
Molecular Targets of the 5-Amido-Carboxamide Bumped Kinase Inhibitor BKI-1748 in Cryptosporidium parvum and HCT-8 Host Cells

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
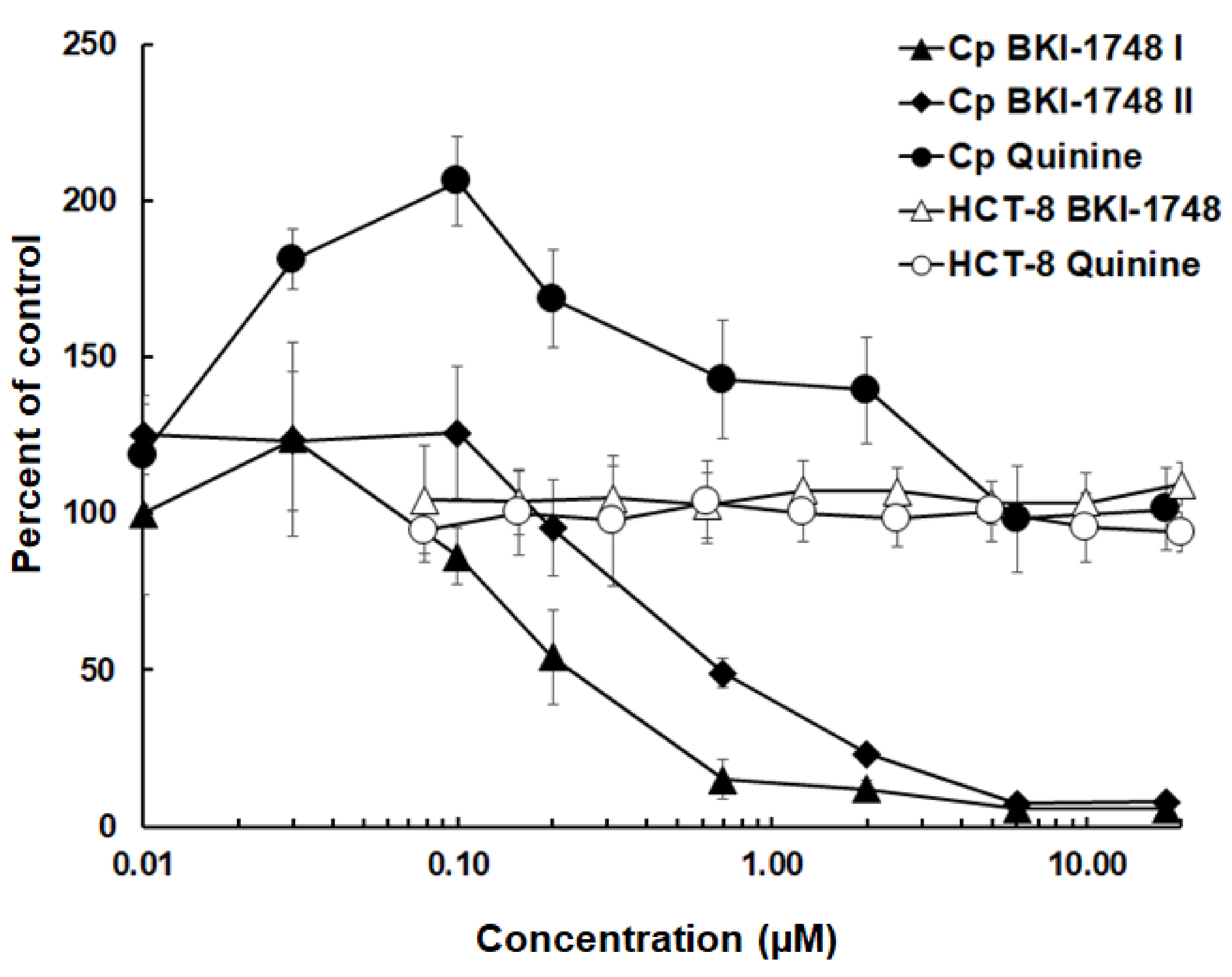
2.1. Inhibition of C. parvum Proliferation In Vitro by BKI-1748
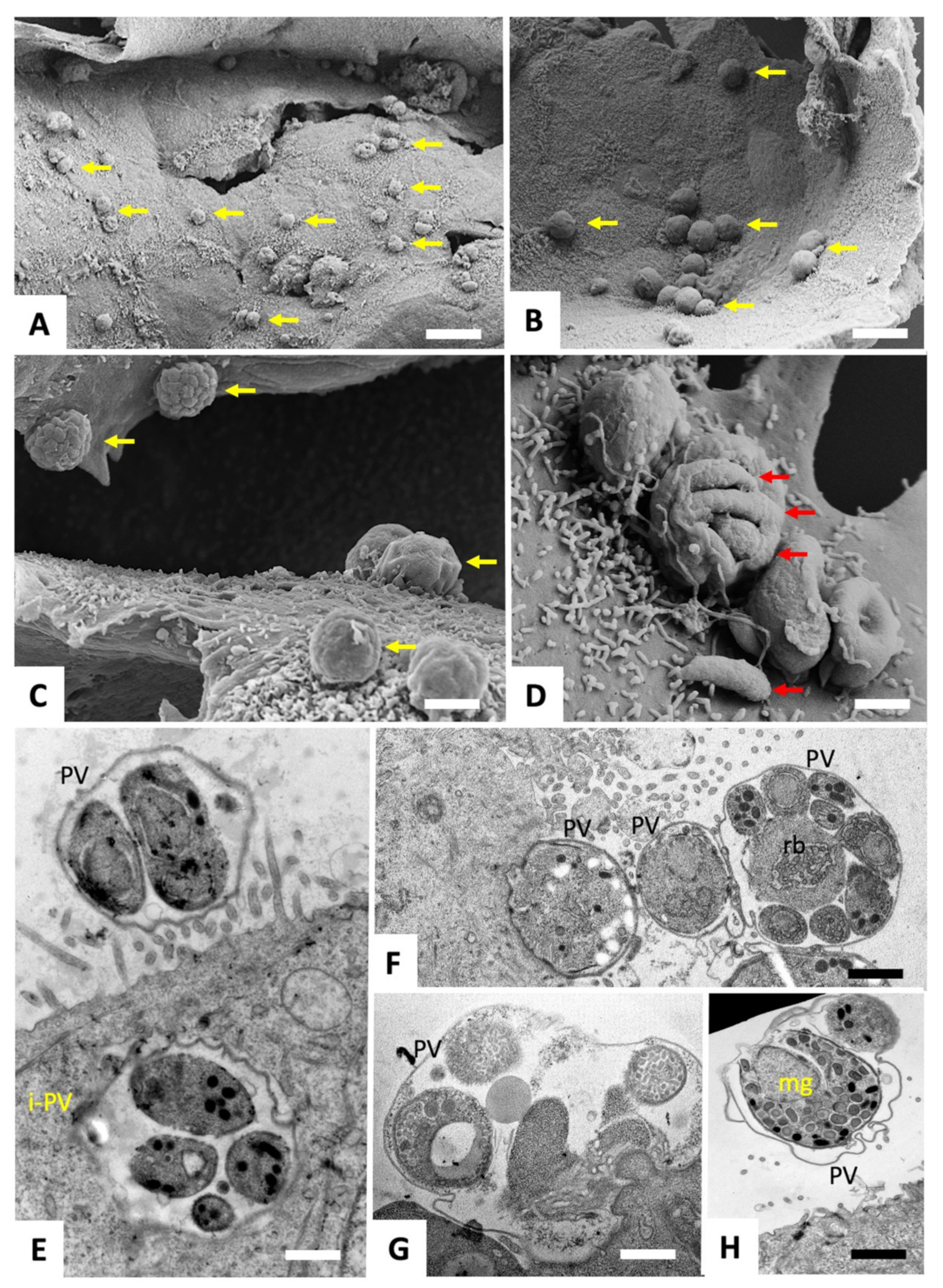
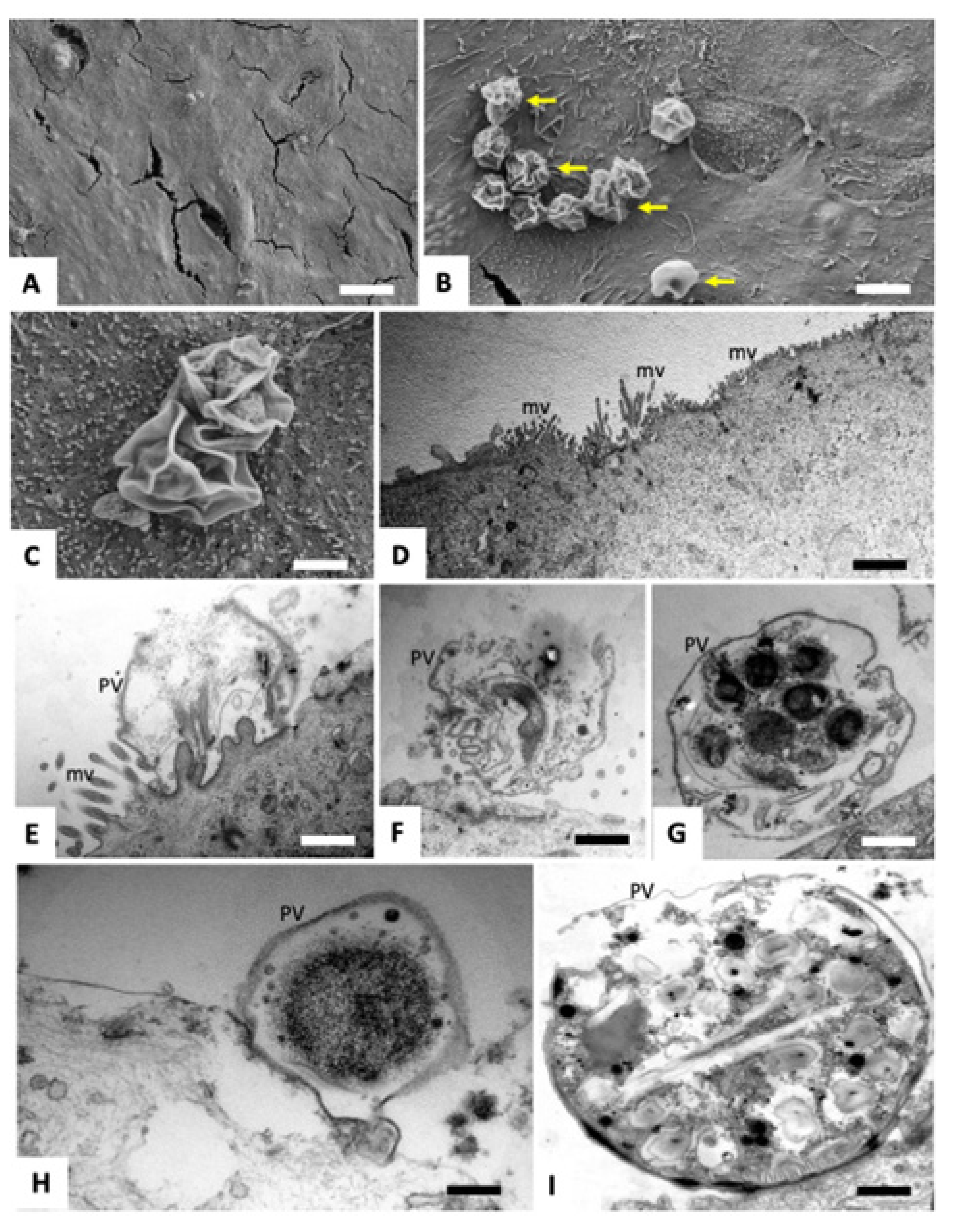
2.2. Scanning and Transmission Electron Microscopy of C. parvum Infected HCT-8 Host Cells Treated or Not with BKI-1748
2.3. DAC Proteomes of Uninfected and C. parvum Infected HCT-8 Host Cells
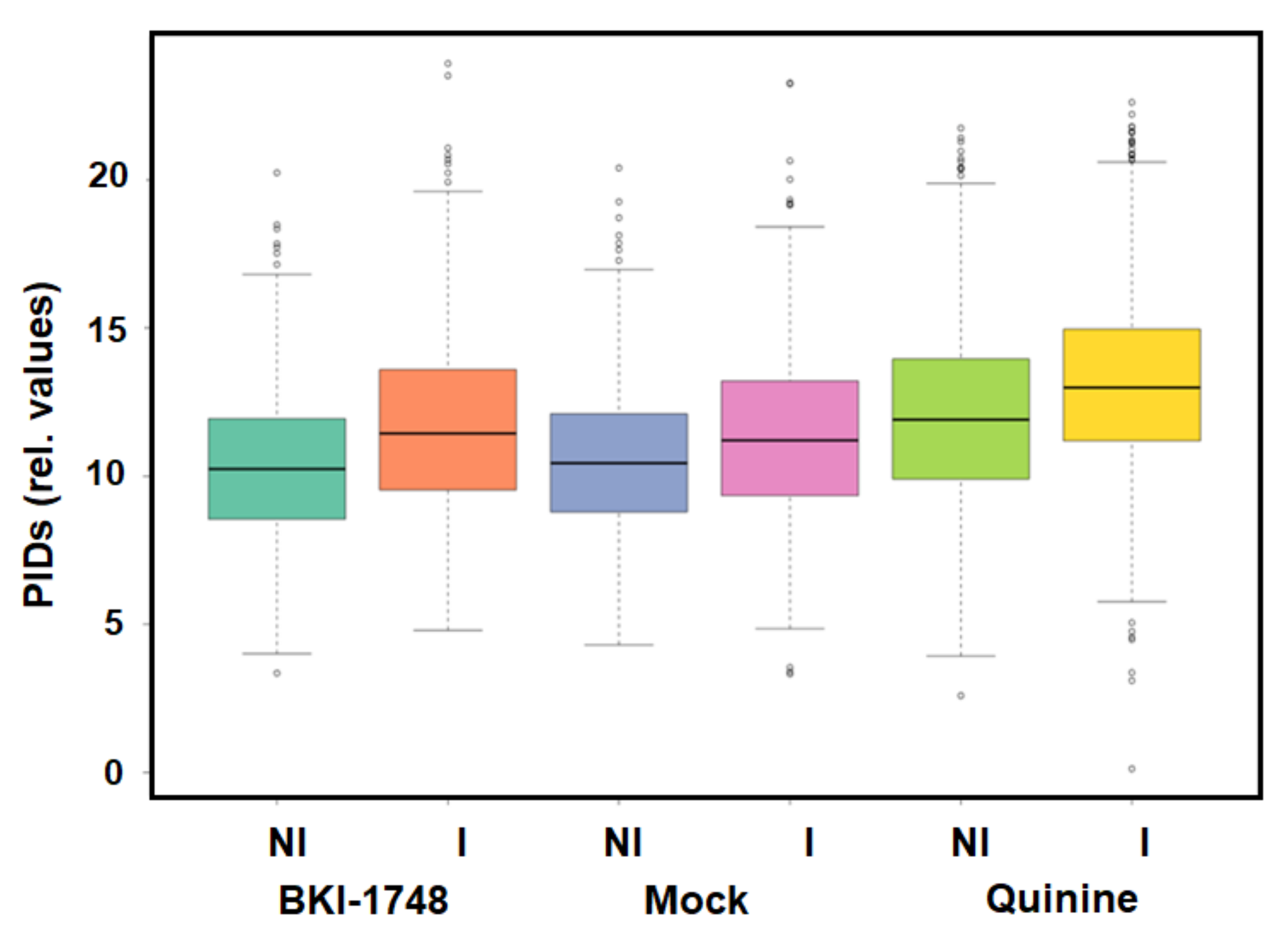
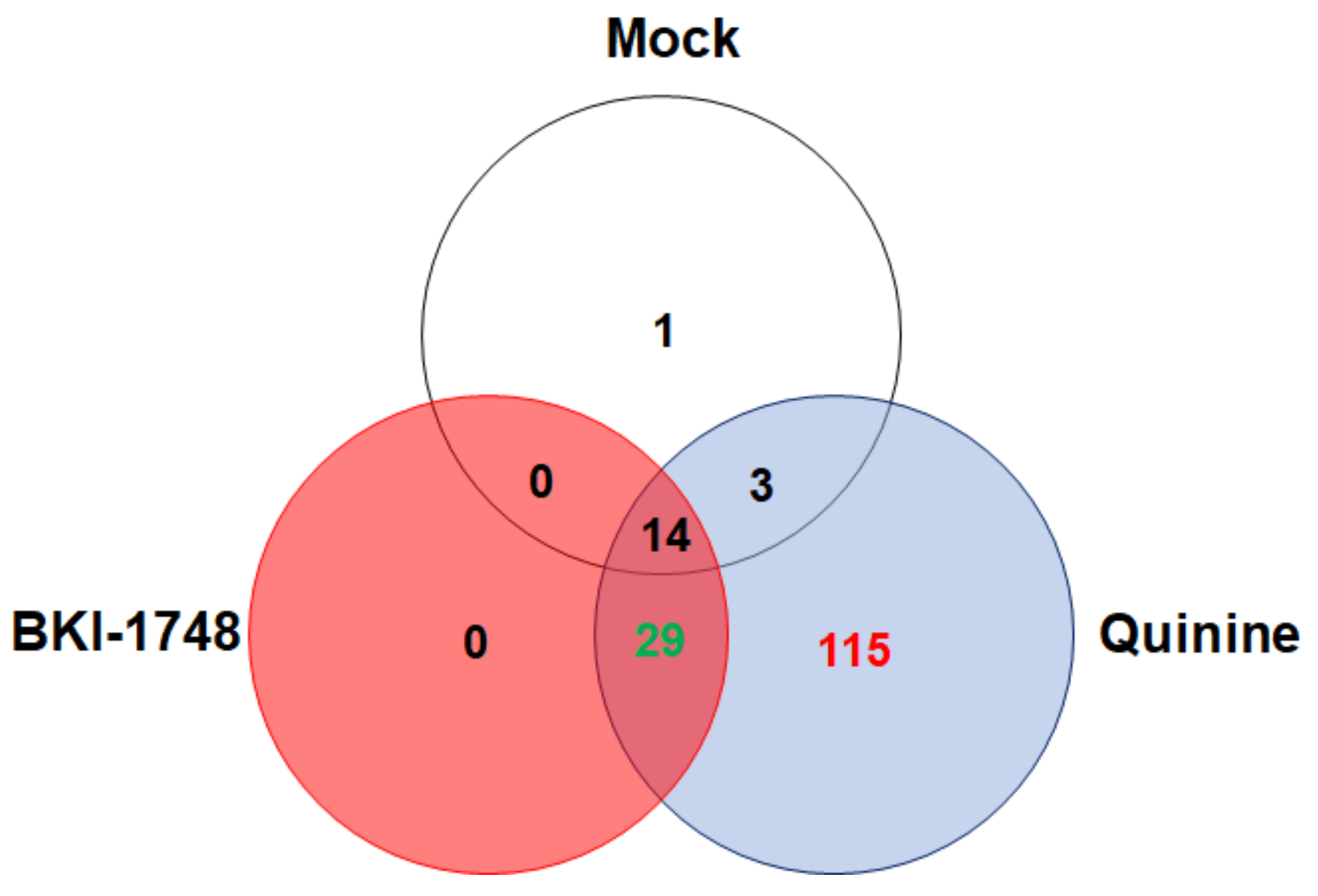
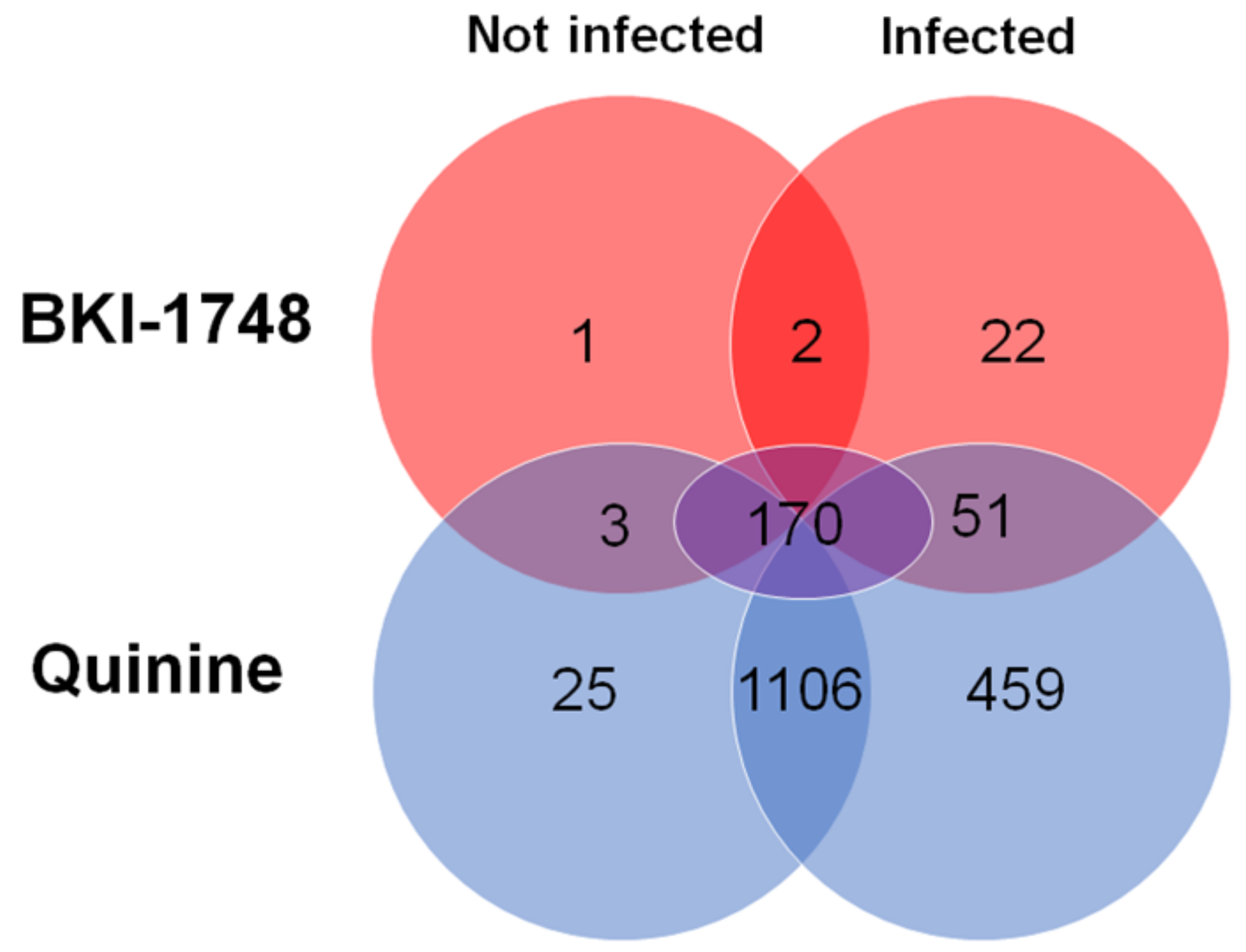
2.3.1. Overview
2.3.2. C. parvum Proteins Identified by DAC
2.3.3. Host Cell Proteins Identified by DAC
3. Discussion
4. Materials and Methods
4.1. Tissue Culture Medium, Biochemicals and Compounds
4.2. In Vitro Culture and Processing of Parasites
4.3. Drug Efficacy Tests
4.4. Scanning and Transmission Electron Microscopy (EM)
4.5. Protein Extraction and Differential Affinity Chromatography
4.6. Proteomic Analysis of the Eluted Proteins by Mass Spectrometry
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pinto, D.J.; Vinayak, S. Cryptosporidium: Host-Parasite Interactions and Pathogenesis. Curr. Clin. Microbiol. Rep. 2021, 8, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Khalil, I.A.; Troeger, C.; Rao, P.C.; Blacker, B.F.; Brown, A.; Brewer, T.G.; Colombara, D.V.; De Hostos, E.L.; Engmann, C.; Guerrant, R.L.; et al. Morbidity, mortality, and long-term consequences associated with diarrhoea from Cryptosporidium infection in children younger than 5 years: A meta-analyses study. Lancet Glob. Health 2018, 6, e758–e768. [Google Scholar] [CrossRef] [PubMed]
- Certad, G. Is Cryptosporidium a hijacker able to drive cancer cell proliferation? Food Waterborne Parasitol. 2022, 27, e00153. [Google Scholar] [CrossRef] [PubMed]
- Guerin, A.; Striepen, B. The biology of the intestinal intracellular parasite Cryptosporidium. Cell Host Microbe 2020, 28, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Zahedi, A.; Paparini, A. Cryptosporidium in humans and animals—A one health approach to prophylaxis. Parasite Immunol. 2016, 38, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Chique, C.; Hynds, P.D.; Andrade, L.; Burke, L.; Morris, D.; Ryan, M.P.; O’Dwyer, J. Cryptosporidium spp. in groundwater supplies intended for human consumption—A descriptive review of global prevalence, risk factors and knowledge gaps. Water Res. 2020, 176, 115726. [Google Scholar] [CrossRef] [PubMed]
- Adeyemo, F.E.; Singh, G.; Reddy, P.; Bux, F.; Stenstrom, T.A. Efficiency of chlorine and UV in the inactivation of Cryptosporidium and Giardia in wastewater. PLoS ONE 2019, 14, e0216040. [Google Scholar] [CrossRef]
- Dhal, A.K.; Panda, C.; Yun, S.I.; Mahapatra, R.K. An update on Cryptosporidium biology and therapeutic avenues. J. Parasite Dis. 2022, 46, 923–939. [Google Scholar] [CrossRef]
- Hemphill, A.; Müller, N.; Müller, J. Thiazolides, a novel class of anti-infective drugs, effective against viruses, bacteria, intracellular and extracellular protozoan parasites and proliferating mammalian cells. Anti-Infect. Agents 2013, 11, 23–31. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, G. High-throughput screening of drugs against the growth of Cryptosporidium parvum in vitro by qRT-PCR. Methods Mol. Biol. 2020, 2052, 319–334. [Google Scholar] [CrossRef]
- Love, M.S.; McNamara, C.W. Phenotypic screening techniques for Cryptosporidium drug discovery. Expert Opin. Drug Discov. 2021, 16, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Bone Relat, R.M.; Winder, P.L.; Bowden, G.D.; Guzman, E.A.; Peterson, T.A.; Pomponi, S.A.; Roberts, J.C.; Wright, A.E.; O’Connor, R.M. High-throughput screening of a marine compound library Identifies anti-Cryptosporidium activity of Leiodolide A. Mar. Drugs 2022, 20, 240. [Google Scholar] [CrossRef] [PubMed]
- Kabir, M.H.B.; Recuenco, F.C.; Mohd Zin, N.K.; Watanabe, N.; Fukuda, Y.; Bando, H.; Watanabe, K.; Bochimoto, H.; Xuan, X.; Kato, K. Identification of potent anti-Cryptosporidium new drug leads by screening traditional Chinese medicines. PLoS Neglected Trop. Dis. 2022, 16, e0010947. [Google Scholar] [CrossRef] [PubMed]
- Ojo, K.K.; Larson, E.T.; Keyloun, K.R.; Castaneda, L.J.; Derocher, A.E.; Inampudi, K.K.; Kim, J.E.; Arakaki, T.L.; Murphy, R.C.; Zhang, L.; et al. Toxoplasma gondii calcium-dependent protein kinase 1 is a target for selective kinase inhibitors. Nat. Struct. Mol. Biol. 2010, 17, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Keyloun, K.R.; Reid, M.C.; Choi, R.; Song, Y.; Fox, A.M.W.; Hillesland, H.K.; Zhang, Z.; Vidadala, R.; Merritt, E.A.; Lau, A.O.T.; et al. The gatekeeper residue and beyond: Homologous calcium-dependent protein kinases as drug development targets for veterinarian Apicomplexa parasites. Parasitology 2014, 141, 1499–1509. [Google Scholar] [CrossRef] [PubMed]
- Van Voorhis, W.C.; Doggett, J.S.; Parsons, M.; Hulverson, M.A.; Choi, R.; Arnold, S.L.M.; Riggs, M.W.; Hemphill, A.; Howe, D.K.; Mealey, R.H.; et al. Extended-spectrum antiprotozoal bumped kinase inhibitors: A review. Exp. Parasitol. 2017, 180, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Choi, R.; Hulverson, M.A.; Huang, W.; Vidadala, R.S.R.; Whitman, G.R.; Barrett, L.K.; Schaefer, D.A.; Betzer, D.P.; Riggs, M.W.; Doggett, J.S.; et al. Bumped Kinase Inhibitors as therapy for apicomplexan parasitic diseases: Lessons learned. Int. J. Parasitol. 2020, 50, 413–422. [Google Scholar] [CrossRef]
- Murphy, R.C.; Ojo, K.K.; Larson, E.T.; Castellanos-Gonzalez, A.; Perera, B.G.; Keyloun, K.R.; Kim, J.E.; Bhandari, J.G.; Muller, N.R.; Verlinde, C.L.; et al. Discovery of potent and selective inhibitors of calcium-dependent protein kinase 1 (CDPK1) from C. parvum and T. gondii. ACS Med. Chem. Lett. 2010, 1, 331–335. [Google Scholar] [CrossRef]
- Castellanos-Gonzalez, A.; Sparks, H.; Nava, S.; Huang, W.; Zhang, Z.; Rivas, K.; Hulverson, M.A.; Barrett, L.K.; Ojo, K.K.; Fan, E.; et al. A novel calcium-dependent kinase inhibitor, bumped kinase inhibitor 1517, cures cryptosporidiosis in immunosuppressed mice. J. Infect. Dis. 2016, 214, 1850–1855. [Google Scholar] [CrossRef]
- Hulverson, M.A.; Vinayak, S.; Choi, R.; Schaefer, D.A.; Castellanos-Gonzalez, A.; Vidadala, R.S.R.; Brooks, C.F.; Herbert, G.T.; Betzer, D.P.; Whitman, G.R.; et al. Bumped-kinase inhibitors for cryptosporidiosis therapy. J. Infect. Dis. 2017, 215, 1275–1284. [Google Scholar] [CrossRef]
- Kuhlenschmidt, T.B.; Rutaganira, F.U.; Long, S.; Tang, K.; Shokat, K.M.; Kuhlenschmidt, M.S.; Sibley, L.D. Inhibition of calcium-dependent protein kinase 1 (CDPK1) in vitro by pyrazolopyrimidine derivatives does not correlate with sensitivity of Cryptosporidium parvum growth in cell culture. Antimicrob. Agents Chemother. 2016, 60, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, H.H.; Nava, M.G.; Gartlan, B.E.; Rose, S.; Vinayak, S. A conditional protein degradation system to study essential gene function in Cryptosporidium parvum. mBio 2020, 11, 10-1128. [Google Scholar] [CrossRef]
- Zhang, Q.; Shao, Q.; Guo, Y.; Li, N.; Li, Y.; Su, J.; Xu, R.; Zhang, Z.; Xiao, L.; Feng, Y. Characterization of three calcium-dependent protein kinases of Cryptosporidium parvum. Front. Microbiol. 2020, 11, 622203. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Shen, Y.; Li, N.; Li, Y.; Zhang, Z.; Xiao, L.; Guo, Y.; Feng, Y. Comparative characterization of CpCDPK1 and CpCDPK9, two potential drug targets against cryptosporidiosis. Microorganisms 2022, 10, 333. [Google Scholar] [CrossRef] [PubMed]
- Ojo, K.K.; Reid, M.C.; Kallur Siddaramaiah, L.; Müller, J.; Winzer, P.; Zhang, Z.; Keyloun, K.R.; Vidadala, R.S.; Merritt, E.A.; Hol, W.G.; et al. Neospora caninum calcium-dependent protein kinase 1 is an effective drug target for neosporosis therapy. PLoS ONE 2014, 9, e92929. [Google Scholar] [CrossRef] [PubMed]
- Winzer, P.; Müller, J.; Aguado-Martinez, A.; Rahman, M.; Balmer, V.; Manser, V.; Ortega-Mora, L.M.; Ojo, K.K.; Fan, E.; Maly, D.J.; et al. In vitro and in vivo effects of the bumped kinase inhibitor 1294 in the related cyst-forming apicomplexans Toxoplasma gondii and Neospora caninum. Antimicrob. Agents Chemother. 2015, 59, 6361–6374. [Google Scholar] [CrossRef] [PubMed]
- Imhof, D.; Anghel, N.; Winzer, P.; Balmer, V.; Ramseier, J.; Hanggeli, K.; Choi, R.; Hulverson, M.A.; Whitman, G.R.; Arnold, S.L.M.; et al. In vitro activity, safety and in vivo efficacy of the novel bumped kinase inhibitor BKI-1748 in non-pregnant and pregnant mice experimentally infected with Neospora caninum tachyzoites and Toxoplasma gondii oocysts. Int. J. Parasitol. Drugs Drug Resist. 2021, 16, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Aguado-Martinez, A.; Balmer, V.; Maly, D.J.; Fan, E.; Ortega-Mora, L.M.; Ojo, K.K.; Van Voorhis, W.C.; Hemphill, A. Two novel calcium-dependent protein kinase 1 inhibitors interfere with vertical transmission in mice infected with Neospora caninum tachyzoites. Antimicrob. Agents Chemother. 2017, 61, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Anghel, N.; Winzer, P.A.; Imhof, D.; Müller, J.; Langa, X.; Rieder, J.; Barrett, L.K.; Vidadala, R.S.R.; Huang, W.; Choi, R.; et al. Comparative assessment of the effects of bumped kinase inhibitors on early zebrafish embryo development and pregnancy in mice. Int. J. Antimicrob. Agents 2020, 56, 106099. [Google Scholar] [CrossRef]
- Müller, J.; Anghel, N.; Imhof, D.; Hanggeli, K.; Uldry, A.C.; Braga-Lagache, S.; Heller, M.; Ojo, K.K.; Ortega-Mora, L.M.; Van Voorhis, W.C.; et al. Common molecular targets of a quinolone based bumped kinase Inhibitor in Neospora caninum and Danio rerio. Int. J. Mol. Sci. 2022, 23, 2381. [Google Scholar] [CrossRef]
- Liles, N.W.; Page, E.E.; Liles, A.L.; Vesely, S.K.; Raskob, G.E.; George, J.N. Diversity and severity of adverse reactions to quinine: A systematic review. Am. J. Hematol. 2016, 91, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Jumani, R.S.; Hasan, M.M.; Stebbins, E.E.; Donnelly, L.; Miller, P.; Klopfer, C.; Bessoff, K.; Teixeira, J.E.; Love, M.S.; McNamara, C.W.; et al. A suite of phenotypic assays to ensure pipeline diversity when prioritizing drug-like Cryptosporidium growth inhibitors. Nat. Commun. 2019, 10, 1862. [Google Scholar] [CrossRef] [PubMed]
- Mauzy, M.J.; Enomoto, S.; Lancto, C.A.; Abrahamsen, M.S.; Rutherford, M.S. The Cryptosporidium parvum transcriptome during in vitro development. PLoS ONE 2012, 7, e31715. [Google Scholar] [CrossRef] [PubMed]
- Neuwald, A.F.; Aravind, L.; Spouge, J.L.; Koonin, E.V. AAA+: A class of chaperone-like ATPases associated with the assembly, operation, and disassembly of protein complexes. Genome Res. 1999, 9, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, Y.; Masutani, C.; Hanaoka, F.; Kimura, H.; Tsurimoto, T. A second proliferating cell nuclear antigen loader complex, Ctf18-replication factor C, stimulates DNA polymerase eta activity. J. Biol. Chem. 2007, 282, 20906–20914. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, B.W.; Heizmann, C.W. The S100 family of EF-hand calcium-binding proteins: Functions and pathology. Trends Biochem. Sci. 1996, 21, 134–140. [Google Scholar] [CrossRef]
- Hu, L.; Liu, Y.; Fu, C.; Zhao, J.; Cui, Q.; Sun, Q.; Wang, H.; Lu, L.; Dai, H.; Xu, X.; et al. The tumorigenic effect of the high expression of ladinin-1 in lung adenocarcinoma and Its potential as a therapeutic target. Molecules 2023, 28, 1103. [Google Scholar] [CrossRef]
- Xu, S.J.; Jin, B.; Zhao, W.J.; Chen, X.X.; Tong, Y.Y.; Ding, X.F.; Chen, Y.Y.; Wang, D.H.; Wang, Z.M.; Dai, B.Q.; et al. The specifically androgen-regulated Gene (SARG) promotes capillary thyroid carcinoma (PTC) lymphatic metastasis through vascular endothelial growth factor C (VEGF-C) and VEGF receptor 3 (VEGFR-3) axis. Front. Oncol. 2022, 12, 817660. [Google Scholar] [CrossRef]
- Theodos, C.M.; Griffiths, J.K.; D’Onfro, J.; Fairfield, A.; Tzipori, S. Efficacy of nitazoxanide against Cryptosporidium parvum in cell culture and in animal models. Antimicrob. Agents Chemother. 1998, 42, 1959–1965. [Google Scholar] [CrossRef]
- Gargala, G.; Delaunay, A.; Li, X.; Brasseur, P.; Favennec, L.; Ballet, J.J. Efficacy of nitazoxanide, tizoxanide and tizoxanide glucuronide against Cryptosporidium parvum development in sporozoite-infected HCT-8 enterocytic cells. J. Antimicrob. Chemother. 2000, 46, 57–60. [Google Scholar] [CrossRef]
- You, X.; Schinazi, R.F.; Arrowood, M.J.; Lejkowski, M.; Juodawlkis, A.S.; Mead, J.R. In-vitro activities of paromomycin and lasalocid evaluated in combination against Cryptosporidium parvum. J. Antimicrob. Chemother. 1998, 41, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Sonzogni-Desautels, K.; Renteria, A.E.; Camargo, F.V.; Di Lenardo, T.Z.; Mikhail, A.; Arrowood, M.J.; Fortin, A.; Ndao, M. Oleylphosphocholine (OlPC) arrests Cryptosporidium parvum growth in vitro and prevents lethal infection in interferon gamma receptor knock-out mice. Front. Microbiol. 2015, 6, 973. [Google Scholar] [CrossRef] [PubMed]
- Stoll, S.W.; Zhao, X.; Elder, J.T. EGF stimulates transcription of CaN19 (S100A2) in HaCaT keratinocytes. J. Investig. Dermatol. 1998, 111, 1092–1097. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Haase-Kohn, C.; Pietzsch, J. S100A2 in cancerogenesis: A friend or a foe? Amino Acids 2011, 41, 849–861. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.L.; Fan, X.C.; Li, Y.H.; Yuan, Y.J.; Yin, Y.L.; Wang, X.T.; Zhang, L.X.; Zhao, G.H. Expression profiles of mRNA and lncRNA in HCT-8 cells infected with Cryptosporidium parvum IId subtype. Front. Microbiol. 2018, 9, 1409. [Google Scholar] [CrossRef] [PubMed]
- Green, R.; Noller, H.F. Ribosomes and translation. Annu. Rev. Biochem. 1997, 66, 679–716. [Google Scholar] [CrossRef] [PubMed]
- Klinge, S.; Voigts-Hoffmann, F.; Leibundgut, M.; Arpagaus, S.; Ban, N. Crystal structure of the eukaryotic 60S ribosomal subunit in complex with initiation factor 6. Science 2011, 334, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Wahl, M.C.; Will, C.L.; Luhrmann, R. The spliceosome: Design principles of a dynamic RNP machine. Cell 2009, 136, 701–718. [Google Scholar] [CrossRef]
- Gold, D.; Stein, B.; Tzipori, S. The utilization of sodium taurocholate in excystation of Cryptosporidium parvum and infection of tissue culture. J. Parasitol. 2001, 87, 997–1000. [Google Scholar] [CrossRef]
- Bessoff, K.; Sateriale, A.; Lee, K.K.; Huston, C.D. Drug repurposing screen reveals FDA-approved inhibitors of human HMG-CoA reductase and isoprenoid synthesis that block Cryptosporidium parvum growth. Antimicrob. Agents Chemother. 2013, 57, 1804–1814. [Google Scholar] [CrossRef]
- O’Brien, J.; Wilson, I.; Orton, T.; Pognan, F. Investigation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur. J. Biochem. 2000, 267, 5421–5426. [Google Scholar] [CrossRef]
- Müller, J.; Aguado-Martinez, A.; Manser, V.; Balmer, V.; Winzer, P.; Ritler, D.; Hostettler, I.; Arranz-Solis, D.; Ortega-Mora, L.; Hemphill, A. Buparvaquone is active against Neospora caninum in vitro and in experimentally infected mice. Int. J. Parasitol. Drugs Drug Resist. 2015, 5, 16–25. [Google Scholar] [CrossRef]
- Schlange, C.; Müller, J.; Imhof, D.; Hänggeli, K.P.A.; Boubaker, G.; Ortega-Mora, L.M.; Wong, H.N.; Haynes, R.K.; Van Voorhis, W.C.; Hemphill, A. Single and combination treatment of Toxoplasma gondii infections with a bumped kinase inhibitor and artemisone in vitro and with artemiside in experimentally infected mice. Exp. Parasitol. 2023, 255, 108655. [Google Scholar] [CrossRef]
- Yu, F.; Haynes, S.E.; Teo, G.C.; Avtonomov, D.M.; Polasky, D.A.; Nesvizhskii, A.I. Fast quantitative analysis of timsTOF PASEF data with MSFragger and IonQuant. Mol. Cell. Proteom. 2020, 19, 1575–1585. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein ID | Annotation | rAbu |
|---|---|---|
| QOY39990.1 | Cold-shock DNA-binding domain-containing protein | 2631 |
| QOY40836.1 | Thioredoxin | 1687 |
| QOY40770.1 | 60S ribosomal protein L6 | 1173 |
| QOY42506.1 | Profilin | 983 |
| QOY42741.1 | 60S ribosomal protein L30 | 565 |
| QOY41364.1 | Tubulin beta chain | 562 |
| QOY42473.1 | Ribosomal protein L14 | 527 |
| QOY40573.1 | 60S ribosomal protein L35A | 463 |
| QOY41580.1 | Nucleoside diphosphate kinase | 438 |
| EAZ51383.1 | Guanine nucleotide-binding protein, putative | 404 |
| QOY43535.1 | Actin | 395 |
| QOY40892.1 | Disulfide-isomerase, signal peptide plus ER retention motif | 377 |
| QOY43201.1 | 60S acidic ribosomal protein LP2 | 354 |
| QOY41122.1 | Divalent ion tolerance protein, CutA/nitrogen regulatory protein PII/ATP phosphoribosyltransferase | 337 |
| QOY41334.1 | Uncharacterized protein with Armadillo-like helical | 296 |
| EAZ51402.1 | 60S ribosomal protein L13, putative | 288 |
| QOY40344.1 | 50S-L18Ae/60S-L20/60S-L18A ribosomal protein | 286 |
| QOY40195.1 | Uncharacterized protein CPATCC_0004510 | 236 |
| EAZ51528.1 | Poly(a)-binding protein fabm, putative | 216 |
| QOY42201.1 | 60S ribosomal protein L31 | 208 |
| Protein ID | Annotation | rAbu BKI-1748 | rAbu Quinine |
|---|---|---|---|
| QOY43223.1 | 60S ribosomal protein L39 | 1386 | 462 |
| EAZ51504.1 | Ribosomal protein S23, partial | 51 | 43 |
| QOY40793.1 | Uncharacterized protein CPATCC_0010570 (RF-C Ctf18p AAA + ATPase) | 48 | 3 |
| QOY40752.1 | Uncharacterized protein CPATCC_0010160 | 28 | 10 |
| QOY41888.1 | Hypothetical protein CPATCC_0017110 | 22 | 16 |
| ABJ09792.1 | Heat shock protein 70, partial | 22 | 20 |
| QOY40881.1 | Uncharacterized protein CPATCC_0011490 | 21 | 13 |
| QOY41279.1 | Uncharacterized protein with tetratricopeptide-like helical (Garp protein) | 9 | 1 |
| Protein ID | Annotation | rAbu |
|---|---|---|
| Not infected cells | ||
| P18887 | DNA repair protein XRCC1 | 100 |
| Q9BW04-2 | Isoform 2 of specifically androgen-regulated gene protein * | 23 |
| O00515 | Ladinin-1 * | 10 |
| Infected cells | ||
| P29034 | Protein S100-A2 | 259 |
| P81605-2 | Isoform 2 of dermcidin | 84 |
| P01036 | Cystatin-S | 83 |
| P10909-2 | Isoform 2 of clusterin | 69 |
| Q9UBH0 | Interleukin-36 receptor antagonist protein | 49 |
| P56134-2 | Isoform 2 of ATP synthase subunit f, mitochondrial | 44 |
| Q9H190-3 | Isoform 3 of syntenin-2 | 40 |
| Q01546 | Keratin, type II cytoskeletal 2 oral | 34 |
| P01037 | Cystatin-SN | 33 |
| P20933 | N(4)-(beta-N-acetylglucosaminyl)-L-asparaginase | 16 |
| Q9UQB8-2 | Isoform 2 of brain-specific angiogenesis inhibitor 1-associated protein 2 | 13 |
| Q3ZCW2 | Galectin-related protein | 13 |
| O14556 | Glyceraldehyde-3-phosphate dehydrogenase, testis specific | 12 |
| P35580-2 | Isoform 2 of myosin-10 | 12 |
| Q96L46 | Calpain small subunit 2 | 11 |
| O95817 | BAG family molecular chaperone regulator 3 | 10 |
| Q9BY77-2 | Isoform 2 of polymerase delta-interacting protein 3 | 8 |
| Q8N6H7-2 | Isoform 2 of ADP-ribosylation factor GTPase-activating protein 2 | 7 |
| Q14157-1 | Isoform 2 of ubiquitin-associated protein 2-like | 6 |
| O00515 | Ladinin-1 * | 4 |
| Q93009-3 | Isoform 3 of ubiquitin carboxyl-terminal hydrolase 7 | 4 |
| P0DOX6 | Immunoglobulin mu heavy chain | 3 |
| Q9BW04-2 | Isoform 2 of specifically androgen-regulated gene protein * | 3 |
| O75592-2 | Isoform 2 of E3 ubiquitin-protein ligase MYCBP2 | 1 |
| Protein ID | Annotation | rAbu NI | BKI-1748 I | rAbu NI | Quinine I |
|---|---|---|---|---|---|
| C9JLW8 | Mapk-regulated corepressor-interacting protein 1 | 221 | 38 | 47 | 26.5 |
| Q15637-2 | Isoform 2 of splicing factor 1 | 86 | 139 | 33 | 17.9 |
| P29144 | Tripeptidyl-peptidase 2 | 83 | 6 | 6 | 5.8 |
| P36957 | Dihydrolipoyllysine-residue succinyltransferase component of 2-oxoglutarate dehydrogenase complex, mitochondrial | 22 | 8 | 5.9 | 6.2 |
| Q13573 | SNW domain-containing protein 1 | 17 | 13 | 17.0 | 7.9 |
| Q9UHB6-4 | Isoform 4 of LIM domain and actin-binding protein 1 | 16 | 11 | 8.5 | 5.6 |
| Protein ID | Annotation | rAbu NI | BKI-1748 I | rAbu NI | Quinine I |
|---|---|---|---|---|---|
| Q15437 | Protein transport protein Sec23B | 94 | 19 | 1234 | 2780 |
| P51991 | Heterogeneous nuclear ribonucleoprotein A3 | 145 | 107 | 1212 | 1938 |
| Q15436 | Protein transport protein Sec23A | 17 | 4 | 719 | 1735 |
| Q13162 | Peroxiredoxin-4 | 22 | 7 | 619 | 1036 |
| Q86X55-1 | Isoform 1 of histone-arginine methyltransferase CARM1 | 26 | 16 | 594 | 1016 |
| Q14103-3 | Isoform 3 of heterogeneous nuclear ribonucleoprotein D0 | 122 | 26 | 512 | 963 |
| Q12906-7 | Isoform 7 of interleukin enhancer-binding factor 3 | 28 | 23 | 731 | 928 |
| O96019 | Actin-like protein 6A | 156 | 4 | 1435 | 883 |
| Q15434 | RNA-binding motif, single-stranded-interacting protein 2 | 38 | 37 | 1091 | 869 |
| Q9Y2R9 | 28S ribosomal protein S7, mitochondrial | 113 | 17 | 1660 | 836 |
| Q9BUJ2-2 | Isoform 2 of heterogeneous nuclear ribonucleoprotein U-like protein 1 | 129 | 217 | 401 | 782 |
| A1KXE4 | Myelin-associated neurite-outgrowth inhibitor | 584 | 306 | 381 | 490 |
| P61247 | 40S ribosomal protein S3a | 390 | 206 | 790 | 384 |
| P09496-2 | Isoform non-brain of clathrin light chain A | 139 | 36 | 630 | 375 |
| P46779 | 60S ribosomal protein L28 | 672 | 264 | 819 | 337 |
| P82930 | 28S ribosomal protein S34, mitochondrial | 26 | 31 | 420 | 314 |
| Q99460 | 26S proteasome non-ATPase regulatory subunit 1 | 16 | 370 | 326 | 165 |
| P15880 | 40S ribosomal protein S2 | 81 | 10 | 291 | 146 |
| Q3MHD2-2 | Isoform 2 of protein LSM12 (NAADP receptor) | 362 | 51 | 225 | 78 |
| Q8WWM7 | Ataxin-2-like protein | 142 | 102 | 228 | 54 |
| Function | HCT-8 Cells | C. parvum |
|---|---|---|
| DNA binding and modification | 18 | 2 |
| RNA binding and modification | 72 | 16 |
| Protein binding and modification | 29 | 1 |
| Cytoskeleton and intracellular transport | 43 | 2 |
| Intracellular signaling | 18 | 0 |
| Energy and intermediary metabolism | 44 | 0 |
| Hypothetical or ambiguous | 0 | 8 |
| Total | 224 | 29 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ajiboye, J.; Uldry, A.-C.; Heller, M.; Naguleswaran, A.; Fan, E.; Van Voorhis, W.C.; Hemphill, A.; Müller, J. Molecular Targets of the 5-Amido-Carboxamide Bumped Kinase Inhibitor BKI-1748 in Cryptosporidium parvum and HCT-8 Host Cells. Int. J. Mol. Sci. 2024, 25, 2707. https://doi.org/10.3390/ijms25052707
Ajiboye J, Uldry A-C, Heller M, Naguleswaran A, Fan E, Van Voorhis WC, Hemphill A, Müller J. Molecular Targets of the 5-Amido-Carboxamide Bumped Kinase Inhibitor BKI-1748 in Cryptosporidium parvum and HCT-8 Host Cells. International Journal of Molecular Sciences. 2024; 25(5):2707. https://doi.org/10.3390/ijms25052707
Chicago/Turabian StyleAjiboye, Jubilee, Anne-Christine Uldry, Manfred Heller, Arunasalam Naguleswaran, Erkang Fan, Wesley C. Van Voorhis, Andrew Hemphill, and Joachim Müller. 2024. "Molecular Targets of the 5-Amido-Carboxamide Bumped Kinase Inhibitor BKI-1748 in Cryptosporidium parvum and HCT-8 Host Cells" International Journal of Molecular Sciences 25, no. 5: 2707. https://doi.org/10.3390/ijms25052707