
A Novel Model of Staphylococcus aureus-Induced Lymphoplasmacytic Rhinosinusitis in Rats

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. S. aureus Clinical Isolate Selection
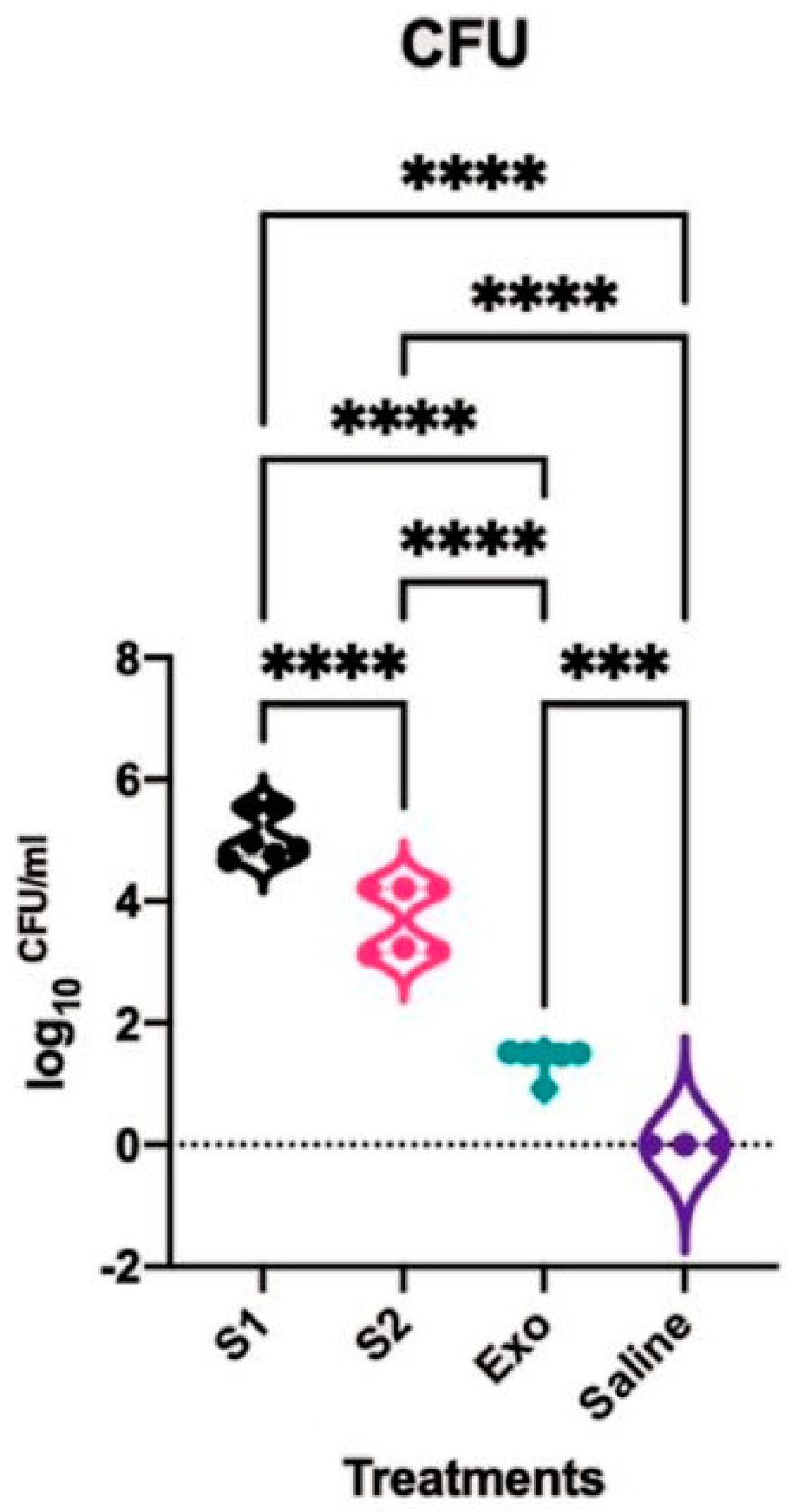
2.2. Rat Nasal Cavity CFU Count
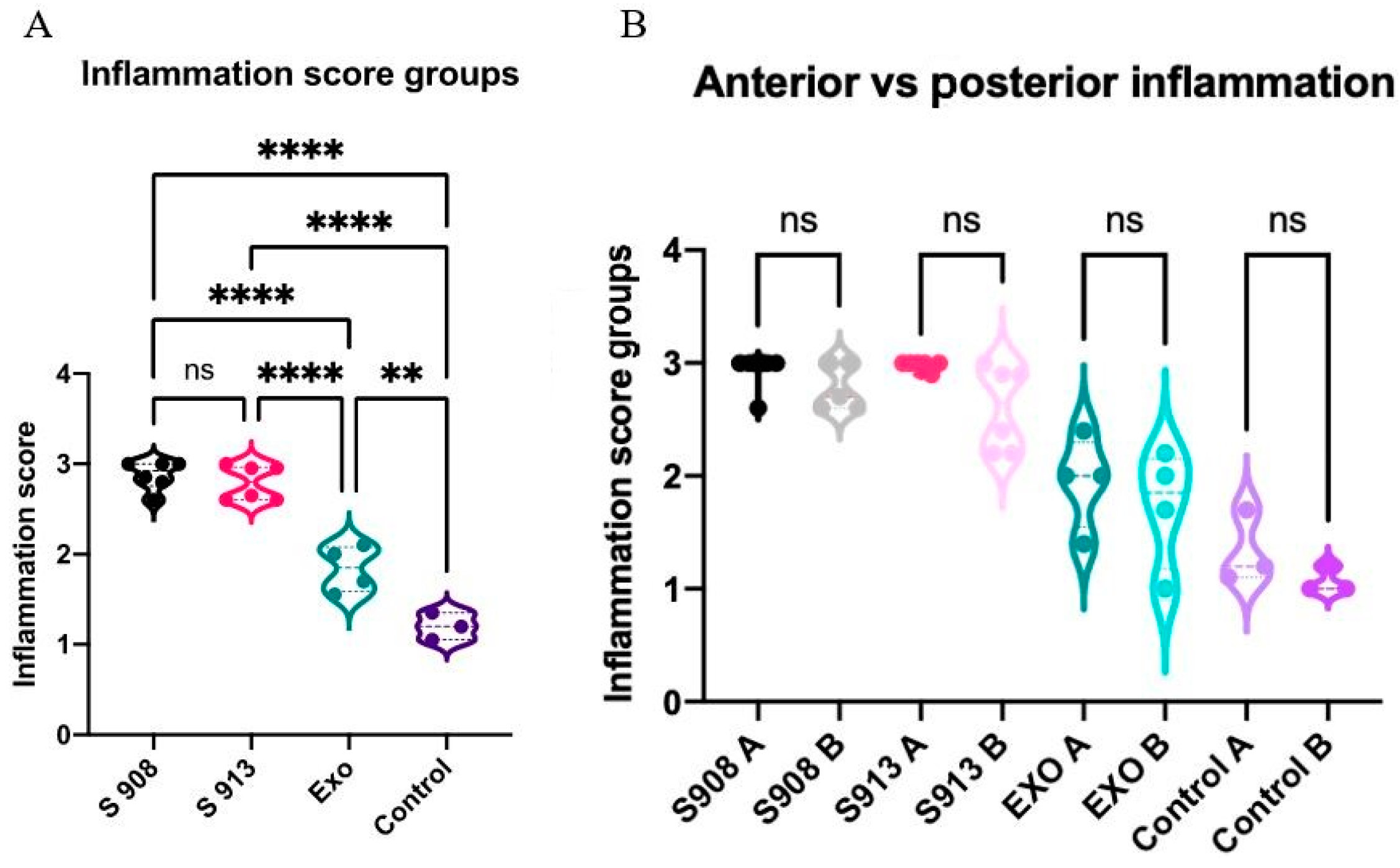
2.3. S. aureus Clinical Isolates and Exoproteins Induced Significant Inflammation
2.4. S. aureus Invades the Nasal Mucosa
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Staphylococcus aureus Clinical Isolates Cultured from Patients
4.3. Staphylococcus aureus Biofilm Exoprotein Preparation
4.4. Bradford Protein Assay
4.5. Selection and Preparation of S. aureus Clinical Isolates for Inoculation
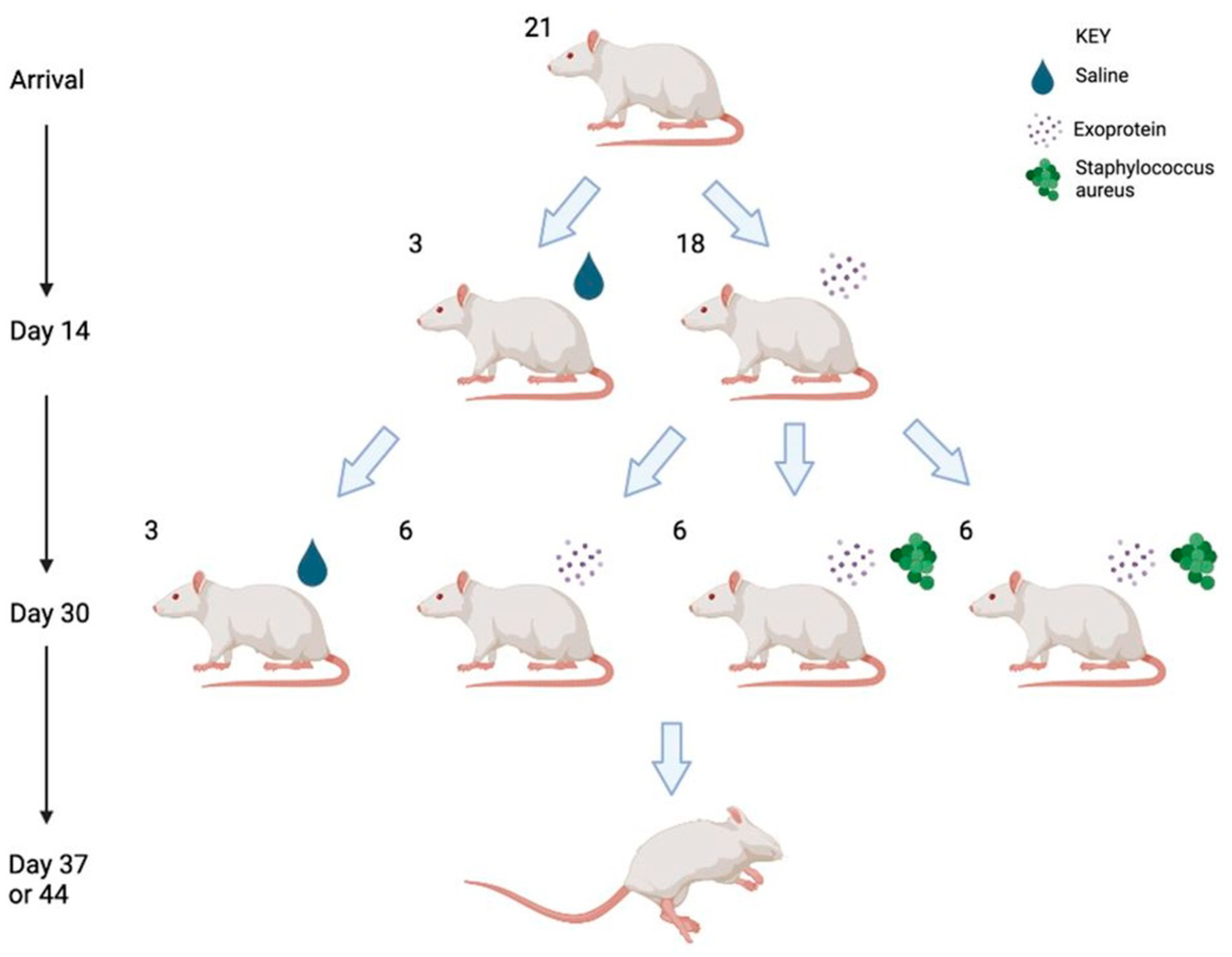
4.6. Rhinosinusitis Rat Model
4.7. CFU Counts
4.8. Histopathology Examination with Haematoxylin and Eosin and Gram Staining
4.9. Genomic DNA Extraction and Sequencing
4.10. Bioinformatics
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benninger, M.S.; Ferguson, B.J.; Hadley, J.A.; Hamilos, D.L.; Jacobs, M.; Kennedy, D.W.; Lanza, D.C.; Marple, B.F.; Osguthorpe, J.D.; Stankiewicz, J.A.; et al. Adult Chronic Rhinosinusitis: Definitions, Diagnosis, Epidemiology, and Pathophysiology. Otolaryngol. Head Neck Surg. 2003, 129, S1–S32. [Google Scholar] [CrossRef]
- Lee, S.; Lane, A.P. Chronic rhinosinusitis as a multifactorial inflammatory disorder. Curr. Infect. Dis. Rep. 2011, 13, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Vlaminck, S.; Acke, F.; Scadding, G.K.; Lambrecht, B.N.; Gevaert, P. Pathophysiological and clinical aspects of chronic rhinosinusitis: Current concepts. Front. Allergy 2021, 2, 741788. [Google Scholar] [CrossRef]
- Van Crombruggen, K.; Zhang, N.; Gevaert, P.; Tomassen, P.; Bachert, C. Pathogenesis of chronic rhinosinusitis: Inflammation. J. Allergy Clin. Immunol. 2011, 128, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Habib, A.-R.; Campbell, R.; Kalish, L.; Wong, E.H.; Grayson, J.; Alvarado, R.; Sacks, R.; Harvey, R.J. The burden of chronic upper airway disorders in Australia: A population-based cross-sectional study. Aust. J. Otolaryngol. 2019, 2, 28. [Google Scholar] [CrossRef]
- Vickery, T.W.; Ramakrishnan, V.R.; Suh, J.D. The Role of Staphylococcus aureus in Patients with Chronic Sinusitis and Nasal Polyposis. Curr. Allergy Asthma Rep. 2019, 19, 21. [Google Scholar] [CrossRef]
- Van Staa, T.P.; Travis, S.; Leufkens, H.G.; Logan, R.F. 5-aminosalicylic acids and the risk of renal disease: A large British epidemiologic study. Gastroenterology 2004, 126, 1733–1739. [Google Scholar] [CrossRef] [PubMed]
- Okifo, O.; Ray, A.; Gudis, D.A. The Microbiology of Acute Exacerbations in Chronic Rhinosinusitis—A Systematic Review. Front. Cell. Infect. Microbiol. 2022, 12, 858196. [Google Scholar] [CrossRef] [PubMed]
- Fokkens, W.J.; Lund, V.J.; Hopkins, C.; Hellings, P.W.; Kern, R.; Reitsma, S.; Toppila-Salmi, S.; Bernal-Sprekelsen, M.; Mullol, J.; Alobid, I.; et al. European position paper on rhinosinusitis and nasal polyps 2020. Rhinology 2020, 58, 1–464. [Google Scholar] [CrossRef]
- Al-Sayed, A.A.; Agu, R.U.; Massoud, E. Models for the study of nasal and sinus physiology in health and disease: A review of the literature. Laryngoscope Investig. Otolaryngol. 2017, 2, 398–409. [Google Scholar] [CrossRef]
- Chow, A.W.; Benninger, M.S.; Brook, I.; Brozek, J.L.; Goldstein, E.J.; Hicks, L.A.; Pankey, G.A.; Seleznick, M.; Volturo, G.; Wald, E.R. IDSA clinical practice guideline for acute bacterial rhinosinusitis in children and adults. Clin. Infect. Dis. 2012, 54, e72–e112. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.; Meng, Y.; Piao, Y.; Zhang, N.; Bachert, C.; Wang, C.; Zhang, L. Cellular phenotyping of chronic rhinosinusitis with nasal polyps. Rhinology 2016, 54, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Liu, W.; Zhang, L.; Bai, J.; Fan, Y.; Xia, W.; Luo, Q.; Zheng, J.; Wang, H.; Li, Z.; et al. Increased neutrophilia in nasal polyps reduces the response to oral corticosteroid therapy. J. Allergy Clin. Immunol. 2012, 129, 1522–1528.e5. [Google Scholar] [CrossRef] [PubMed]
- Kirtsreesakul, V.; Atchariyasathian, V. Nasal Polyposis: Role of Allergy on Therapeutic Response of Eosinophil- and Noneosinophil-Dominated Inflammation. Am. J. Rhinol. 2006, 20, 95–100. [Google Scholar] [CrossRef]
- Ho, J.; Alvarado, R.; Rimmer, J.; Sewell, W.A.; Walter, S.; Earls, P.; Campbell, R.G.; Sacks, R.; Kalish, L.H.; Harvey, R.J. Comparison of Sinonasal Histopathological Changes in Biological Treatment of Eosinophilic Chronic Rhinosinusitis. Am. J. Rhinol. Allergy 2021, 36, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Huang, Y.; Delemarre, T.; Cavaliere, C.; Zhang, N.; Bachert, C. Advances in chronic rhinosinusitis in 2020 and 2021. J. Allergy Clin. Immunol. 2021, 149, 854–866. [Google Scholar] [CrossRef] [PubMed]
- Barré-Sinoussi, F.; Montagutelli, X.; MacLeod, R.; Hillert, E.-K.; Cameron, R.T.; Baillie, G.S.; Caffrey, C.R.; White, J.T.; Crossman, M.; Subramanyam, M. Animal models are essential to biological research: Issues and perspectives. Futur. Sci. OA 2015, 1, FSO63. [Google Scholar] [CrossRef]
- Hilding, A. Experimental sinus surgery. Effects of operative windows on nasal sinuses. Ann. Otol. Rhinol. Laryngol. 1933, 17, 321–327. [Google Scholar]
- Hoggard, M.; Biswas, K.; Zoing, M.; Mackenzie, B.W.; Taylor, M.W.; Douglas, R.G. Evidence of microbiota dysbiosis in chronic rhinosinusitis. Int. Forum Allergy Rhinol. 2016, 7, 230–239. [Google Scholar] [CrossRef]
- Lux, C.; Douglas, R.; Cho, D.-Y.; Taylor, M.; Biswas, K. Animal models for inflammatory mucosal disease and their potential for studying the microbiome in chronic rhinosinusitis. Rhinol. Online 2019, 2, 69–80. [Google Scholar] [CrossRef]
- Mapara, M.; Thomas, B.S.; Bhat, K.M. Rabbit as an animal model for experimental research. Dent. Res. J. 2012, 9, 111–118. [Google Scholar] [CrossRef]
- Lindsay, R.; Slaughter, T.; Britton-Webb, J.; Mog, S.R.; Conran, R.; Tadros, M.; Earl, N.; Fox, D.; Roberts, J.; Bolger, W.E. Development of a Murine Model of Chronic Rhinosinusitis. Otolaryngol. Neck Surg. 2006, 134, 724–730. [Google Scholar] [CrossRef]
- Lavelle, G.M.; White, M.M.; Browne, N.; McElvaney, N.G.; Reeves, E.P. Animal Models of Cystic Fibrosis Pathology: Phenotypic Parallels and Divergences. BioMed Res. Int. 2016, 2016, 525872. [Google Scholar] [CrossRef]
- Marks, S.C. Acute Sinusitis in the Rabbit: A New Rhinogenic Model. Laryngoscope 1997, 107, 1579–1585. [Google Scholar] [CrossRef] [PubMed]
- Ha, K.R.; Psaltis, A.J.; Tan, L.; Wormald, P.-J. A Sheep Model for the Study of Biofilms in Rhinosinusitis. Am. J. Rhinol. 2007, 21, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Boase, S.; Jervis-Bardy, J.; Cleland, E.; Pant, H.; Tan, L.; Wormald, P. Bacterial-induced epithelial damage promotes fungal biofilm formation in a sheep model of sinusitis. Int. Forum Allergy Rhinol. 2013, 3, 341–348. [Google Scholar] [CrossRef]
- Boase, S.; Valentine, R.; Singhal, D.; Tan, L.W.; Wormald, P. A sheep model to investigate the role of fungal biofilms in sinusitis: Fungal and bacterial synergy. Int. Forum Allergy Rhinol. 2011, 1, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Smolich, J.J.; Stratford, B.F.; Maloney, J.E.; Ritchie, B.C. New features in the development of the submucosal gland of the respiratory tract. J. Anat. 1978, 127, 223–238. [Google Scholar]
- Psaltis, A.J.; Wormald, P.-J. Therapy of Sinonasal Microbiome in CRS: A Critical Approach. Curr. Allergy Asthma Rep. 2017, 17, 59. [Google Scholar] [CrossRef]
- Mackenzie, B.W.; Waite, D.W.; Hoggard, M.; Douglas, R.G.; Taylor, M.W.; Biswas, K. Bacterial community collapse: A meta-analysis of the sinonasal microbiota in chronic rhinosinusitis. Environ. Microbiol. 2017, 19, 381–392. [Google Scholar] [CrossRef]
- Jia, M.; Chen, Z.; Du, X.; Guo, Y.; Sun, T.; Zhao, X. A Simple Animal Model of Staphylococcus Aureus Biofilm in Sinusitis. Am. J. Rhinol. Allergy 2014, 28, e115–e119. [Google Scholar] [CrossRef] [PubMed]
- Redington, A.E. Fibrosis and airway remodelling. Clin. Exp. Allergy 2000, 30, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.A.; Bercik, P. Macrophages are related to goblet cell hyperplasia and induce MUC5B but not MUC5AC in human bronchus epithelial cells. Mod. Pathol. 2012, 92, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Spurzem, J.R.; Thompson, A.B.; Daughton, D.M.; Mueller, M.; Linder, J.; Rennard, S.I. Chronic Inflammation is Associated with an Increased Proportion of Goblet Cells Recovered by Bronchial Lavage. Chest 1991, 100, 389–393. [Google Scholar] [CrossRef]
- Ramadan, H.H.; Hinerman, R.A. Smoke Exposure and Outcome of Endoscopic Sinus Surgery in Children. Otolaryngol. Neck Surg. 2002, 127, 546–548. [Google Scholar] [CrossRef]
- Sethi, S. Bacterial Infection and the Pathogenesis of COPD. Chest 2000, 117, 286S–291S. [Google Scholar] [CrossRef]
- Marino, M.J.; Garcia, J.O.; Zarka, M.A.; Lal, D. Inflammatory cell predominance and patterns in chronic rhinosinusitis with and without nasal polyposis patients. Laryngoscope Investig. Otolaryngol. 2019, 4, 573–577. [Google Scholar] [CrossRef]
- Khonsary, S. Guyton and Hall: Textbook of Medical Physiology. Surg. Neurol. Int. 2017, 8, 275. [Google Scholar] [CrossRef]
- Tsybikov, N.N.; Egorova, E.V.; Kuznik, B.I.; Fefelova, E.V.; Magen, E. Anticytokine autoantibodies in chronic rhinosinusitis. Allergy Asthma Proc. 2015, 36, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, N.; Zheng, M.; Li, Y.; Meng, L.; Ruan, Y.; Han, J.; Zhao, N.; Wang, X.; Zhang, L.; et al. Cross-talk between TH2 and TH17 pathways in patients with chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 2019, 144, 1254–1264. [Google Scholar] [CrossRef] [PubMed]
- Hulse, K.E.; Norton, J.E.; Suh, L.; Zhong, Q.; Mahdavinia, M.; Simon, P.; Kern, R.C.; Conley, D.B.; Chandra, R.K.; Tan, B.K.; et al. Chronic rhinosinusitis with nasal polyps is characterized by B-cell inflammation and EBV-induced protein 2 expression. J. Allergy Clin. Immunol. 2013, 131, 1075–1083.e7. [Google Scholar] [CrossRef] [PubMed]
- Feldman, S.; Kasjanski, R.; Poposki, J.; Hernandez, D.; Chen, J.N.; Norton, J.E.; Suh, L.; Carter, R.G.; Stevens, W.W.; Peters, A.T.; et al. Chronic airway inflammation provides a unique environment for B cell activation and antibody production. Clin. Exp. Allergy 2016, 47, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Jia, L.; Bai, L.; He, L.; Yang, B.; Wu, C.; Li, H. Phenotypic and functional characteristics of IL-21-expressing CD8+ T cells in human nasal polyps. Sci. Rep. 2016, 6, 30362. [Google Scholar] [CrossRef] [PubMed]
- Gevaert, P.; Holtappels, G.; Johansson, S.G.O.; Cuvelier, C.; van Cauwenberge, P.; Bachert, C. Organization of secondary lymphoid tissue and local IgE formation to Staphylococcus aureus enterotoxins in nasal polyp tissue. Allergy 2004, 60, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Lau, A.; Lester, S.; Moraitis, S.; Ou, J.; Psaltis, A.J.; McColl, S.; Rischmueller, M.; Wormald, P.-J.; Vreugde, S. Tertiary lymphoid organs in recalcitrant chronic rhinosinusitis. J. Allergy Clin. Immunol. 2017, 139, 1371–1373.e6. [Google Scholar] [CrossRef] [PubMed]
- Hamilos, D.; Leung, D.; Wood, R.; Cunningham, L.; Bean, D.; Yasruel, Z.; Schotman, E.; Hamid, Q. Evidence for distinct cytokine expression in allergic versus nonallergic chronic sinusitis. J. Allergy Clin. Immunol. 1995, 96, 537–544. [Google Scholar] [CrossRef]
- Miljkovic, D.; Psaltis, A.; Wormald, P.; Vreugde, S. T regulatory and Th17 cells in chronic rhinosinusitis with polyps. Int. Forum Allergy Rhinol. 2016, 6, 826–834. [Google Scholar] [CrossRef]
- Sharma, S.; Watanabe, S.; Sivam, A.; Wang, J.; Neuwirth, S.J.; Perez, R.I.; De Tineo, M.; Baroody, F.M.; Naclerio, R.M.; Pinto, J.M. Peripheral Blood and Tissue T Regulatory Cells in Chronic Rhinosinusitis. Am. J. Rhinol. Allergy 2012, 26, 371–379. [Google Scholar] [CrossRef]
- Shen, S.; Lou, H.; Wang, C.; Zhang, L. Macrolide antibiotics in the treatment of chronic rhinosinusitis: Evidence from a meta-analysis. J. Thorac. Dis. 2018, 10, 5913–5923. [Google Scholar] [CrossRef]
- Chen, J.B.; James, L.K.; Davies, A.M.; Wu, Y.B.; Rimmer, J.; Lund, V.J.; Chen, J.H.; McDonnell, J.M.; Chan, Y.C.; Hutchins, G.H.; et al. Antibodies and superantibodies in patients with chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 2017, 139, 1195–1204.e11. [Google Scholar] [CrossRef]
- Antunes, M.B.; Gudis, D.A.; Cohen, N.A. Epithelium, Cilia, and Mucus: Their Importance in Chronic Rhinosinusitis. Immunol. Allergy Clin. N. Am. 2009, 29, 631–643. [Google Scholar] [CrossRef]
- Lund, V.J.; Mackay, I.S. Staging in rhinosinusitis. Rhinology 1993, 31, 183. [Google Scholar] [PubMed]
- Psaltis, A.J.; Li, G.; Vaezeafshar, R.; Cho, K.; Hwang, P.H. Modification of the lund-kennedy endoscopic scoring system improves its reliability and correlation with patient-reported outcome measures. Laryngoscope 2014, 124, 2216–2223. [Google Scholar] [CrossRef] [PubMed]
- Lund, V.J.; Kennedy, D.W. Staging for rhinosinusitis. Otolaryngol.–Head Neck Surg. 1997, 117 (Suppl. S3), S35–S40. [Google Scholar] [CrossRef]
- Hopkins, C.; Gillett, S.; Slack, R.; Lund, V.; Browne, J. Psychometric validity of the 22-item Sinonasal Outcome Test. Clin. Otolaryngol. 2009, 34, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Naidoo, Y.; Tan, N.; Singhal, D.; Wormald, P.J. Chronic rhinosinusitis assessment using the Adelaide Disease Severity Score. J. Laryngol. Otol. 2013, 127, S24–S28. [Google Scholar] [CrossRef]
- Houtak, G.; Nepal, R.; Bouras, G.; Shaghayegh, G.; Bennett, C.; Finnie, J.; Fenix, K.; Psaltis, A.J.; Wormald, P.J.; Vreugde, S. Staphylococcus aureus biofilm secreted factors cause mucosal damage, mast cell infiltration and goblet cell hyperplasia in a rat rhinosinusitis model. bioRxiv 2023. [Google Scholar] [CrossRef]
- Wick, R.; Volkening, J.; Loman, N. Porechop; v0.2.4; Github: San Francisco, CA, USA, 2017; Available online: https://github.com/rrwick/porechop (accessed on 2 May 2023).
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Seemann, T. mlst; v2.19.0; Github: San Francisco, CA, USA; Available online: https://github.com/tseemann/mlst (accessed on 12 July 2020).




{kind=link}
{kind=link}
{kind=link}
| CI-182 | CI-908 | CI-913 |
|---|---|---|
| Male | Female | Female |
| 62 yo | 73 yo | 53 yo |
| GORD and asthma | Asthma | Asthma and aspirin sensitivity |
| CRSwNP | CRSwNP | CRSwNP |
| SNOT 22–34 | SNOT 22–71 | SNOT 22–69 |
| LM: 20 | LM: 20 | LM: 24 |
| LK: 18 | LK: 18 | LK: 20 |
| Histology | Grade |
|---|---|
 | Grade 0: Unaffected area of the pseudostratified ciliated columnar epithelium with a normal number of goblet cells and minimal inflammatory infiltration. The lining epithelium was intact. |
 | Grade 1: Mild lymphoplasmacytic infiltration in the lamina propria. |
 | Grade 2: Moderate lymphoplasmacytic infiltration in the lamina propria with the lymphocytic infiltration of the epithelium lining, resulting in some disorganization and disruption of the epithelium and loss of cilia. |
 | Grade 3: Severe lymphoplasmacytic infiltration of the edematous lamina propria. The respiratory epithelium is disrupted and sometimes ulcerated (top right of image), with marked epithelial lymphocytic infiltration and goblet cell hyperplasia. |
| Magnification | Saline | Exoprotein | Staph CI908 | Staph CI913 |
|---|---|---|---|---|
| 10×#1 |  | |||
| 40×#1 |  | |||
| 10×#2 |  | |||
| 40×#2 |  | |||
| 10×#3 |  | |||
| 40×#3 |  | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murphy, W.; Liu, S.; Hon, K.; Finnie, J.; Bouras, G.S.; Feizi, S.; Houtak, G.; Shaghayegh, G.; Vyskocil, E.; Wormald, P.-J.; et al. A Novel Model of Staphylococcus aureus-Induced Lymphoplasmacytic Rhinosinusitis in Rats. Int. J. Mol. Sci. 2024, 25, 3336. https://doi.org/10.3390/ijms25063336
Murphy W, Liu S, Hon K, Finnie J, Bouras GS, Feizi S, Houtak G, Shaghayegh G, Vyskocil E, Wormald P-J, et al. A Novel Model of Staphylococcus aureus-Induced Lymphoplasmacytic Rhinosinusitis in Rats. International Journal of Molecular Sciences. 2024; 25(6):3336. https://doi.org/10.3390/ijms25063336
Chicago/Turabian StyleMurphy, William, Sha Liu, Karen Hon, John Finnie, George Spyro Bouras, Sholeh Feizi, Ghais Houtak, Gohar Shaghayegh, Erich Vyskocil, Peter-John Wormald, and et al. 2024. "A Novel Model of Staphylococcus aureus-Induced Lymphoplasmacytic Rhinosinusitis in Rats" International Journal of Molecular Sciences 25, no. 6: 3336. https://doi.org/10.3390/ijms25063336