclair, B.J.; Meier, R. Morphology and terminology of Diptera larvae. In Contributions to a Manual of Palaearctic Diptera (with Special Reference to Flies of Economic Importance); Papp, L., Darvas, B., Eds.; Science Herald Press: Budapest, Hungary, 2000; pp. 85–161. [Google Scholar]
- Grzywacz, A.; Wallman, J.F.; Piwczyński, M. To be or not to be a valid genus: The systematic position of Ophyra R.-D. revised (Diptera: Muscidae). Syst. Entomol. 2017, 42, 714–723. [Google Scholar] [CrossRef] [Green Version]
- Bernasconi, M.V.; Valsangiacomo, C.; Piffaretti, J.C.; Ward, P.I. Phylogenetic relationships among Muscoidea (Diptera: Calyptratae) based on mitochondrial DNA sequences. Insect Mol. Biol. 2000, 9, 67–74. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Szpila, K. Key for the Identification of Third Instars of European Blowflies (Diptera: Calliphoridae) of Forensic Importance. In Current Concepts in Forensic Entomology; Amendt, J., Campobasso, C., Pietro Goff, M.L., Grassberger, M., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2010; pp. 43–56. [Google Scholar] [CrossRef]
- Fremdt, H.; Szpila, K.; Huijbregts, J.; Lindström, A.; Zehner, R.; Amendt, J. Lucilia silvarum Meigen, 1826 (Diptera: Calliphoridae)—A new species of interest for forensic entomology in Europe. Forensic Sci. Int. 2012, 222, 335–339. [Google Scholar] [CrossRef]
- Martín-Vega, D.; Díaz-Aranda, L.M.; Baz, A. The immature stages of the necrophagous fly Liopiophila varipes and considerations on the genus Liopiophila (Diptera: Piophilidae). Dtsch. Entomol. Z. 2014, 61, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Grassberger, M.; Friedrich, E.; Reiter, C. The blowfly Chrysomya albiceps (Wiedemann) (Diptera: Calliphoridae) as a new forensic indicator in Central Europe. Int. J. Legal Med. 2003, 117, 75–81. [Google Scholar] [CrossRef]
- Richards, C.S.; Paterson, I.D.; Villet, M.H. Estimating the age of immature Chrysomya albiceps (Diptera: Calliphoridae), correcting for temperature and geographical latitude. Int. J. Legal Med. 2008, 122, 271–279. [Google Scholar] [CrossRef]
- Meier, R.; Shiyang, K.; Vaidya, G.; Ng, P.K.L. DNA barcoding and taxonomy in Diptera: A tale of high intraspecific variability and low identification success. Syst. Biol. 2006, 55, 715–728. [Google Scholar] [CrossRef] [Green Version]
- Collins, R.A.; Cruickshank, R.H. The seven deadly sins of DNA barcoding. Mol. Ecol. Resour. 2013, 13, 969–975. [Google Scholar] [CrossRef]
- Meyer, C.P.; Paulay, G. DNA Barcoding: Error rates based on comprehensive sampling. PLoS Biol. 2005, 3, e422. [Google Scholar] [CrossRef] [Green Version]
- Kahanpää, J.; Haarto, A. Notes on Fanniidae (Diptera) of Finland, with a description of the female of Fannia stigi Rognes, 1982. Entomol. Fenn. 2013, 24, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Wang, Q.; Wang, M. Description of females of Fannia imperatoria Nishida and Phaonia vagata Xue & Wang (Diptera: Muscoidea). Entomol. Fenn. 2019, 22, 274–278. [Google Scholar] [CrossRef] [Green Version]
- Barták, M.; Preisler, J.; Kubík, Š.; Šuláková, H.; Sloup, V. Fanniidae (Diptera): New synonym, new records and an updated key to males of European species of Fannia. Zookeys 2016, 593, 91–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- d’Assis Fonseca, E.C.M. Diptera Cyclorrhapha Calyptrata Section (b) Muscidae. Handb. Identif. Br. Insects 1968, 10, 1–118. [Google Scholar]
- Sonet, G.; Jordaens, K.; Braet, Y.; Bourguignon, L.; Dupont, E.; Backeljau, T.; De Meyer, M.; Desmyter, S. Utility of GenBank and the Barcode of Life Data Systems (BOLD) for the identification of forensically important Diptera from Belgium and France. Zookeys 2013, 365, 307–328. [Google Scholar] [CrossRef] [PubMed]
- Marchenko, M.I. Medicolegal relevance of cadaver entomofauna for the determination of the time of death. Forensic Sci. Int. 2001, 120, 89–109. [Google Scholar] [CrossRef]
- Shabani Kordshouli, R.; Grzywacz, A.; Akbarzadeh, K.; Azam, K.; AliMohammadi, A.; Ghadi Pasha, M.; Ali Oshaghi, M. Thermal requirements of immature stages of Chrysomya albiceps (Diptera: Calliphoridae) as a common forensically important fly. Sci. Justice 2021. [Google Scholar] [CrossRef]
- Matuszewski, S.; Bajerlein, D.; Konwerski, S.; Szpila, K. Insect succession and carrion decomposition in selected forests of Central Europe. Part 2: Composition and residency patterns of carrion fauna. Forensic Sci. Int. 2010, 195, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Grzywacz, A.; Hall, M.J.R.; Pape, T.; Szpila, K. Muscidae (Diptera) of forensic importance—An identification key to third instar larvae of the western Palaearctic region and a catalogue of the muscid carrion community. Int. J. Legal Med. 2017, 131, 855–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szpila, K. Annotated list of blowflies (Diptera: Calliphoridae) recorded during studies of insect succession on large carrion in Poland. Dipteron 2017, 33, 85–93. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grzywacz, A.; Jarmusz, M.; Walczak, K.; Skowronek, R.; Johnston, N.P.; Szpila, K. DNA Barcoding Identifies Unknown Females and Larvae of Fannia R.-D. (Diptera: Fanniidae) from Carrion Succession Experiment and Case Report. Insects 2021, 12, 381. https://doi.org/10.3390/insects12050381
Grzywacz A, Jarmusz M, Walczak K, Skowronek R, Johnston NP, Szpila K. DNA Barcoding Identifies Unknown Females and Larvae of Fannia R.-D. (Diptera: Fanniidae) from Carrion Succession Experiment and Case Report. Insects. 2021; 12(5):381. https://doi.org/10.3390/insects12050381
Chicago/Turabian StyleGrzywacz, Andrzej, Mateusz Jarmusz, Kinga Walczak, Rafał Skowronek, Nikolas P. Johnston, and Krzysztof Szpila. 2021. "DNA Barcoding Identifies Unknown Females and Larvae of Fannia R.-D. (Diptera: Fanniidae) from Carrion Succession Experiment and Case Report" Insects 12, no. 5: 381. https://doi.org/10.3390/insects12050381