

Rel Family Transcription Factor NFAT5 Upregulates COX2 via HIF-1α Activity in Ishikawa and HEC1a Cells
 , , , ,
, , , ,
Abstract
:1. Introduction
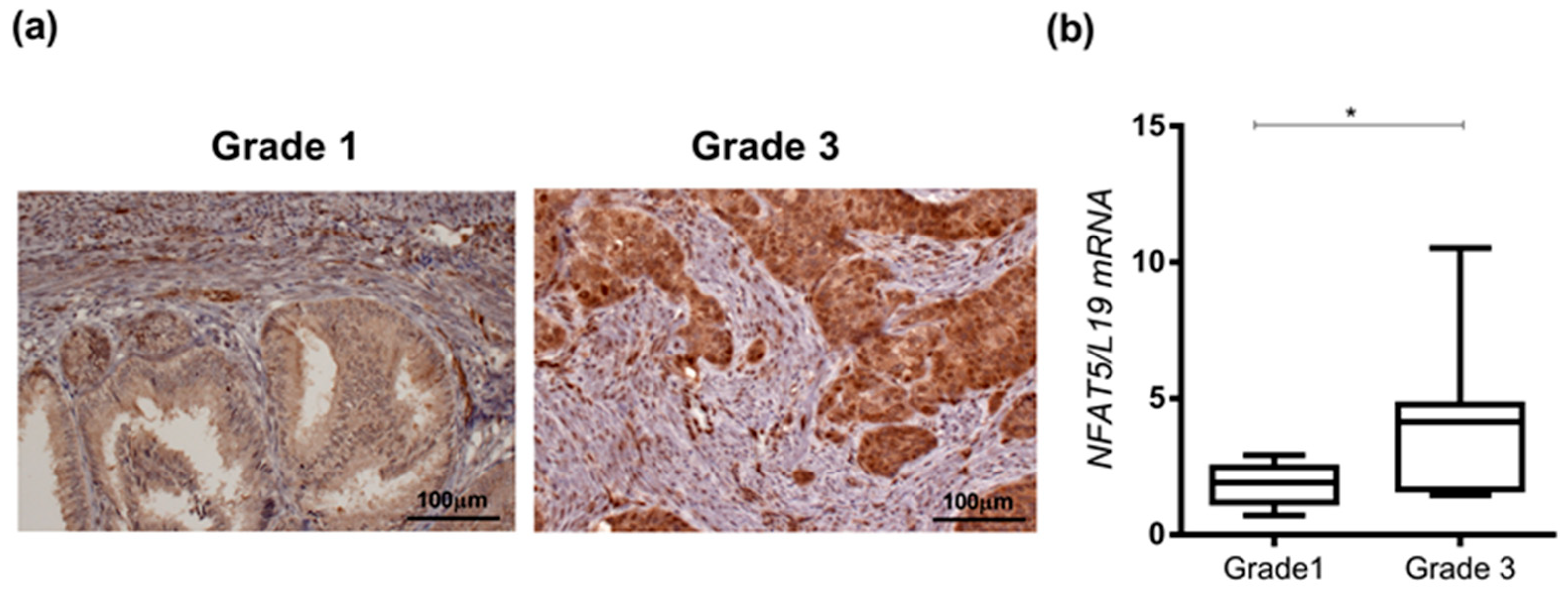
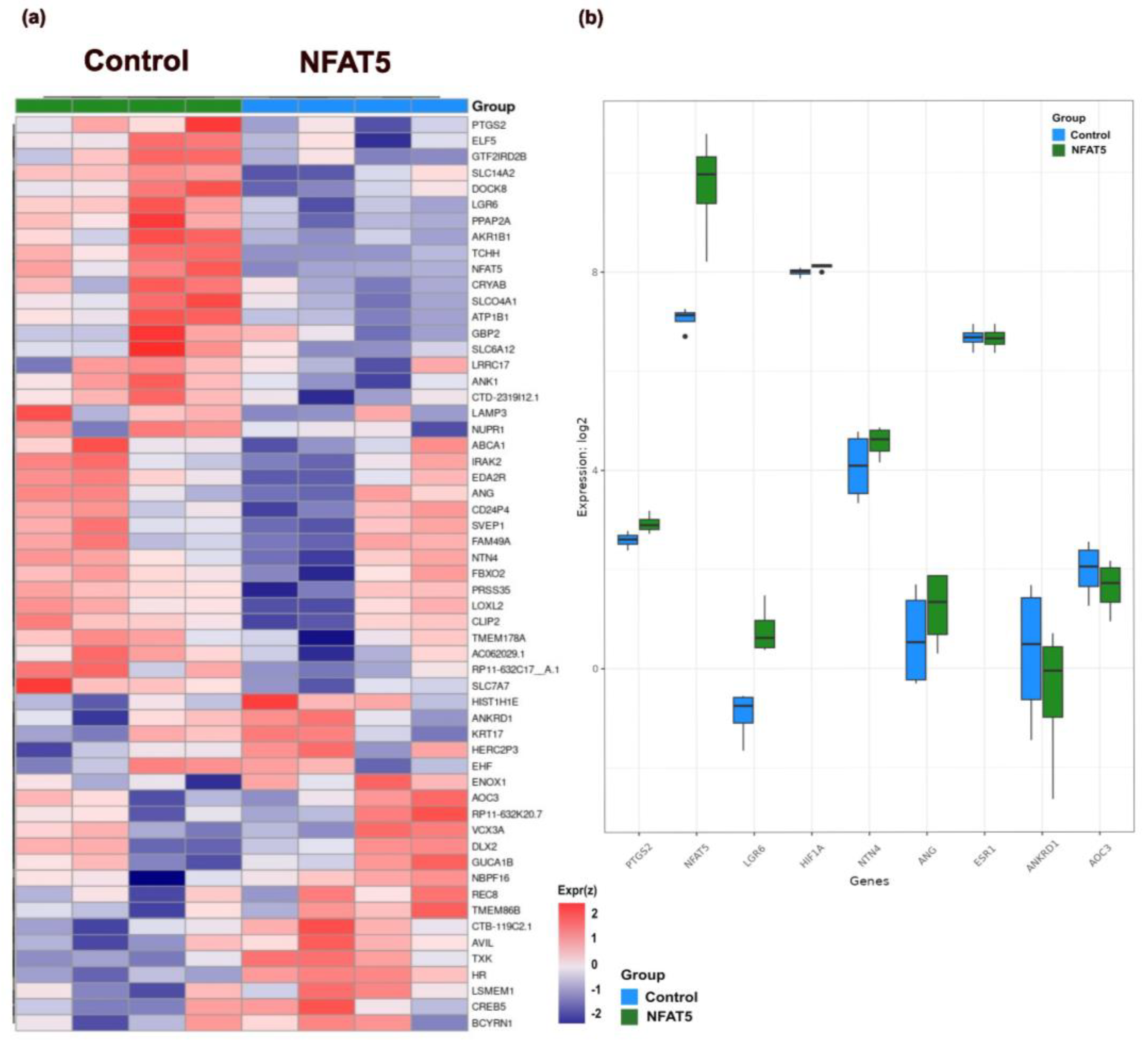
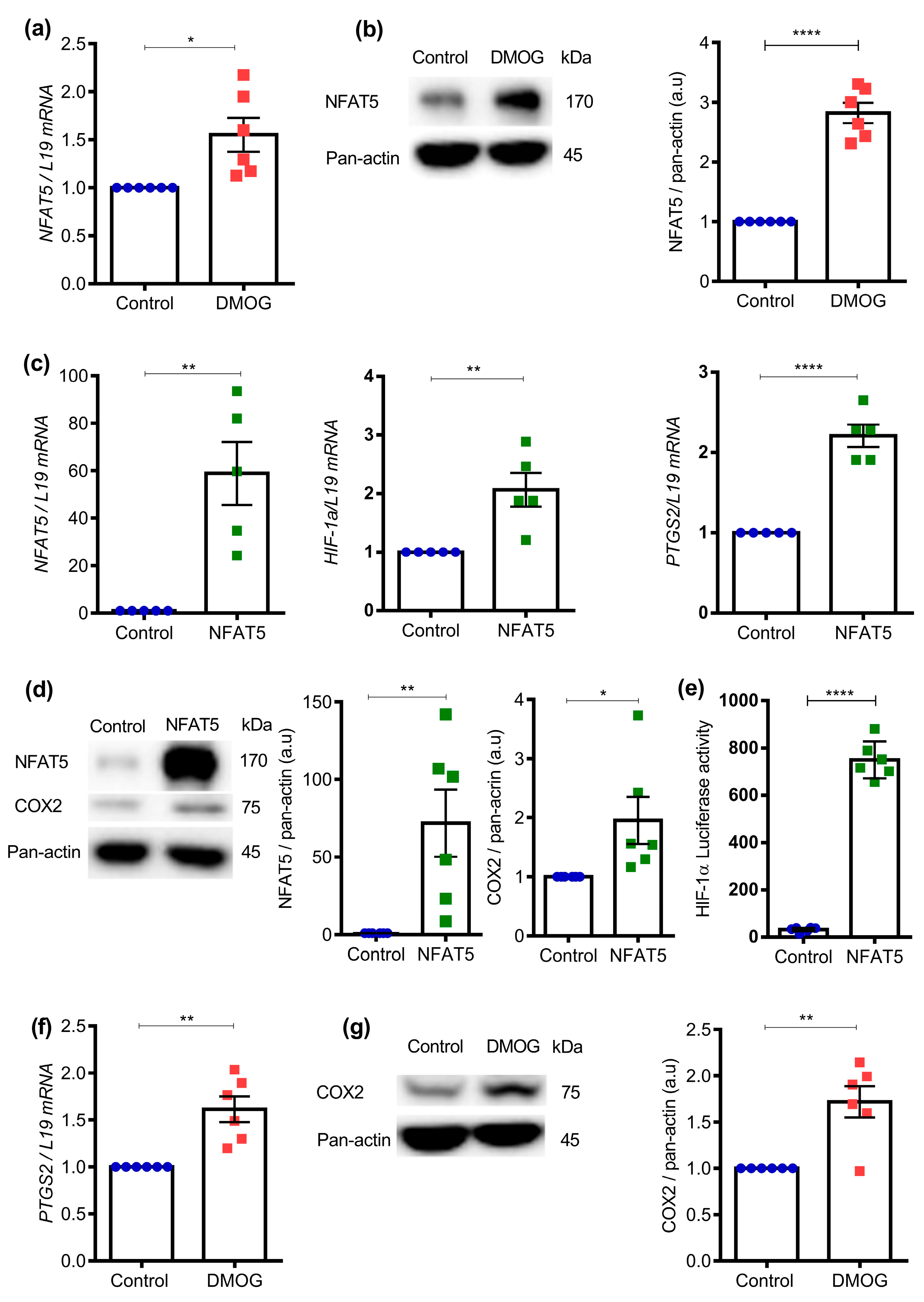
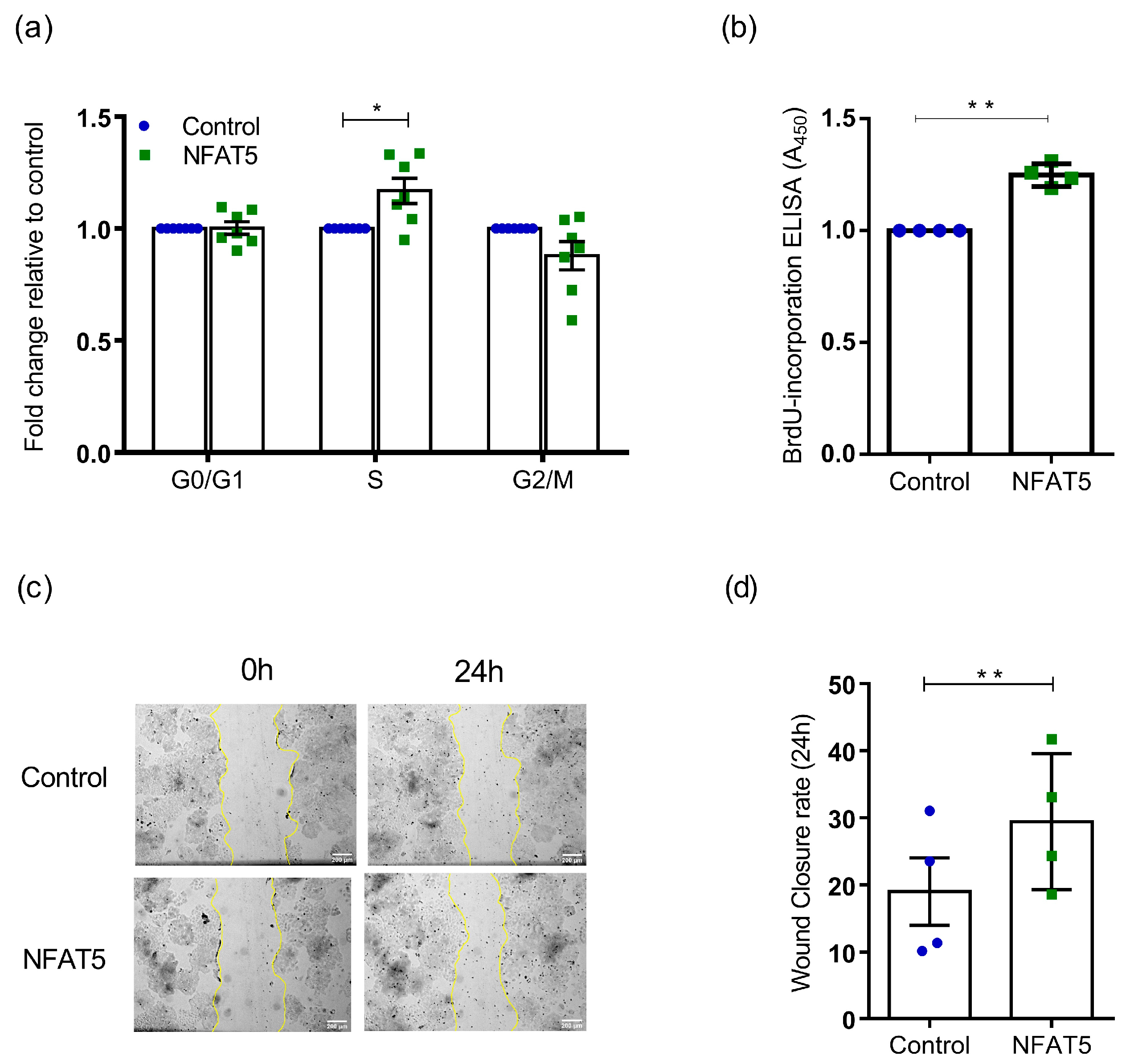
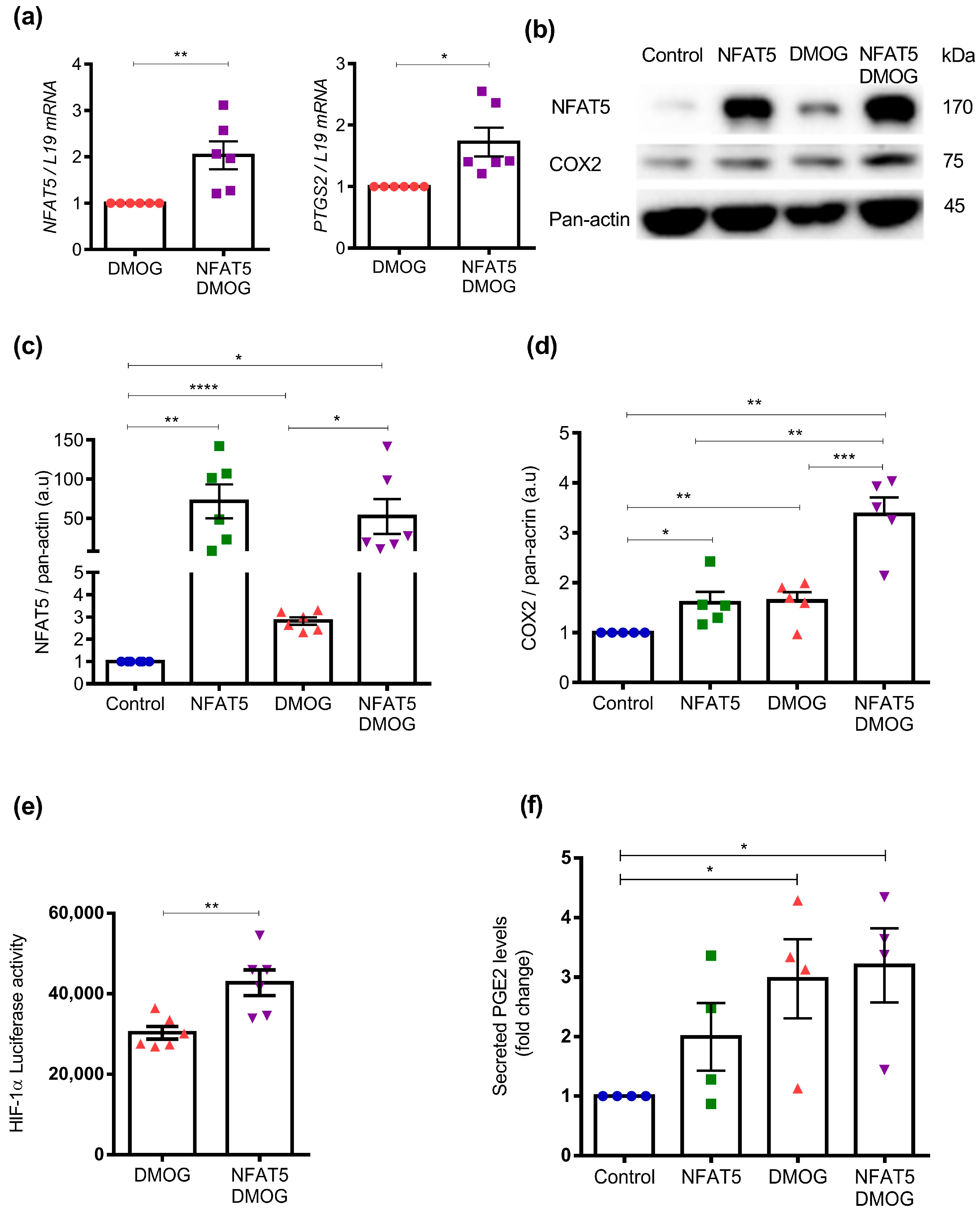
2. Results
3. Discussion
4. Materials and Methods
4.1. Clinical Sample Collection
4.2. Immunohistochemistry
4.3. Cell Culture
4.4. Plasmid DNA Transfection
4.5. RNA Sequencing and Data Analysis
4.6. Ingenuity Pathway Analysis (IPA)
4.7. Messenger RNA (mRNA) Extraction and Quantitative Real-Time Reverse Transcriptase PCR (qRT-PCR)
4.8. Western Blotting
4.9. Luciferase Reporter Assay
4.10. Enzyme-Linked Immunosorbent Assay (ELISA)
4.11. Cell-Cycle Anaylsis with Flow Cytometry
4.12. BrdU ELISA Cell Profileration Assay
4.13. Wound Scratch Assay
4.14. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raglan, O.; Kalliala, I.; Markozannes, G.; Cividini, S.; Gunter, M.J.; Nautiyal, J.; Gabra, H.; Paraskevaidis, E.; Martin-Hirsch, P.; Tsilidis, K.K.; et al. Risk factors for endometrial cancer: An umbrella review of the literature. Int. J. Cancer 2019, 145, 1719–1730. [Google Scholar] [CrossRef] [PubMed]
- Crosbie, E.J.; Kitson, S.J.; McAlpine, J.N.; Mukhopadhyay, A.; Powell, M.E.; Singh, N. Endometrial cancer. Lancet 2022, 399, 1412–1428. [Google Scholar] [CrossRef] [PubMed]
- Leone Roberti Maggiore, U.; Spanò Bascio, L.; Alboni, C.; Chiarello, G.; Savelli, L.; Bogani, G.; Martinelli, F.; Chiappa, V.; Ditto, A.; Raspagliesi, F. Sentinel lymph node biopsy in endometrial cancer: When, how and in which patients. Eur. J. Surg. Oncol. 2024, 50, 107956. [Google Scholar] [CrossRef] [PubMed]
- Makker, V.; MacKay, H.; Ray-Coquard, I.; Levine, D.A.; Westin, S.N.; Aoki, D.; Oaknin, A. Endometrial cancer. Nat. Rev. Dis. Primers 2021, 7, 88. [Google Scholar] [CrossRef] [PubMed]
- Charo, L.M.; Plaxe, S.C. Recent advances in endometrial cancer: A review of key clinical trials from 2015 to 2019. F1000Research 2019, 8, 849. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Sun, W.; Liu, H.; Zhang, D. Age at Menopause and Risk of Developing Endometrial Cancer: A Meta-Analysis. BioMed Res. Int. 2019, 2019, 8584130. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef]
- Son, J.; Carr, C.; Yao, M.; Radeva, M.; Priyadarshini, A.; Marquard, J.; Michener, C.M.; AlHilli, M. Endometrial cancer in young women: Prognostic factors and treatment outcomes in women aged ≤40 years. Int. J. Gynecol. Cancer 2020, 30, 631–639. [Google Scholar] [CrossRef]
- Wong, A.; Ngeow, J. Hereditary Syndromes Manifesting as Endometrial Carcinoma: How Can Pathological Features Aid Risk Assessment? BioMed Res. Int. 2015, 2015, 219012. [Google Scholar] [CrossRef] [PubMed]
- Onstad, M.A.; Schmandt, R.E.; Lu, K.H. Addressing the Role of Obesity in Endometrial Cancer Risk, Prevention, and Treatment. J. Clin. Oncol. 2016, 34, 4225–4230. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Wang, B.; Xu, W.; Liu, K.; Gao, Y.; Guo, C.; Chen, J.; Kamal, M.A.; Yuan, C. Endometrial Cancer: Genetic, Metabolic Characteristics, Therapeutic Strategies and Nanomedicine. Curr. Med. Chem. 2021, 28, 8755–8781. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Nie, D.; Li, Z. Targeting Six Hallmarks of Cancer in Ovarian Cancer Therapy. Curr. Cancer Drug Targets 2020, 20, 853–867. [Google Scholar] [CrossRef] [PubMed]
- Terzic, M.; Aimagambetova, G.; Kunz, J.; Bapayeva, G.; Aitbayeva, B.; Terzic, S.; Laganà, A.S. Molecular Basis of Endometriosis and Endometrial Cancer: Current Knowledge and Future Perspectives. Int. J. Mol. Sci. 2021, 22, 9274. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Lee-Kwon, W.; Kwon, H.M. The evolving role of TonEBP as an immunometabolic stress protein. Nat. Rev. Nephrol. 2020, 16, 352–364. [Google Scholar] [CrossRef]
- Lee, N.; Kim, D.; Kim, W.U. Role of NFAT5 in the Immune System and Pathogenesis of Autoimmune Diseases. Front. Immunol. 2019, 10, 270. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X. How do kinases contribute to tonicity-dependent regulation of the transcription factor NFAT5? World J. Nephrol. 2016, 5, 20–32. [Google Scholar] [CrossRef]
- Ma, S.; Zhao, Y.; Lee, W.C.; Ong, L.T.; Lee, P.L.; Jiang, Z.; Oguz, G.; Niu, Z.; Liu, M.; Goh, J.Y.; et al. Hypoxia induces HIF1α-dependent epigenetic vulnerability in triple negative breast cancer to confer immune effector dysfunction and resistance to anti-PD-1 immunotherapy. Nat. Commun. 2022, 13, 4118. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, K.; Xavier, D.; Klein-Hessling, S.; Azeem, M.; Rauschenberger, T.; Murti, K.; Avots, A.; Goebeler, M.; Klein, M.; Bopp, T.; et al. NFAT5 Controls the Integrity of Epidermis. Front. Immunol. 2021, 12, 780727. [Google Scholar] [CrossRef]
- Chen, B.L.; Li, Y.; Xu, S.; Nie, Y.; Zhang, J. NFAT5 Regulated by STUB1, Facilitates Malignant Cell Survival and p38 MAPK Activation by Upregulating AQP5 in Chronic Lymphocytic Leukemia. Biochem. Genet. 2021, 59, 870–883. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.L.; Zeng, M.L.; Shao, L.; Jiang, G.T.; Cheng, J.J.; Chen, T.X.; Han, S.; Yin, J.; Liu, W.H.; He, X.H.; et al. NFAT5 and HIF-1α Coordinate to Regulate NKCC1 Expression in Hippocampal Neurons After Hypoxia-Ischemia. Front. Cell Dev. Biol. 2019, 7, 339. [Google Scholar] [CrossRef]
- Lang, F.; Guelinckx, I.; Lemetais, G.; Melander, O. Two Liters a Day Keep the Doctor Away? Considerations on the Pathophysiology of Suboptimal Fluid Intake in the Common Population. Kidney Blood Press. Res. 2017, 42, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Cen, L.; Xing, F.; Xu, L.; Cao, Y. Potential Role of Gene Regulator NFAT5 in the Pathogenesis of Diabetes Mellitus. J. Diabetes Res. 2020, 2020, 6927429. [Google Scholar] [CrossRef] [PubMed]
- Aramburu, J.; López-Rodríguez, C. Regulation of Inflammatory Functions of Macrophages and T Lymphocytes by NFAT5. Front. Immunol. 2019, 10, 535. [Google Scholar] [CrossRef] [PubMed]
- Leibrock, C.B.; Alesutan, I.; Voelkl, J.; Pakladok, T.; Michael, D.; Schleicher, E.; Kamyabi-Moghaddam, Z.; Quintanilla-Martinez, L.; Kuro-o, M.; Lang, F. NH4Cl Treatment Prevents Tissue Calcification in Klotho Deficiency. J. Am. Soc. Nephrol. 2015, 26, 2423–2433. [Google Scholar] [CrossRef] [PubMed]
- Neubert, P.; Weichselbaum, A.; Reitinger, C.; Schatz, V.; Schroder, A.; Ferdinand, J.R.; Simon, M.; Bar, A.L.; Brochhausen, C.; Gerlach, R.G.; et al. HIF1A and NFAT5 coordinate Na(+)-boosted antibacterial defense via enhanced autophagy and autolysosomal targeting. Autophagy 2019, 15, 1899–1916. [Google Scholar] [CrossRef]
- Meng, X.; Li, Z.; Zhou, S.; Xiao, S.; Yu, P. miR-194 suppresses high glucose-induced non-small cell lung cancer cell progression by targeting NFAT5. Thorac. Cancer 2019, 10, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, H.; Shi, B.; Li, N.; Xu, G.; Yan, X.; Xu, L. Exosomal MFI2-AS1 sponge miR-107 promotes non-small cell lung cancer progression through NFAT5. Cancer Cell Int. 2023, 23, 51. [Google Scholar] [CrossRef]
- Zhen, H.; Yao, Y.; Yang, H. SAFB2 Inhibits the Progression of Breast Cancer by Suppressing the Wnt/β-Catenin Signaling Pathway via NFAT5. Mol. Biotechnol. 2023, 65, 1465–1475. [Google Scholar] [CrossRef]
- Arroyo, J.A.; Teng, C.; Battaglia, F.C.; Galan, H.L. Determination of the NFAT5/TonEBP transcription factor in the human and ovine placenta. Syst. Biol. Reprod. Med. 2009, 55, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Amara, S.; Alotaibi, D.; Tiriveedhi, V. NFAT5/STAT3 interaction mediates synergism of high salt with IL-17 towards induction of VEGF-A expression in breast cancer cells. Oncol. Lett. 2016, 12, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Chernyakov, D.; Groß, A.; Fischer, A.; Bornkessel, N.; Schultheiss, C.; Gerloff, D.; Edemir, B. Loss of RANBP3L leads to transformation of renal epithelial cells towards a renal clear cell carcinoma like phenotype. J. Exp. Clin. Cancer Res. 2021, 40, 226. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Zheng, J.; Liu, X.; Xue, Y.; Shen, S.; Zhao, L.; Li, Z.; Liu, Y. Transcription Factor NFAT5 Promotes Glioblastoma Cell-driven Angiogenesis via SBF2-AS1/miR-338-3p-Mediated EGFL7 Expression Change. Front. Mol. Neurosci. 2017, 10, 301. [Google Scholar] [CrossRef] [PubMed]
- Dzhalilova, D.S.; Makarova, O.V. HIF-Dependent Mechanisms of Relationship between Hypoxia Tolerance and Tumor Development. Biochemistry 2021, 86, 1163–1180. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.; Li, Y.; Jian, L.; Yang, Y.; Zhao, L.; Wei, M. The hypoxia-driven crosstalk between tumor and tumor-associated macrophages: Mechanisms and clinical treatment strategies. Mol. Cancer 2022, 21, 177. [Google Scholar] [CrossRef] [PubMed]
- Tam, S.Y.; Wu, V.W.C.; Law, H.K.W. Hypoxia-Induced Epithelial-Mesenchymal Transition in Cancers: HIF-1α and Beyond. Front. Oncol. 2020, 10, 486. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Li, C.; Guo, T.; Chen, J.; Wang, H.T.; Wang, Y.T.; Xiao, Y.S.; Li, J.; Liu, P.; Liu, Z.S.; et al. Upregulation of DARS2 by HBV promotes hepatocarcinogenesis through the miR-30e-5p/MAPK/NFAT5 pathway. J. Exp. Clin. Cancer Res. 2017, 36, 148. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, J.; Xie, F.; Peng, J.; Wu, X. Macrophage migration inhibitory factor promotes Warburg effect via activation of the NFkappaB/HIF1alpha pathway in lung cancer. Int. J. Mol. Med. 2018, 41, 1062–1068. [Google Scholar] [CrossRef]
- Hidalgo-Estévez, A.M.; Stamatakis, K.; Jiménez-Martínez, M.; López-Pérez, R.; Fresno, M. Cyclooxygenase 2-Regulated Genes an Alternative Avenue to the Development of New Therapeutic Drugs for Colorectal Cancer. Front. Pharmacol. 2020, 11, 533. [Google Scholar] [CrossRef] [PubMed]
- Kirkby, N.S.; Chan, M.V.; Zaiss, A.K.; Garcia-Vaz, E.; Jiao, J.; Berglund, L.M.; Verdu, E.F.; Ahmetaj-Shala, B.; Wallace, J.L.; Herschman, H.R.; et al. Systematic study of constitutive cyclooxygenase-2 expression: Role of NF-κB and NFAT transcriptional pathways. Proc. Natl. Acad. Sci. USA 2016, 113, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Shukla, V.; Kaushal, J.B.; Sankhwar, P.; Manohar, M.; Dwivedi, A. Inhibition of TPPP3 attenuates β-catenin/NF-κB/COX-2 signaling in endometrial stromal cells and impairs decidualization. J. Endocrinol. 2019, 240, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Lyndin, M.; Kravtsova, O.; Sikora, K.; Lyndina, Y.; Kuzenko, Y.; Awuah, W.A.; Abdul-Rahman, T.; Hyriavenko, N.; Sikora, V.; Romaniuk, A. COX2 Effects on endometrial carcinomas progression. Pathol. Res. Pract. 2022, 238, 154082. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Jiao, S.C.; Zhang, G.Q.; Guan, Y.; Wang, J.L. Tumor-associated immune factors are associated with recurrence and metastasis in non-small cell lung cancer. Cancer Gene Ther. 2017, 24, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Piasecka, D.; Braun, M.; Mieszkowska, M.; Kowalczyk, L.; Kopczynski, J.; Kordek, R.; Sadej, R.; Romanska, H.M. Upregulation of HIF1-α via an NF-κB/COX2 pathway confers proliferative dominance of HER2-negative ductal carcinoma in situ cells in response to inflammatory stimuli. Neoplasia 2020, 22, 576–589. [Google Scholar] [CrossRef]
- Ding, Y.; Zhuang, S.; Li, Y.; Yu, X.; Lu, M.; Ding, N. Hypoxia-induced HIF1α dependent COX2 promotes ovarian cancer progress. J. Bioenerg. Biomembr. 2021, 53, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Cizkova, K.; Foltynkova, T.; Gachechiladze, M.; Tauber, Z. Comparative Analysis of Immunohistochemical Staining Intensity Determined by Light Microscopy, ImageJ and QuPath in Placental Hofbauer Cells. Acta Histochem. Cytochem. 2021, 54, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Eritja, N.; Chen, B.J.; Rodríguez-Barrueco, R.; Santacana, M.; Gatius, S.; Vidal, A.; Martí, M.D.; Ponce, J.; Bergadà, L.; Yeramian, A.; et al. Autophagy orchestrates adaptive responses to targeted therapy in endometrial cancer. Autophagy 2017, 13, 608–624. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Seike, M.; Chiba, M.; Takahashi, S.; Nakamichi, S.; Matsumoto, M.; Takeuchi, S.; Minegishi, Y.; Noro, R.; Kunugi, S.; et al. Ankyrin Repeat Domain 1 Overexpression is Associated with Common Resistance to Afatinib and Osimertinib in EGFR-mutant Lung Cancer. Sci. Rep. 2018, 8, 14896. [Google Scholar] [CrossRef] [PubMed]
- Ahmad Zawawi, S.S.; Mohd Azram, N.A.S.; Sulong, S.; Zakaria, A.D.; Lee, Y.Y.; Che Jalil, N.A.; Musa, M. Identification of AOC3 and LRRC17 as Colonic Fibroblast Activation Markers and Their Potential Roles in Colorectal Cancer Progression. Asian Pac. J. Cancer Prev. 2023, 24, 3099–3107. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.; Shi, X.; Zhang, S.; Umbach, A.T.; Chen, H.; Salker, M.S.; Lang, F. Prolyl hydroxylase 3 (PHD3) expression augments the development of regulatory T cells. Mol. Immunol. 2016, 76, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Ahmed, M.A.; Forsyth, N.R. Dimethyloxalylglycine (DMOG), a Hypoxia Mimetic Agent, Does Not Replicate a Rat Pheochromocytoma (PC12) Cell Biological Response to Reduced Oxygen Culture. Biomolecules 2022, 12, 541. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Bleiziffer, O.; Boos, A.M.; Sun, J.; Brandl, A.; Beier, J.P.; Arkudas, A.; Schmitz, M.; Kneser, U.; Horch, R.E. PHDs inhibitor DMOG promotes the vascularization process in the AV loop by HIF-1a up-regulation and the preliminary discussion on its kinetics in rat. BMC Biotechnol. 2014, 14, 112. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Shah, Y.M. Hypoxia-inducible factor-2alpha is essential in activating the COX2/mPGES-1/PGE2 signaling axis in colon cancer. Carcinogenesis 2013, 34, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Serman, Y.; Fuentealba, R.A.; Pasten, C.; Rocco, J.; Ko, B.C.B.; Carrion, F.; Irarrazabal, C.E. Emerging new role of NFAT5 in inducible nitric oxide synthase in response to hypoxia in mouse embryonic fibroblast cells. Am. J. Physiol. Cell Physiol. 2019, 317, C31–C38. [Google Scholar] [CrossRef] [PubMed]
- Sahu, I.; Pelzl, L.; Sukkar, B.; Fakhri, H.; Al-Maghout, T.; Cao, H.; Hauser, S.; Gutti, R.; Gawaz, M.; Lang, F. NFAT5-sensitive Orai1 expression and store-operated Ca2+ entry in megakaryocytes. FASEB J. 2017, 31, 3439–3448. [Google Scholar] [CrossRef] [PubMed]
- Maouyo, D.; Kim, J.Y.; Lee, S.D.; Wu, Y.; Woo, S.K.; Kwon, H.M. Mouse TonEBP-NFAT5: Expression in early development and alternative splicing. Am. J. Physiol. Renal Physiol. 2002, 282, F802–F809. [Google Scholar] [CrossRef]
- Packialakshmi, B.; Hira, S.; Lund, K.; Zhang, A.H.; Halterman, J.; Feng, Y.; Scott, D.W.; Lees, J.R.; Zhou, X. NFAT5 contributes to the pathogenesis of experimental autoimmune encephalomyelitis (EAE) and decrease of T regulatory cells in female mice. Cell. Immunol. 2022, 375, 104515. [Google Scholar] [CrossRef]
- Lee, H.H.; Jeong, G.W.; Ye, B.J.; Yoo, E.J.; Son, K.S.; Kim, D.K.; Park, H.K.; Kang, B.H.; Lee-Kwon, W.; Kwon, H.M.; et al. TonEBP in Myeloid Cells Promotes Obesity-Induced Insulin Resistance and Inflammation Through Adipose Tissue Remodeling. Diabetes 2022, 71, 2557–2571. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Zhang, M.; Zhao, M.; Davis, L.S.; Blackwell, T.S.; Yull, F.; Breyer, M.D.; Hao, C.M. Increased dietary sodium induces COX2 expression by activating NFkappaB in renal medullary interstitial cells. Pflugers Arch. 2014, 466, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Alauddin, M.; Okumura, T.; Rajaxavier, J.; Khozooei, S.; Pöschel, S.; Takeda, S.; Singh, Y.; Brucker, S.Y.; Wallwiener, D.; Koch, A.; et al. Gut Bacterial Metabolite Urolithin A Decreases Actin Polymerization and Migration in Cancer Cells. Mol. Nutr. Food Res. 2020, 64, 1900390. [Google Scholar] [CrossRef] [PubMed]
- Johnson, Z.I.; Doolittle, A.C.; Snuggs, J.W.; Shapiro, I.M.; Le Maitre, C.L.; Risbud, M.V. TNF-α promotes nuclear enrichment of the transcription factor TonEBP/NFAT5 to selectively control inflammatory but not osmoregulatory responses in nucleus pulposus cells. J. Biol. Chem. 2017, 292, 17561–17575. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Qu, B.; Li, Y.M.; Yang, L.B.; Fan, K.X.; Zheng, H.; Huang, H.D.; Gu, J.W.; Kuang, Y.Q.; Ma, Y. NFAT5 protects astrocytes against oxygen-glucose-serum deprivation/restoration damage via the SIRT1/Nrf2 pathway. J. Mol. Neurosci. 2017, 61, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-T.; Hung, W.-C.; Kang, W.-Y.; Huang, Y.-C.; Su, Y.-C.; Yang, C.-H.; Chai, C.-Y. Overexpression of cyclooxygenase-2 in urothelial carcinoma in conjunction with tumor-associated-macrophage infiltration, hypoxia-inducible factor-1α expression, and tumor angiogenesis. APMIS 2009, 117, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Blando, J.M.; Perez, C.J.; Lal, P.; Feldman, M.D.; Smyth, E.M.; Ricciotti, E.; Grosser, T.; Benavides, F.; Kazanietz, M.G. COX-2 mediates pro-tumorigenic effects of PKCε in prostate cancer. Oncogene 2018, 37, 4735–4749. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Comaills, V.; Burr, R.; Boulay, G.; Miyamoto, D.T.; Wittner, B.S.; Emmons, E.; Sil, S.; Koulopoulos, M.W.; Broderick, K.T.; et al. COX-2 mediates tumor-stromal prolactin signaling to initiate tumorigenesis. Proc. Natl. Acad. Sci. USA 2019, 116, 5223–5232. [Google Scholar] [CrossRef] [PubMed]
- Alexanian, A.; Sorokin, A. Cyclooxygenase 2: Protein-protein interactions and posttranslational modifications. Physiol. Genom. 2017, 49, 667–681. [Google Scholar] [CrossRef] [PubMed]
- Greenhough, A.; Smartt, H.J.; Moore, A.E.; Roberts, H.R.; Williams, A.C.; Paraskeva, C.; Kaidi, A. The COX-2/PGE2 pathway: Key roles in the hallmarks of cancer and adaptation to the tumour microenvironment. Carcinogenesis 2009, 30, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Nasry, W.H.S.; Rodriguez-Lecompte, J.C.; Martin, C.K. Role of COX-2/PGE2 Mediated Inflammation in Oral Squamous Cell Carcinoma. Cancers 2018, 10, 348. [Google Scholar] [CrossRef] [PubMed]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, A.S.; Budac, D.; Khayrullina, T.; Staal, R.; Chandrasena, G. Quantitative analysis of lipids: A higher-throughput LC-MS/MS-based method and its comparison to ELISA. Future Sci. OA 2017, 3, Fso157. [Google Scholar] [CrossRef]
- Kefayat, A.; Ghahremani, F.; Safavi, A.; Hajiaghababa, A.; Moshtaghian, J. C-phycocyanin: A natural product with radiosensitizing property for enhancement of colon cancer radiation therapy efficacy through inhibition of COX-2 expression. Sci. Rep. 2019, 9, 19161. [Google Scholar] [CrossRef] [PubMed]
- Li, X.H.; Li, J.J.; Zhang, H.W.; Sun, P.; Zhang, Y.L.; Cai, S.H.; Ren, X.D. Nimesulide inhibits tumor growth in mice implanted hepatoma: Overexpression of Bax over Bcl-2. Acta Pharmacol. Sin. 2003, 24, 1045–1050. [Google Scholar]
- Gui, D.; Li, Y.; Chen, X.; Gao, D.; Yang, Y.; Li, X. HIF1 signaling pathway involving iNOS, COX2 and caspase9 mediates the neuroprotection provided by erythropoietin in the retina of chronic ocular hypertension rats. Mol. Med. Rep. 2015, 11, 1490–1496. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Grigsby, C.L.; Law, C.S.; Ni, X.; Nekrep, N.; Olsen, K.; Humphreys, M.H.; Gardner, D.G. Tonicity-dependent induction of Sgk1 expression has a potential role in dehydration-induced natriuresis in rodents. J. Clin. Investig. 2009, 119, 1647–1658. [Google Scholar] [CrossRef] [PubMed]
- Lang, F.; Shumilina, E. Regulation of ion channels by the serum- and glucocorticoid-inducible kinase SGK1. FASEB J. 2013, 27, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Roche, J. The Epithelial-to-Mesenchymal Transition in Cancer. Cancers 2018, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Glubb, D.M.; O’Mara, T.A. Dietary Factors and Endometrial Cancer Risk: A Mendelian Randomization Study. Nutrients 2023, 15, 603. [Google Scholar] [CrossRef] [PubMed]
- Balan, Y.; Packirisamy, R.M.; Mohanraj, P.S. High dietary salt intake activates inflammatory cascades via Th17 immune cells: Impact on health and diseases. Arch. Med. Sci. 2022, 18, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Li, G.; Jiang, X.; Shen, X.; Li, H.; Yang, L.; Liu, W. NFAT5 directs hyperosmotic stress-induced fibrin deposition and macrophage infiltration via PAI-1 in endothelium. Aging 2020, 13, 3661–3679. [Google Scholar] [CrossRef]
- Schröder, A.; Leikam, A.; Käppler, P.; Neubert, P.; Jantsch, J.; Neuhofer, W.; Deschner, J.; Proff, P.; Kirschneck, C. Impact of salt and the osmoprotective transcription factor NFAT-5 on macrophages during mechanical strain. Immunol. Cell Biol. 2021, 99, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Madonna, R.; Giovannelli, G.; Confalone, P.; Renna, F.V.; Geng, Y.J.; De Caterina, R. High glucose-induced hyperosmolarity contributes to COX-2 expression and angiogenesis: Implications for diabetic retinopathy. Cardiovasc. Diabetol. 2016, 15, 18. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ochoa, E.O.; Robison, P.; Contreras, M.; Shen, T.; Zhao, Z.; Schneider, M.F. Elevated extracellular glucose and uncontrolled type 1 diabetes enhance NFAT5 signaling and disrupt the transverse tubular network in mouse skeletal muscle. Exp. Biol. Med. 2012, 237, 1068–1083. [Google Scholar] [CrossRef]
- Padmanabhan, S.; Caulfield, M.; Dominiczak, A.F. Genetic and Molecular Aspects of Hypertension. Circ. Res. 2015, 116, 937–959. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef]
- Chen, Y.; Lun, A.T.; Smyth, G.K. From reads to genes to pathways: Differential expression analysis of RNA-Seq experiments using Rsubread and the edgeR quasi-likelihood pipeline. F1000Research 2016, 5, 1438. [Google Scholar] [CrossRef]
- Chicco, D.; Jurman, G. A brief survey of tools for genomic regions enrichment analysis. Front. Bioinform. 2022, 2, 968327. [Google Scholar] [CrossRef]
- Thomas, P.D.; Ebert, D.; Muruganujan, A.; Mushayahama, T.; Albou, L.P.; Mi, H. PANTHER: Making genome-scale phylogenetics accessible to all. Protein Sci. 2022, 31, 8–22. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Fabregat, A.; Sidiropoulos, K.; Viteri, G.; Forner, O.; Marin-Garcia, P.; Arnau, V.; D’Eustachio, P.; Stein, L.; Hermjakob, H. Reactome pathway analysis: A high-performance in-memory approach. BMC Bioinform. 2017, 18, 142. [Google Scholar] [CrossRef]
- Schriml, L.M.; Mitraka, E.; Munro, J.; Tauber, B.; Schor, M.; Nickle, L.; Felix, V.; Jeng, L.; Bearer, C.; Lichenstein, R.; et al. Human Disease Ontology 2018 update: Classification, content and workflow expansion. Nucleic Acids Res. 2019, 47, D955–D962. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ayakannu, T.; Taylor, A.H.; Willets, J.M.; Brown, L.; Lambert, D.G.; McDonald, J.; Davies, Q.; Moss, E.L.; Konje, J.C. Validation of endogenous control reference genes for normalizing gene expression studies in endometrial carcinoma. Mol. Hum. Reprod. 2015, 21, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Salker, M.S.; Singh, Y.; Durairaj, R.R.P.; Yan, J.; Alauddin, M.; Zeng, N.; Steel, J.H.; Zhang, S.; Nautiyal, J.; Webster, Z.; et al. LEFTY2 inhibits endometrial receptivity by downregulating Orai1 expression and store-operated Ca2+ entry. J. Mol. Med. 2018, 96, 173–182. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Characteristics | Total | NFAT5 Score | p-Value | ||
|---|---|---|---|---|---|
| 1 | 2 | 3 | |||
| Age > 60 years | 15 (100%) | 5(33.3%) | 6 (40%) | 4 (26.7%) | p = 0.941 |
| Age < 60 years | 11 (100%) | 3 (27.3%) | 5 (45.5%) | 3 (27.3%) | |
| Premenopausal | 7 (100%) | 3 (42.9%) | 3 (42.9%) | 1 (14.3%) | |
| Postmenopausal | 19 (100%) | 5 (26.3%) | 8 (42.1%) | 6 (31.6%) | p = 0.599 |
| Endometriod | 24 (100%) | 8 (33.3%) | 9 (37.5%) | 7 (29.2%) | p = 0.228 |
| Serous | 2 (100%) | 0 (0%) | 2 (100%) | 0 (0%) | |
| Grade 1/2 | 15 (100%) | 8 (53.3%) | 7 (46.7%) | 0 (0%) | p < 0.001 |
| Grade 3 | 11 (100%) | 0 (0%) | 4 (36.4%) | 7 (63.6%) | |
| pT1a | 15 (100%) | 7 (46.7%) | 7 (46.7%) | 1 (6.7%) | p = 0.091 |
| pT1b | 6 (100%) | 0 (0%) | 3 (50%) | 3 (50%) | |
| pT2 | 2 (100%) | 1 (50%) | 0 (0%) | 1 (50%) | |
| pT3a | 3 (100%) | 0 (0%) | 1 (33.3%) | 2 (66.7%) | |
| pT1a | 15 (100%) | 7 (46.7%) | 7 (46.7%) | 1 (6.7%) | p = 0.014 |
| >pT1b | 11 (100%) | 1 (9.1%) | 4 (36.4%) | 6 (54.5%) | |
| Regional Nodes pN0 | 21 (100%) | 7 (33.3%) | 10 (47.6%) | 4 (19%) | p = 0.176 |
| Regional Nodes pN1 | 5 (100%) | 1 (20%) | 1 (20%) | 3 (60%) | |
| Metastatsis 0 | 20 (100%) | 7 (35%) | 10 (50%) | 3 (15%) | p = 0.043 |
| Metastatsis 1 | 6 (100%) | 1 (16.7%) | 1 (16.7%) | 4 (66.7%) | |
| Lymph Vessel L0 | 19 (100%) | 8 (42.1%) | 7 (36.8%) | 4 (21.1%) | p = 0.114 |
| Lymph Vessel L1 | 7 (100%) | 0 (0%) | 4 (57.1%) | 3 (42.9%) | |
| Gene | Primer Sequence |
|---|---|
| L19 | Forward (5′-3′): GCGGAAGGGTACAGCCAA |
| Reverse (5′-3′): GCAGCCGGCGCAAA | |
| NFAT5 | Forward (5′-3′): GAGCAGAGCTGCAGTAT |
| Reverse (5′-3′): AGCTGAGAAAGCACATAG | |
| PTGS2 | Forward (5′-3′): GCTCAAACATGATGTTTGCATTC |
| Reverse (5′-3′): GCTGGCCCTCGCTTATGA | |
| HIF1A | Forward (5′-3′): TCTGGACTTGCCTTTCCTTCTC |
| Reverse (5′-3′): AACTTATCTTTTTCTTGTCGTTCGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okumura, T.; Raja Xavier, J.P.; Pasternak, J.; Yang, Z.; Hang, C.; Nosirov, B.; Singh, Y.; Admard, J.; Brucker, S.Y.; Kommoss, S.; et al. Rel Family Transcription Factor NFAT5 Upregulates COX2 via HIF-1α Activity in Ishikawa and HEC1a Cells. Int. J. Mol. Sci. 2024, 25, 3666. https://doi.org/10.3390/ijms25073666
Okumura T, Raja Xavier JP, Pasternak J, Yang Z, Hang C, Nosirov B, Singh Y, Admard J, Brucker SY, Kommoss S, et al. Rel Family Transcription Factor NFAT5 Upregulates COX2 via HIF-1α Activity in Ishikawa and HEC1a Cells. International Journal of Molecular Sciences. 2024; 25(7):3666. https://doi.org/10.3390/ijms25073666
Chicago/Turabian StyleOkumura, Toshiyuki, Janet P. Raja Xavier, Jana Pasternak, Zhiqi Yang, Cao Hang, Bakhtiyor Nosirov, Yogesh Singh, Jakob Admard, Sara Y. Brucker, Stefan Kommoss, and et al. 2024. "Rel Family Transcription Factor NFAT5 Upregulates COX2 via HIF-1α Activity in Ishikawa and HEC1a Cells" International Journal of Molecular Sciences 25, no. 7: 3666. https://doi.org/10.3390/ijms25073666