Abundance and Distribution of Korean Flower Flies (Diptera: Syrphidae): Dominant Influence of Latitude on Regional Distribution


Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sampling
2.2. Data Analysis
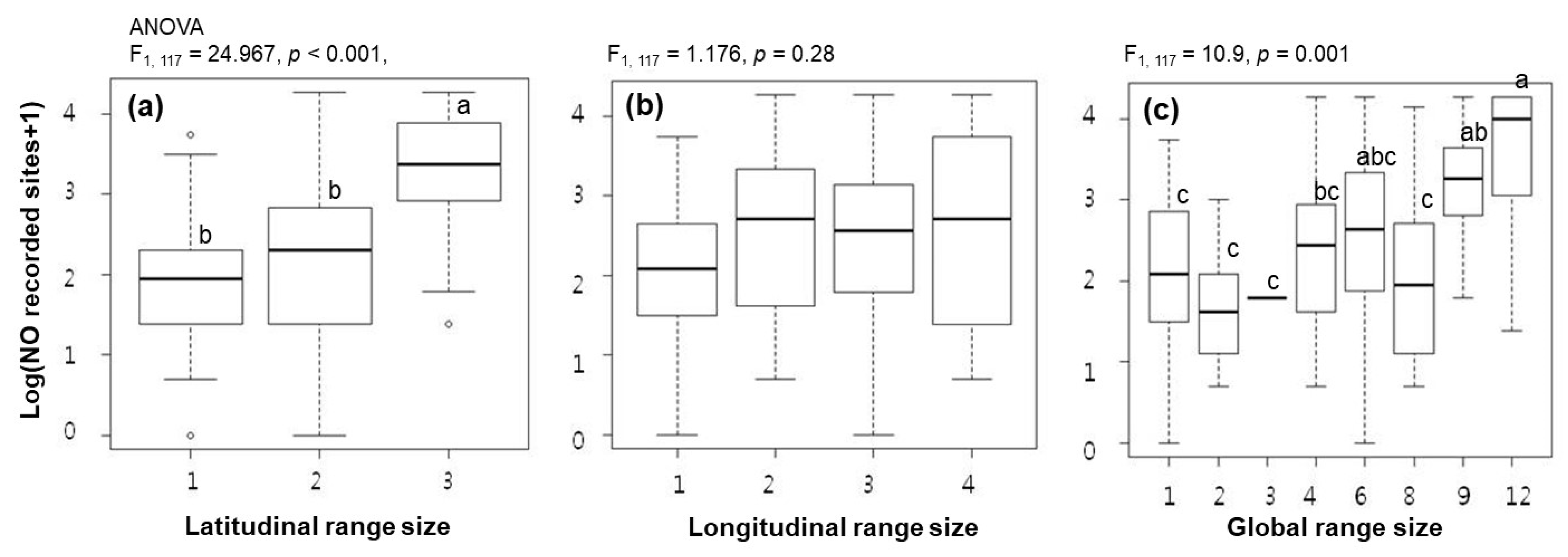
3. Results
4. Discussion
4.1. Local and Regional Occurrence
4.2. Regional and Global Occurrence
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Borregaard, M.K.; Rahbek, G. Causality of the relationship between geographic distribution and species abundance. Quart. Rev. Biol. 2010, 85, 3–25. [Google Scholar] [CrossRef] [Green Version]
- Collins, S.L.; Glenn, S.M. Importance of spatial and temporal dynamics in species regional abundance and distribution. Ecology 1991, 72, 654–664. [Google Scholar] [CrossRef]
- Zuckerberg, B.; Woods, A.M.; Porter, W.F. Poleward shifts in breeding bird distributions in New York State. Glob. Chang. Biol. 2009, 15, 1866–1883. [Google Scholar] [CrossRef]
- Brown, J.H. On the relationship between abundance and distribution of species. Am. Nat. 1984, 124, 255–279. [Google Scholar] [CrossRef]
- Hartley, S.H. A positive relationship between local abundance and regional occupancy is almost inevitable (but not all positive relationships are the same). J. Anim. Ecol. 1998, 67, 992–994. [Google Scholar] [CrossRef] [PubMed]
- Bell, G. Neutral macroecology. Science 2001, 293, 2413–2418. [Google Scholar] [CrossRef]
- Bock, C.E. Distribution-abundance relationships of some Arizona landbirds: A matter of scale? Ecology 1987, 68, 124–129. [Google Scholar] [CrossRef]
- Frisk, M.G.; Duplisea, D.E.; Trenkei, V.M. Exploring the abundance-occupancy relationships for the Georges Bank finfish and shellfish community from 1963 to 2006. Ecol. Appl. 2011, 21, 227–240. [Google Scholar] [CrossRef]
- Gaston, K.J.; Borges, P.A.V.; He, F.; Gaspar, C. Abundance, spatial variance and occupancy: Arthropod species distribution in the Azores. J. Anim. Ecol. 2006, 75, 646–656. [Google Scholar] [CrossRef] [Green Version]
- Verberk, W.C.E.P.; van der Velde, G.; Esselink, H. Explaining abundance-occupancy relationships in specialists and generalists: A case study on aquatic macroinvertebrates in standing waters. J. Anim. Ecol. 2010, 79, 589–601. [Google Scholar] [CrossRef]
- Kim, S.-S.; Kwon, T.-S. Changes in butterfly assemblages and increase of open-land inhabiting species after forest flies. J. Asia-Pac. Biodivers. 2018, 11, 39–48. [Google Scholar] [CrossRef]
- Owen, J.; Gilbert, F.S. On the abundance of hoverflies (Syrphidae). Oikos 1989, 55, 183–193. [Google Scholar] [CrossRef]
- Petanidou, T.; Vujic, A.; Ellis, W. Hoverfly diversity (Diptera: Syrphidae) in a Mediterranean scrub community near Athens, Greece. Ann. Soc. Entomol. Fr. 2011, 47, 168–175. [Google Scholar] [CrossRef]
- Bankowska, R. Hover flies (Diptera, Syrphidae) of moist meadows on the Mazovian lowland. Memorab. Zool. 1989, 43, 329–347. [Google Scholar]
- Gilbert, F.S.; Harding, E.F.; Line, J.M.; Perry, I. Morphological approaches to community structure in hoverflies (Diptera, Syrphidae). Proc. the R. Soc. B 1985, 224, 115–130. [Google Scholar]
- Gibbons, D.W.; Donald, P.F.; Bauer, H.-G.; Fornasari, L.; Dawson, I.K. Mapping avian distributions: The evolution of bird atlases. Bird Study 2007, 54, 324–334. [Google Scholar] [CrossRef] [Green Version]
- Kwon, T.S.; Lee, C.M.; Kim, S. Northward range shifts in Korean butterflies. Clim. Chang. 2014, 126, 163–174. [Google Scholar] [CrossRef]
- Donald, P.F.; Fuller, R.J. Ornithological atlas data: A review of uses and limitations. Bird Study 1998, 45, 129–145. [Google Scholar] [CrossRef] [Green Version]
- Robertson, M.P.; Cumming, G.S.; Erasmus, B.F.N. Getting the most out of atlas data. Divers. Distrib. 2010, 16, 363–375. [Google Scholar] [CrossRef]
- Han, H.-Y.; Choi, D.-S. Diptera (Syrphidae): Economic Insects of Korea 15, Insecta Koreana Suppl. 22; National Institute of Agricultural Science and Technology: Suwon, Korea, 2001. [Google Scholar]
- Abellán, P.; Ribera, I. Geographic location and phylogeny are the main determinants of the size of the geographical range in aquatic beetles. BMC Evol. Biol. 2011, 11, 344. [Google Scholar] [CrossRef] [Green Version]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Luo, B.; Santana, S.E.; Pang, Y.; Wang, M.; Xiao, Y.; Feng, J. Wing morphology predicts geographic range size in vespertilionid bats. Sci. Rep. 2019, 9, 4526. [Google Scholar] [CrossRef] [PubMed]
- Malacrida, A.R.; Gomulski, L.M.; Bonizzoni, M.; Bertin, S.; Gasperi, G.; Guglielmino, C.R. Globalization and fruitfly invasion and expansion: The medfly paradigm. Genetica 2007, 131, 1. [Google Scholar] [CrossRef] [PubMed]
- Gittings, T.; O’Halloran, J.; Kelly, T.; Giller, P.S. The contribution of open spaces to the maintenance of hoverfly (Diptera, Syrphidae) biodiversity in Irish plantation forests. For. Ecol. Manag. 2006, 237, 290–300. [Google Scholar] [CrossRef]
- Lee, C.M.; Kwon, T.-S.; Ji, O.Y.; Kim, S.-S.; Park, G.-E.; Lim, J.-H. Prediction of abundance of forest flies (Diptera) according to climate scenarios RCP 4.5 and RCP 8.5 in South Korea. J. Asia-Pac. Biodivers. 2015, 349–370. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.M.; Kwon, T.-S.; Cheon, K. Response of ground beetles (Coleoptera: Carabidae) to forest gaps formed by a typhoon in a red pine forest at Gwangneung Forest, Republic of Korea. J. For. Res. 2017, 28, 173–181. [Google Scholar] [CrossRef]
- Lee, C.M.; Kwon, T.S. Change of butterfly communities after clear cutting in Gwangneung forest. Korean J. Appl. Entomol. 2014, 53, 347–354. [Google Scholar] [CrossRef]
- Kwon, T.-S. Distribution of Korean Ants: Temperature and Habitat; National Institute of Forest Science: Seoul, Korea, 2018. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 3 March 2017).
- da Silva, L.J. Laercio: Duncan Test, Tukey Test and Scott-Knott Test. 2010. Available online: https://CRAN.R-project.org/package=laercio (accessed on 23 June 2019).
- Byun, B.-K.; Lee, B.-W.; Park, S.-Y.; Lee, Y.-M.; Jo, D.-G. Insect fauna of Mt. Nam-san in Seoul, Korea. J. Korean Nat. 2009, 2, 137–153. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.-B.; Yoon, S.-J.; Yoon, S.-M.; Ryu, J.-W.; Min, H.-K.; Oh, K.-S. Insect fauna of Gyeongju National Park, Korea. J. Korean Nat. 2008, 1, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Hong, E.-J.; Jeon, Y.-L.; Yoon, J.-C.; Kim, J.-Y.; Lee, M.-H.; Kim, J.-W.; Park, S.-J.; Kim, K.-G.; Kim, J.-H.; Kim, B.-J. Insect diversity of Mt. Oseosan. J. Korean Nat. 2012, 5, 251–266. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.-W.; Lim, J.-S.; Park, S.-Y.; Jo, D.-G. Insect fauna of Mt. Jan-san, Yeongwol-gun, Gangwon-do, Korea. J. Korean Nat. 2011, 4, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.-S.; Park, S.-Y.; Lim, J.-O.; Lee, B.-W. A faunistic study of insects from Is. Ulleungdo and its nearby islands in South Korea. J. Asia-Pac. Biodivers. 2013, 6, 93–121. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-J.; Lim, H.-M.; Hong, E.-J.; Jeon, Y.-L.; Kim, B.-J. Survey on insect diversity of Yeonpyeong-do Island, Korea. J. Korean Nat. 2012, 5, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Ricarte, A.; Ángeles Marcos-García, M.; Moreno, C.E. Assessing the effects of vegetation type on hoverfly (Diptera: Syrphidae) diversity in a Mediterranean landscape: Implications for conservation. J. Insect Conserv. 2011, 15, 865–877. [Google Scholar] [CrossRef]
- Wilson, P.D. The pervasive influence of sampling and methodological artefacts on a macroecological pattern: The abundance-occupancy relationship. Glob. Ecol. Biog. 2008, 17, 457–464. [Google Scholar] [CrossRef]
- Dennis, E.B.; Brereton, T.M.; Morgan, B.J.T.; Fox, R.; Shortall, C.R.; Prescott, T.; Foster, S. Trends and indicators for quantifying moth abundance and occupancy in Scotland. J. Insect Conserv. 2019, 23, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, T.B.; Cassey, P.; Gaston, K.J. Variations on a theme: Sources of heterogeneity in the form of the interspecific relationship between abundance and distribution. J. Anim. Ecol. 2006, 75, 1426–1439. [Google Scholar] [CrossRef]
- Stomp, M.; Huisman, J.; Mittelbach, G.G.; Litchman, E.; Klausmeier, C.A. Large-scale biodiversity patterns in freshwater phytoplankton. Ecology 2011, 92, 2096–2107. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Dependent Variable | Independent Variable | Coefficient | P (t-test) | adj. R2 | AIC |
|---|---|---|---|---|---|
| Abundance | National occupancy | 0.5566 | <0.05 | 0.085 | 172.7 |
| Latitudinal range size (Lat) | −0.4439 | ns | |||
| Longitudinal range size (Log) | 0.2147 | ns | |||
| Abundance | National occupancy | 0.491 | <0.05 | 0.077 | 172.2 |
| Global range size (Lat × Log) | −0.006 | ns | |||
| Abundance | National occupancy | 0.4858 | <0.05 | 0.098 | 170.2 |
| Local occupancy | National occupancy | 0.68765 | <0.05 | 0.093 | 188.9 |
| Latitudinal range size (Lat) | −0.85275 | ns | |||
| Longitudinal range size (Log) | 0.03561 | ns | |||
| Local occupancy | National occupancy | 0.59422 | <0.05 | 0.073 | 189 |
| Global range size (Lat × Log) | −0.11588 | ns | |||
| Local occupancy | National occupancy | 0.4921 | 0.0502 | 0.064 | 188.5 |
| National occupancy | Latitudinal range size | 0.8774 | <0.001 | 0.177 | 338.1 |
| Longitudinal range size | −0.1609 | ns | |||
| National occupancy | Global range size (Lat × Log) | 0.1099 | <0.01 | 0.077 | 350.8 |
| National occupancy | Latitudinal range size | 0.7502 | <0.001 | 0.169 | 338.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, T.-S.; Lee, C.M.; Ji, O.Y.; Athar, M.; Jung, S.C.; Lim, J.-H.; Park, Y.-S. Abundance and Distribution of Korean Flower Flies (Diptera: Syrphidae): Dominant Influence of Latitude on Regional Distribution. Insects 2020, 11, 213. https://doi.org/10.3390/insects11040213
Kwon T-S, Lee CM, Ji OY, Athar M, Jung SC, Lim J-H, Park Y-S. Abundance and Distribution of Korean Flower Flies (Diptera: Syrphidae): Dominant Influence of Latitude on Regional Distribution. Insects. 2020; 11(4):213. https://doi.org/10.3390/insects11040213
Chicago/Turabian StyleKwon, Tae-Sung, Cheol Min Lee, Ok Yeong Ji, Mohammad Athar, Sung Cheol Jung, Jong-Hwan Lim, and Young-Seuk Park. 2020. "Abundance and Distribution of Korean Flower Flies (Diptera: Syrphidae): Dominant Influence of Latitude on Regional Distribution" Insects 11, no. 4: 213. https://doi.org/10.3390/insects11040213