
Expression Patterns and Molecular Mechanisms Regulating Drought Tolerance of Soybean [Glycine max (L.) Merr.] Conferred by Transcription Factor Gene GmNAC19
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Expression of Soybean Transcription Factor Gene GmNAC19
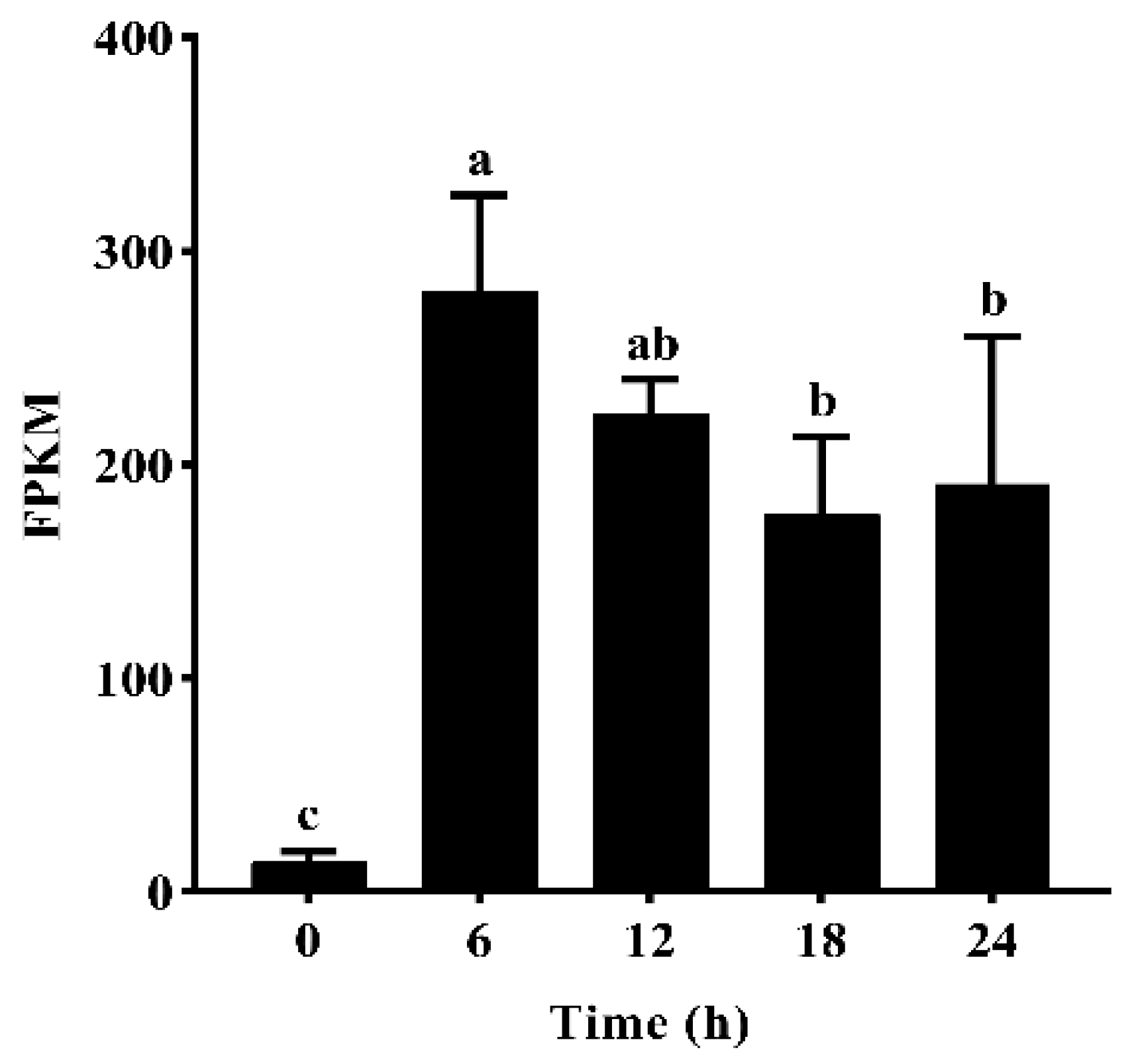
2.1.1. Gene Expression of GmNAC19 Based on Transcriptome Analysis of Drought-Treated Soybean Roots
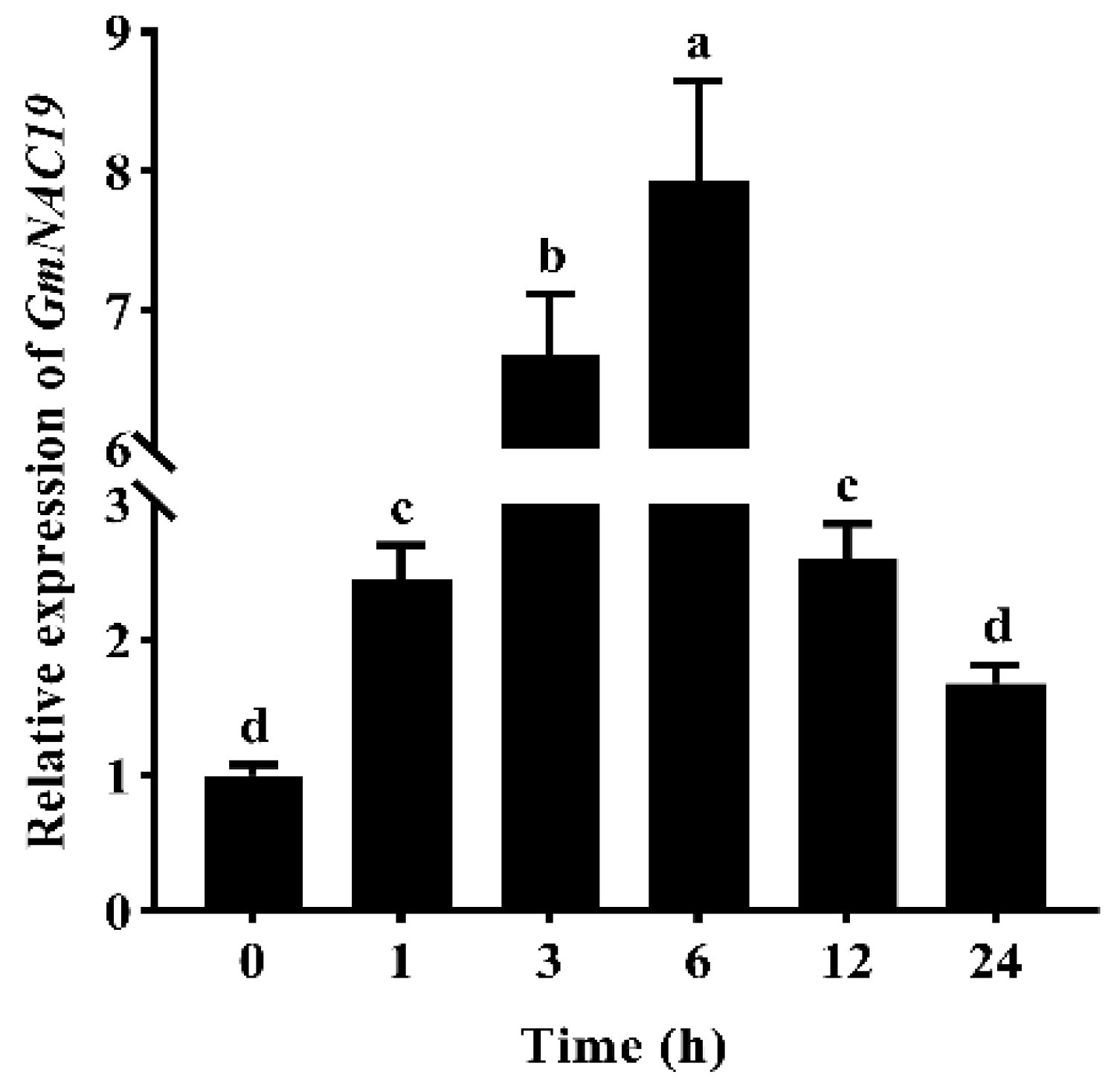
2.1.2. Gene Expression of GmNAC19 in Soybean Treated with PEG6000 Based on Quantitative Real-Time PCR
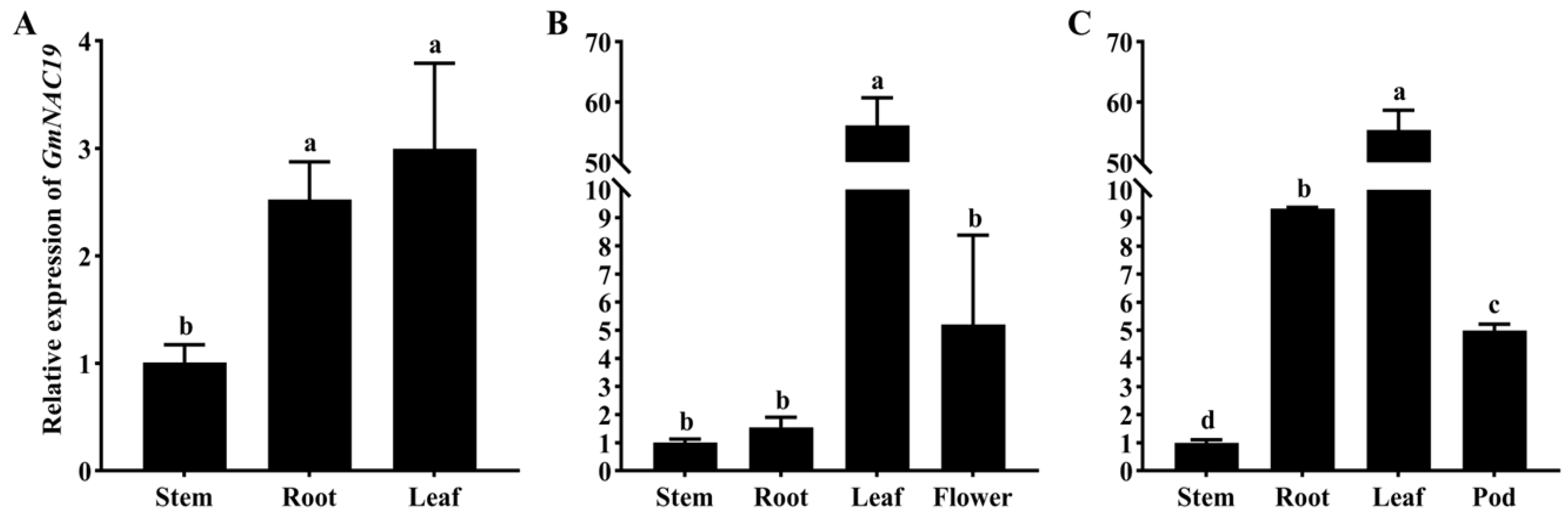
2.1.3. Gene Expression of GmNAC19 in Different Organs of Soybean at Three Different Developmental Stages
2.2. Molecular Mechanism Underlying the Enhancement in Drought Tolerance by Soybean GmNAC19
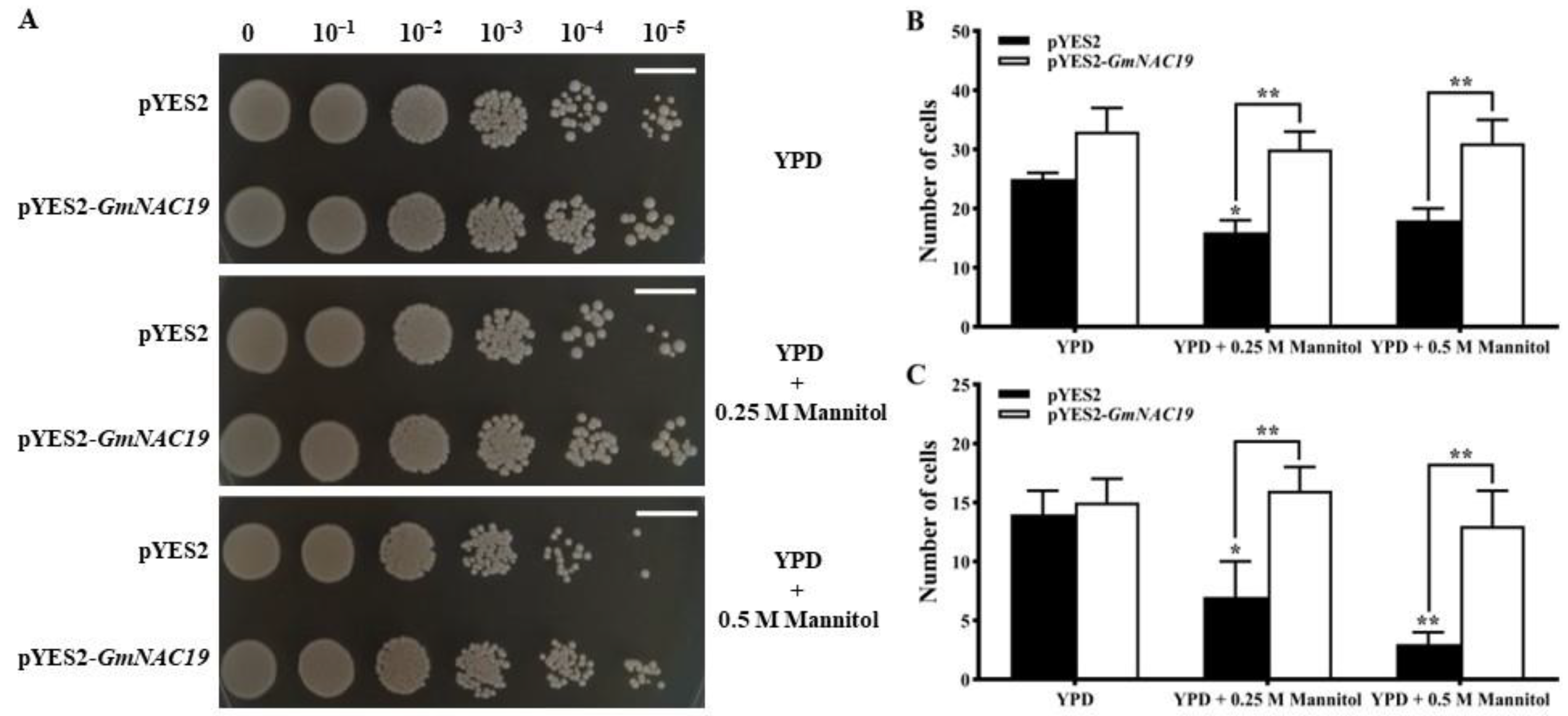
2.2.1. Enhanced Drought Tolerance in Saccharomyces cerevisiae Conferred by Soybean GmNAC19
2.2.2. Enhanced Drought Tolerance in Transgenic Arabidopsis thaliana with GmNAC19
- Drought tolerance of transgenic Arabidopsis thaliana with GmNAC19 under drought stress simulated by PEG6000
- (1)
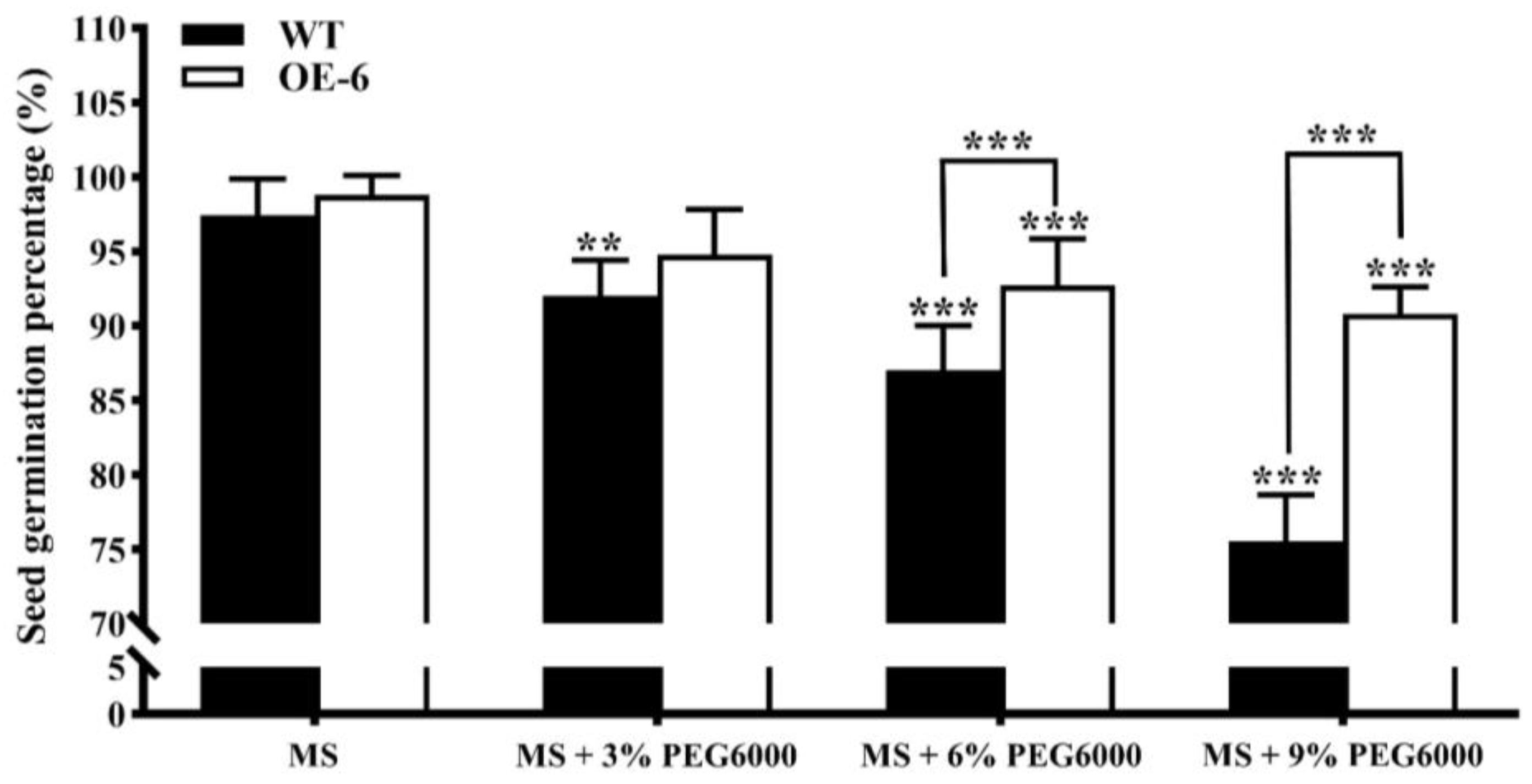
- Seed germination of transgenic Arabidopsis thaliana with GmNAC19
- (2)
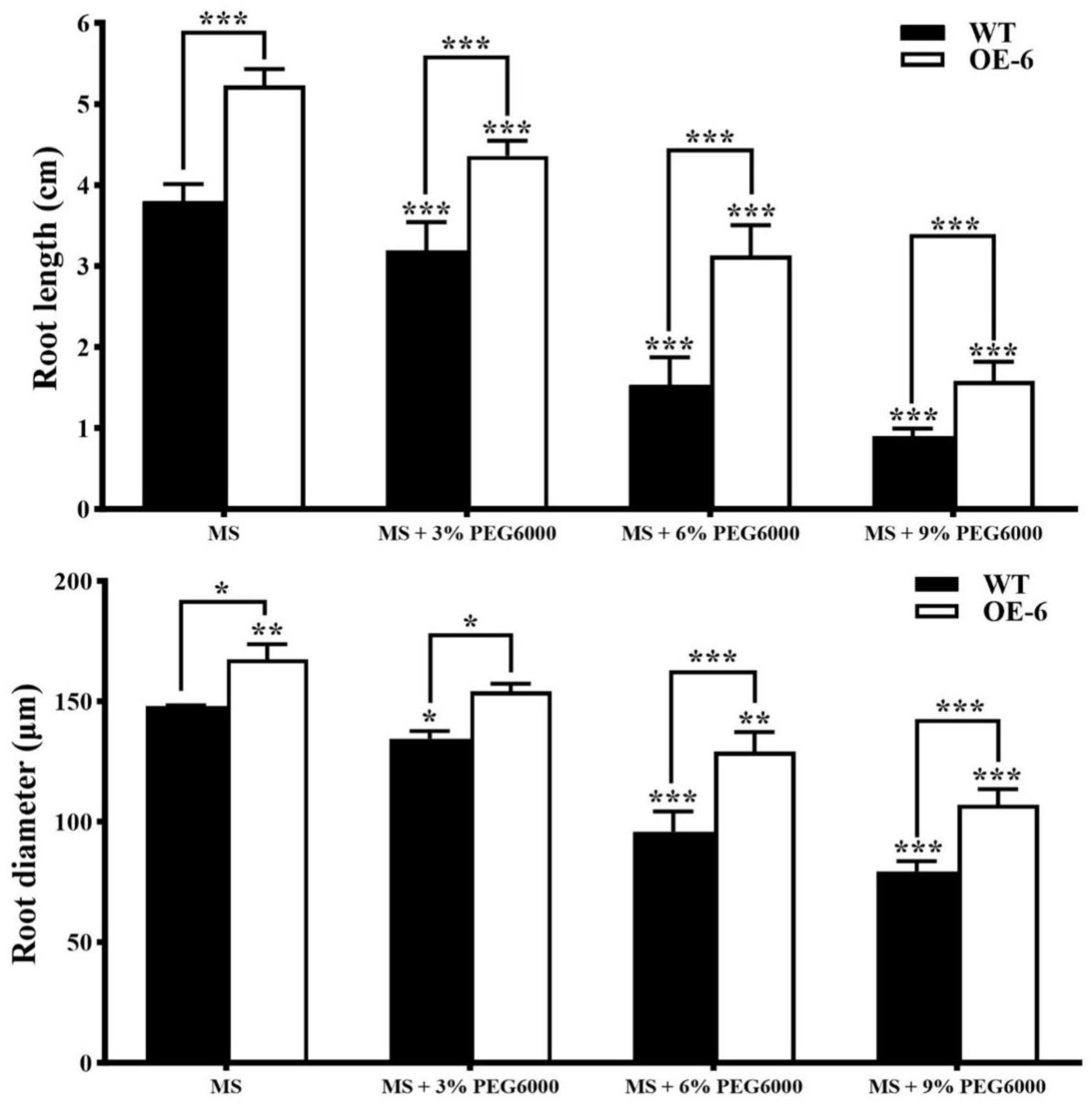
- Growth of roots in transgenic Arabidopsis thaliana with GmNAC19
- (3)
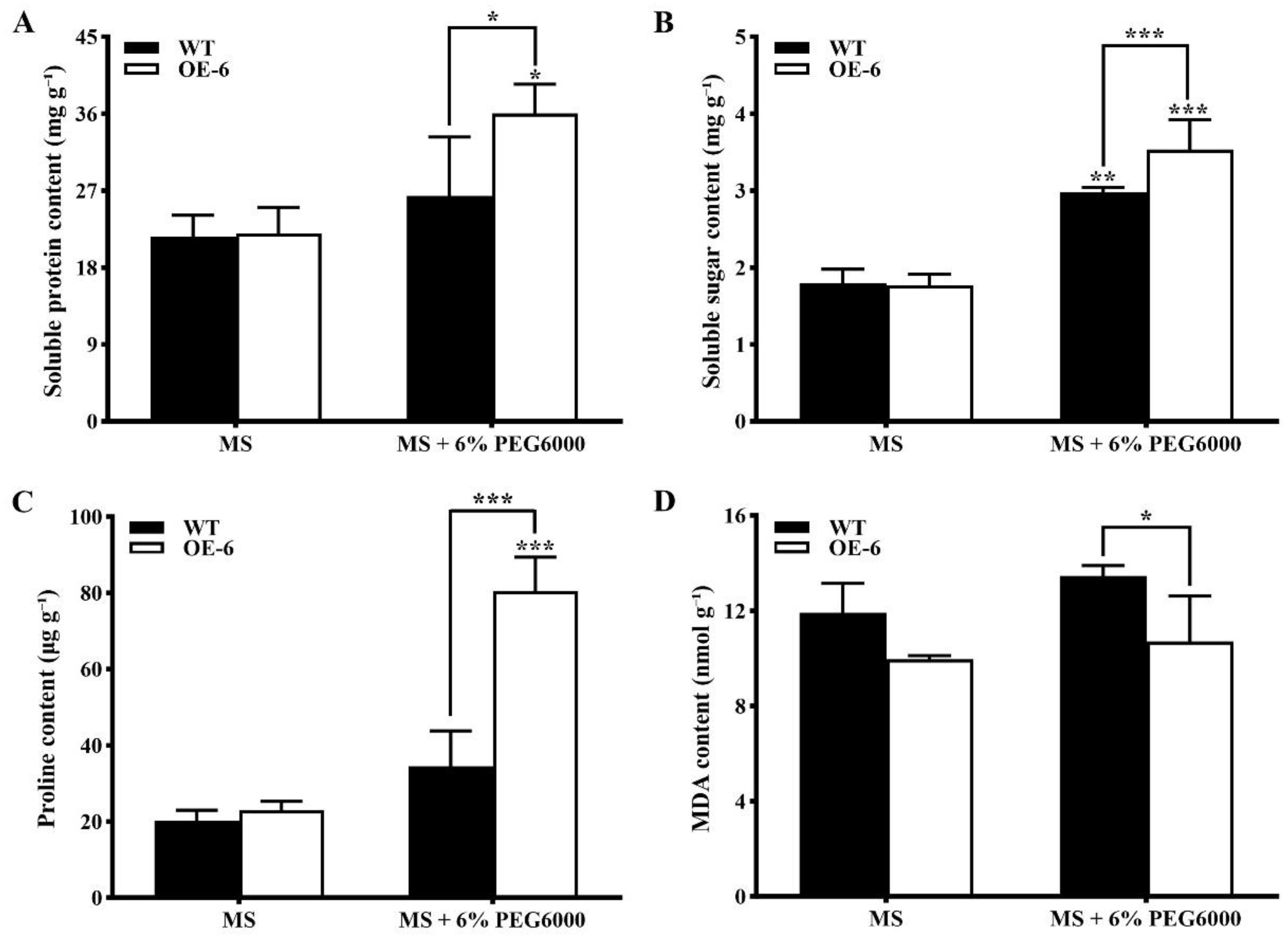
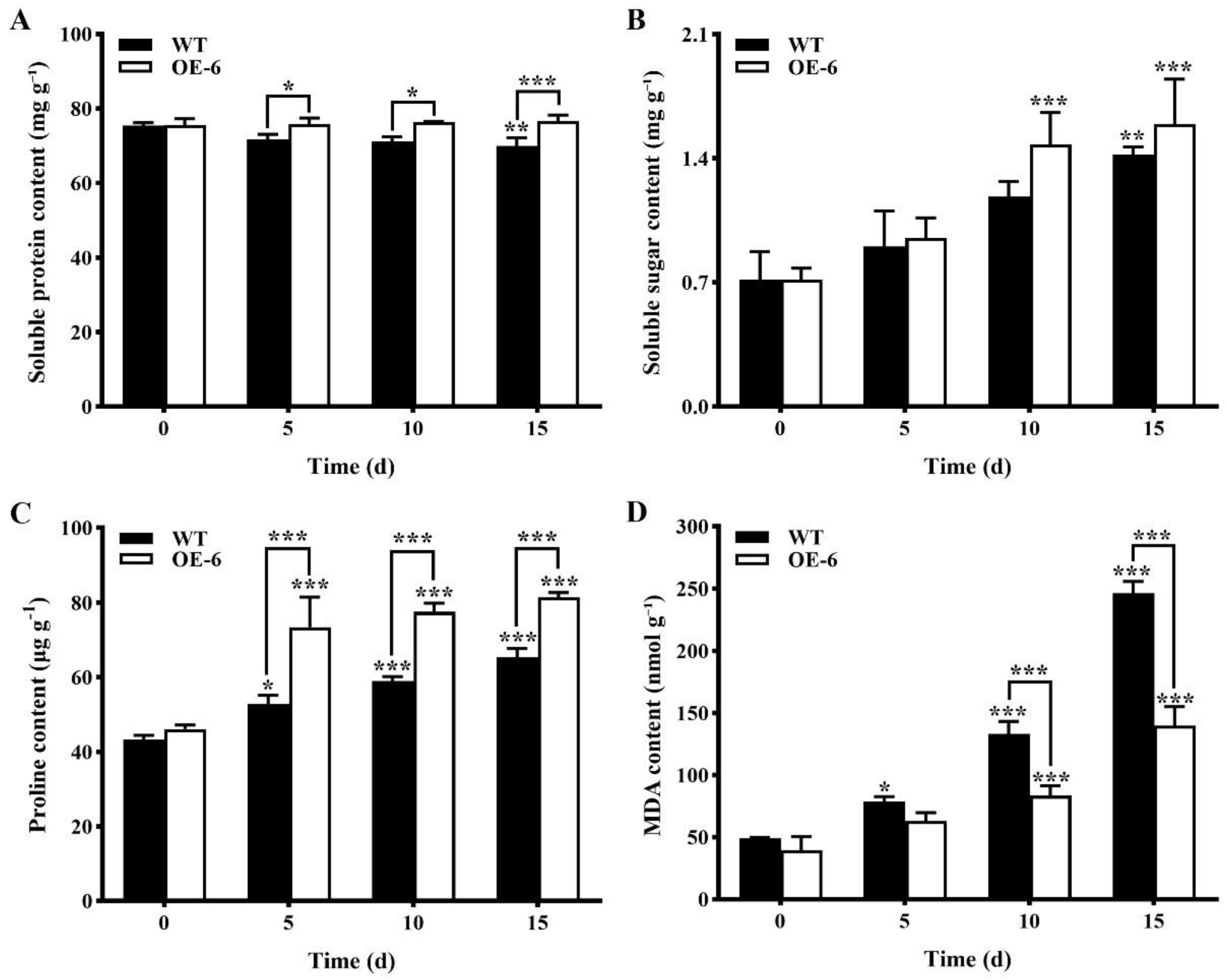
- Contents of physiological and biochemical indices related to drought stress in transgenic Arabidopsis thaliana with GmNAC19 under drought stress simulated by PEG6000
- (4)
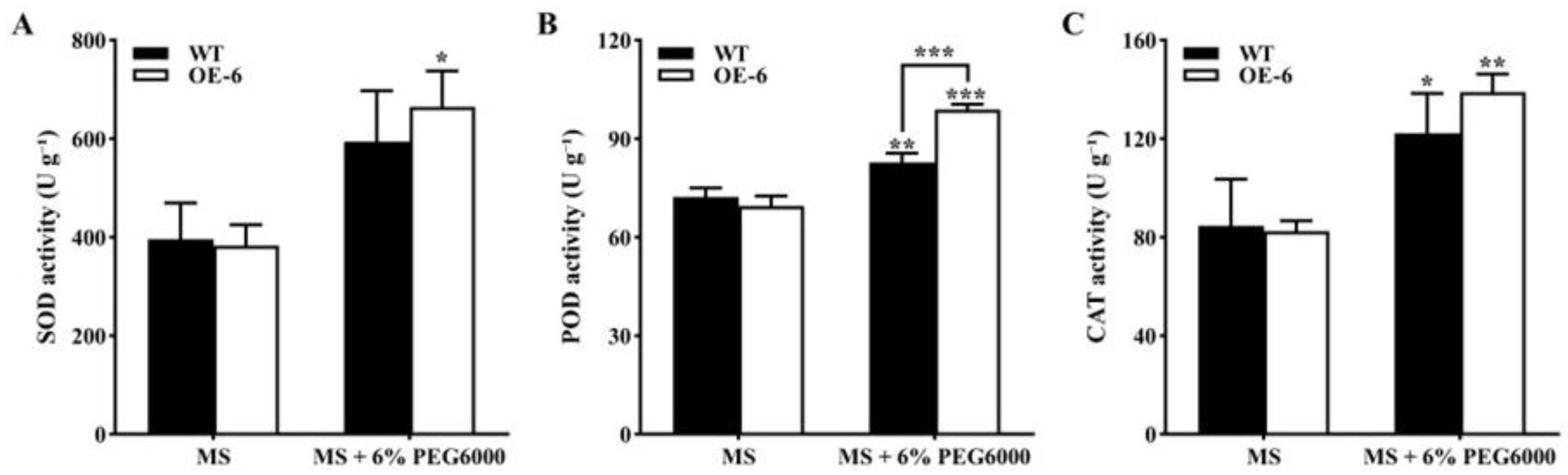
- Activities of antioxidant enzymes in transgenic Arabidopsis thaliana with GmNAC19 under drought stress simulated by PEG6000
- Drought tolerance of transgenic Arabidopsis thaliana with GmNAC19 under drought stress induced by water loss
- (1)
- Contents of physiological and biochemical indices related to drought stress of transgenic Arabidopsis thaliana with GmNAC19 under drought stress induced by water loss
- (2)
- Activities of antioxidant enzymes of transgenic Arabidopsis thaliana with GmNAC19 under drought stress induced by water loss
2.2.3. Enhanced Drought Tolerance in Transgenic Soybean Composite Lines with GmNAC19
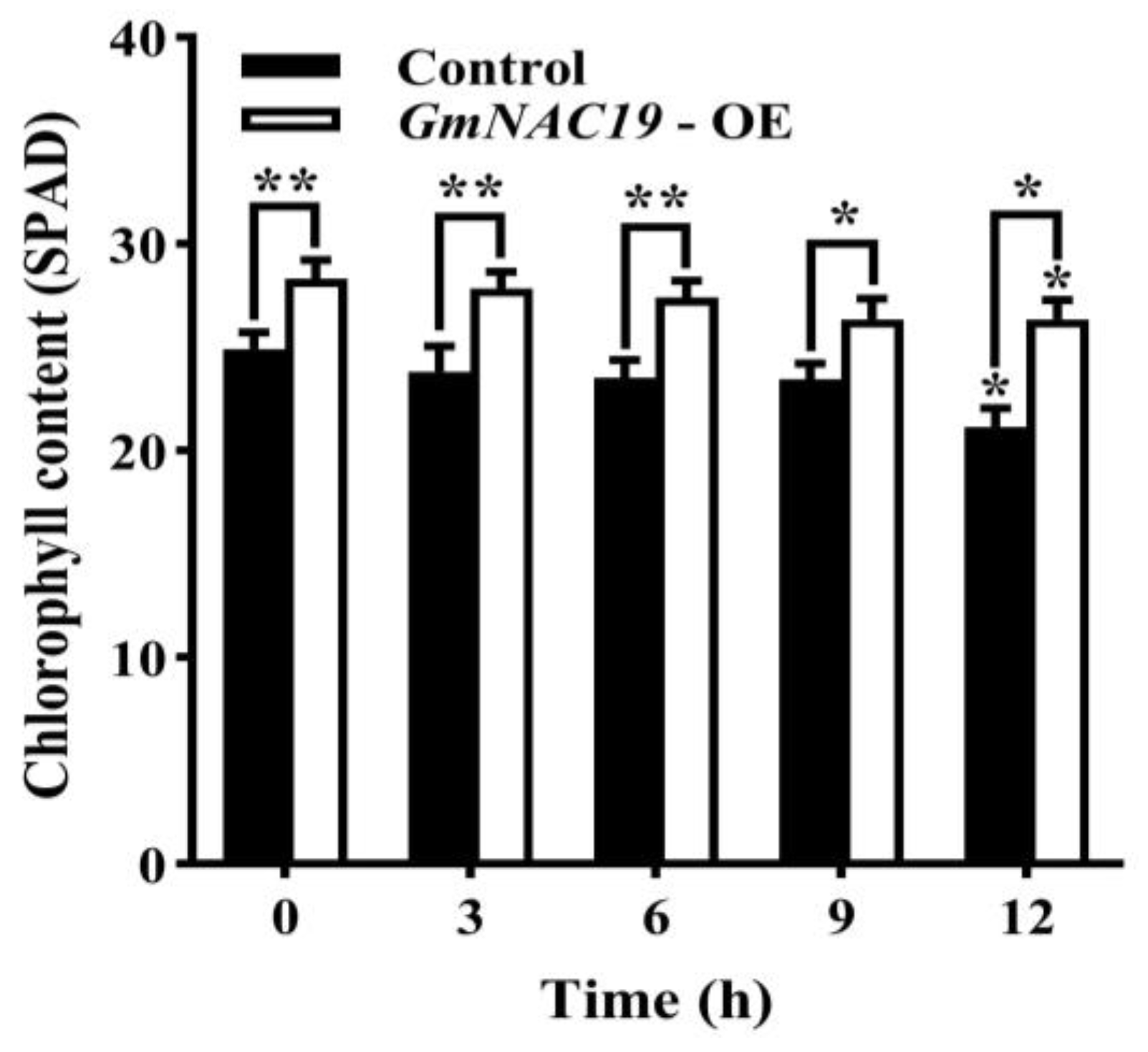
- Content of chlorophyll in transgenic soybean composite lines with GmNAC19 under drought stress simulated by 20% PEG6000
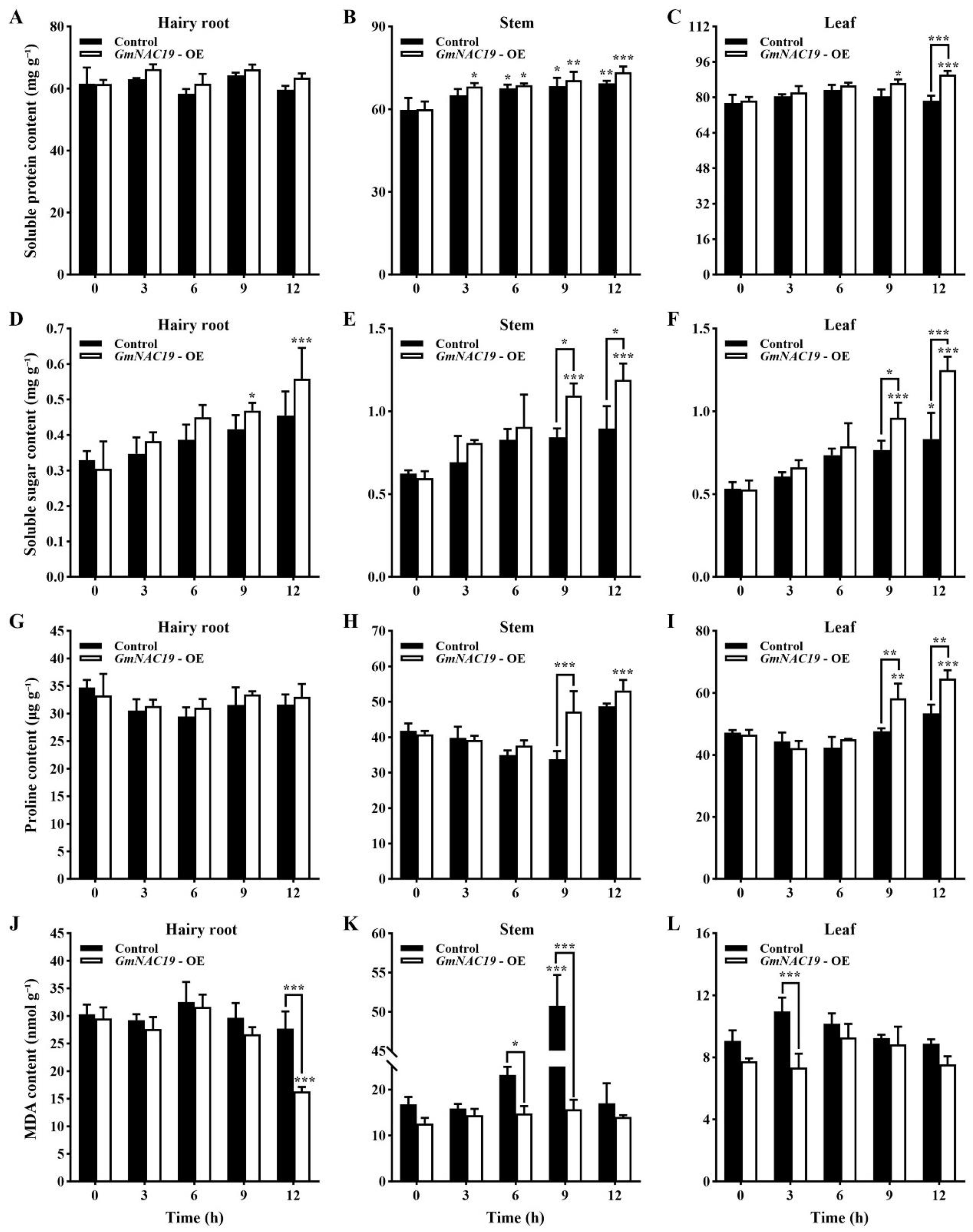
- Contents of physiological and biochemical indicators related to drought stress in transgenic soybean composite lines with GmNAC19 under drought stress simulated by 20% PEG6000
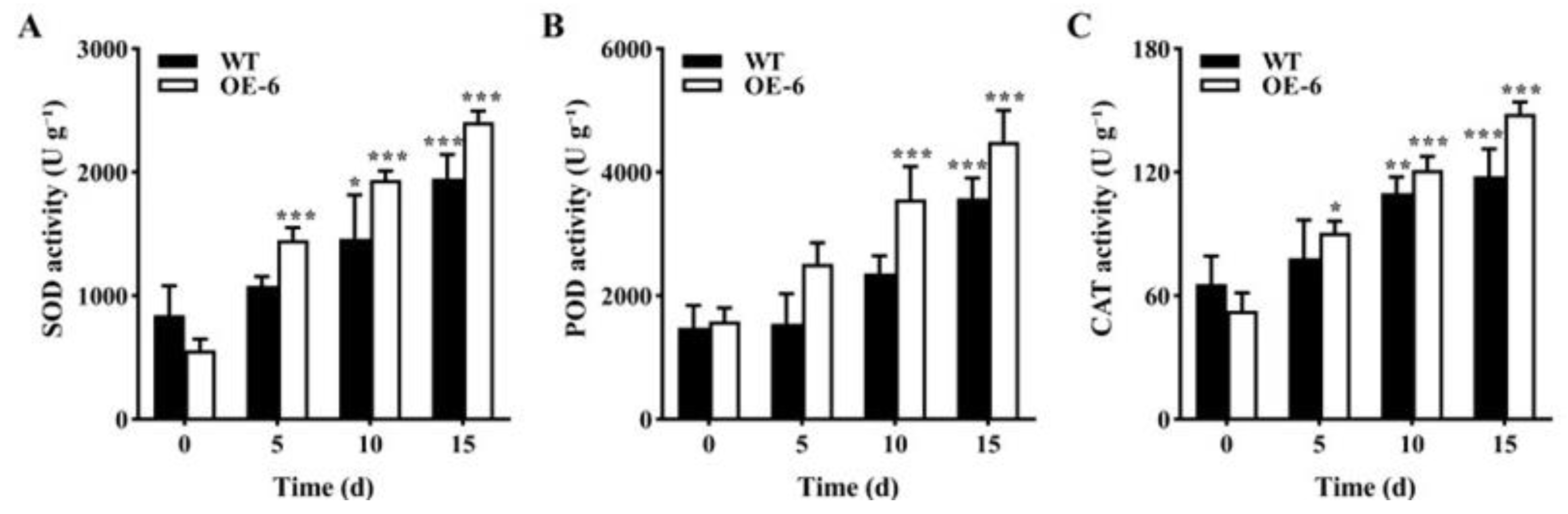
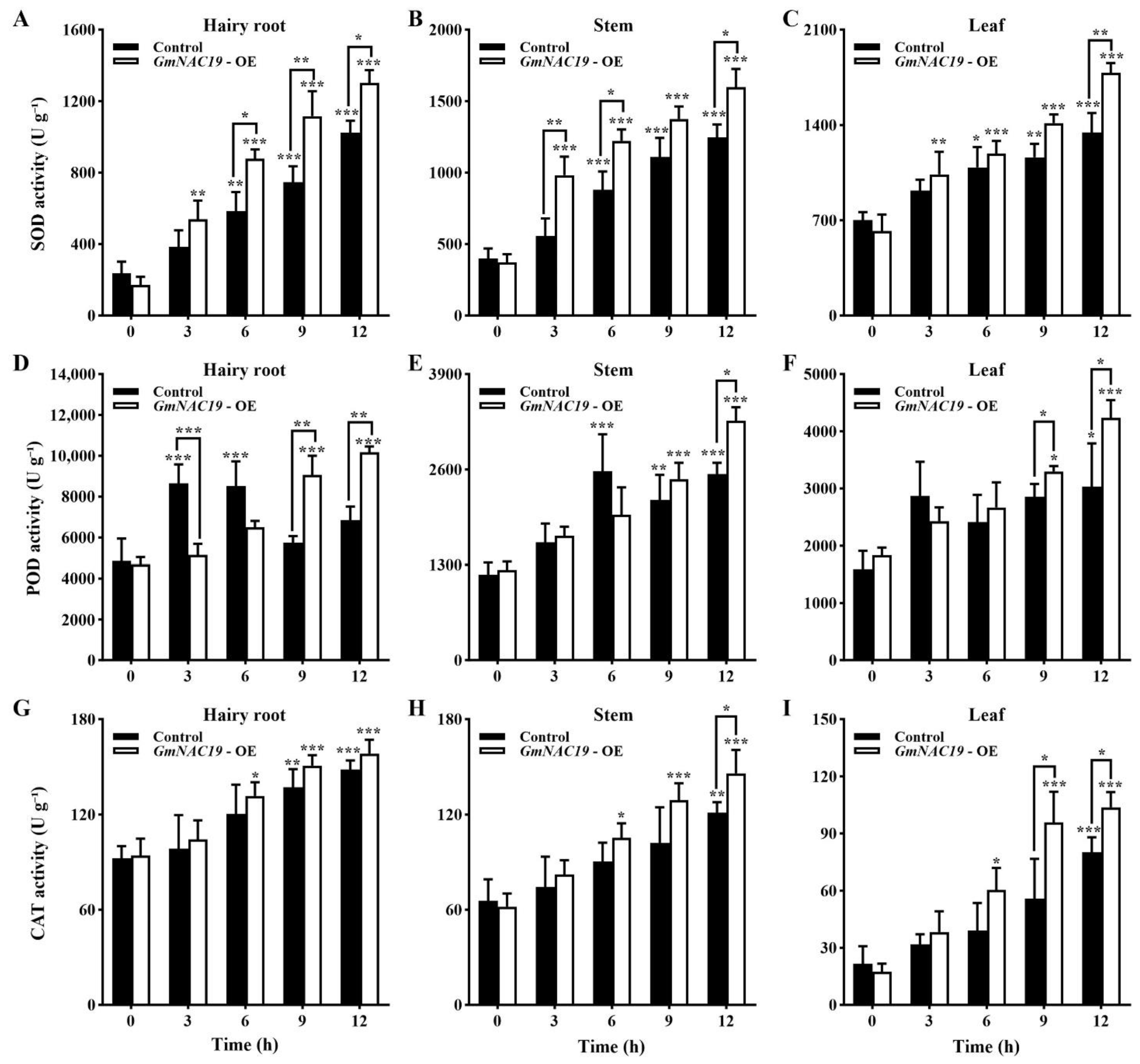
- Antioxidant enzyme activities of transgenic soybean composite lines with GmNAC19 under drought stress simulated by 20% PEG6000
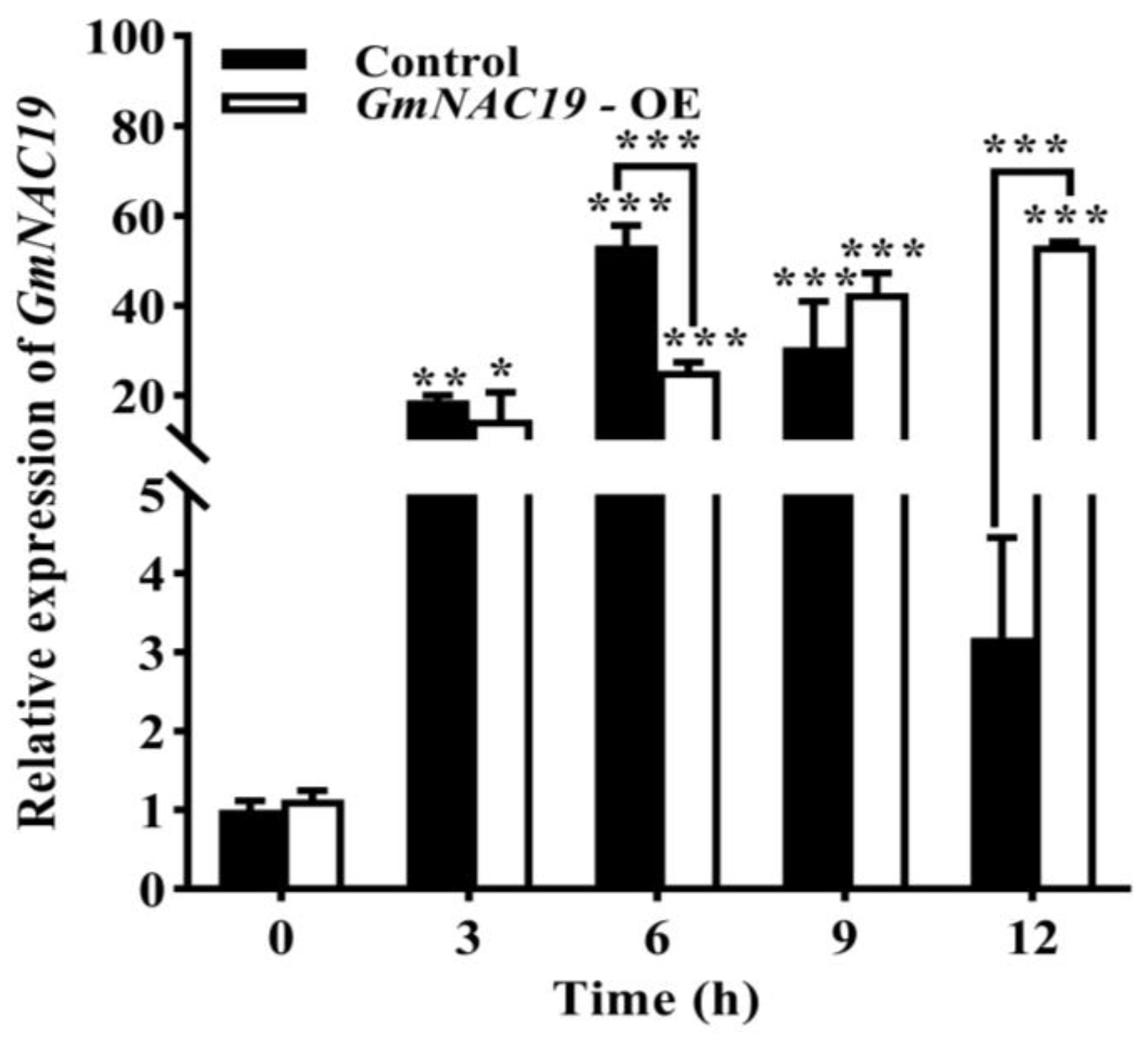
- Gene expression of GmNAC19 in soybean hairy roots under drought stress simulated by 20% PEG6000
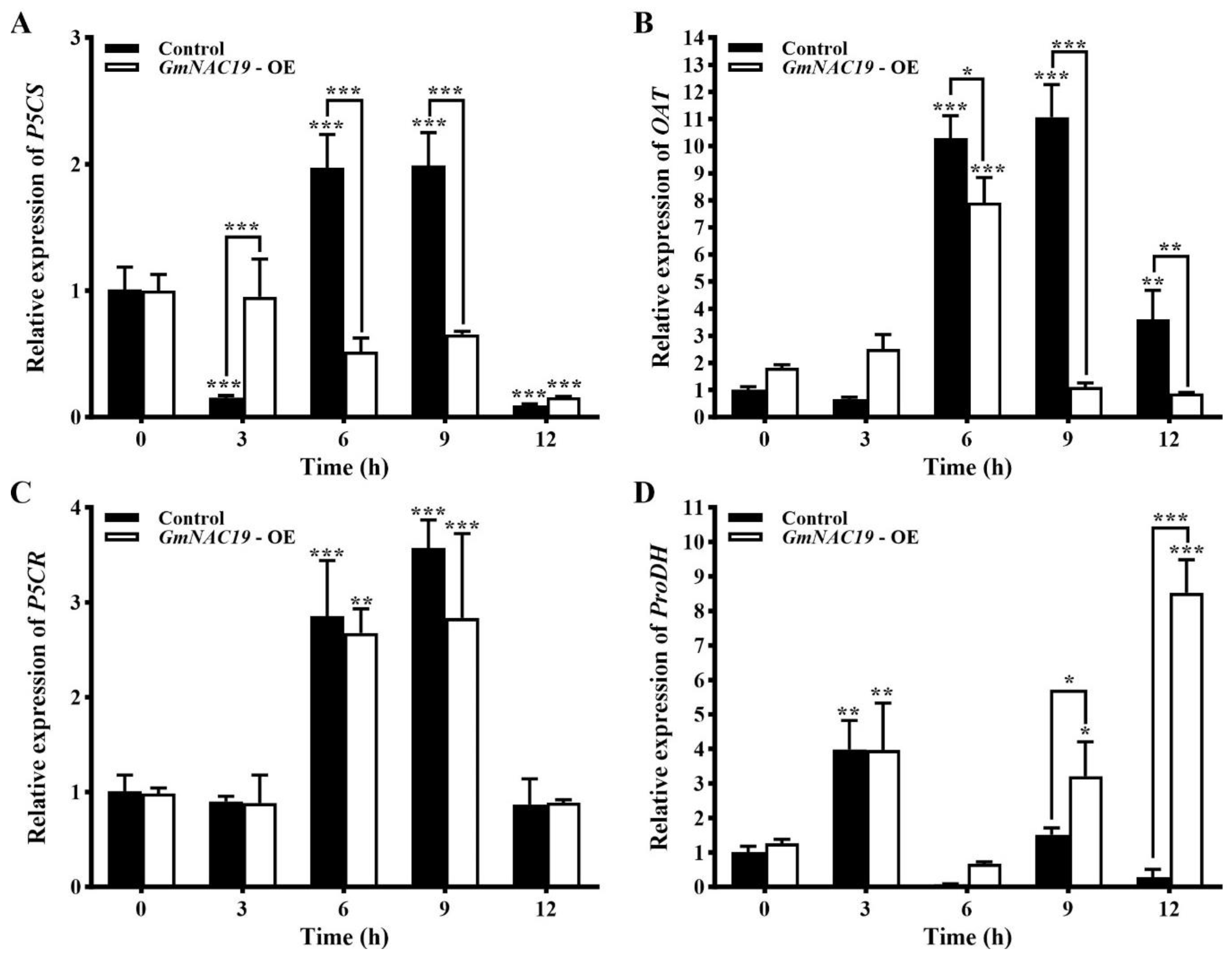
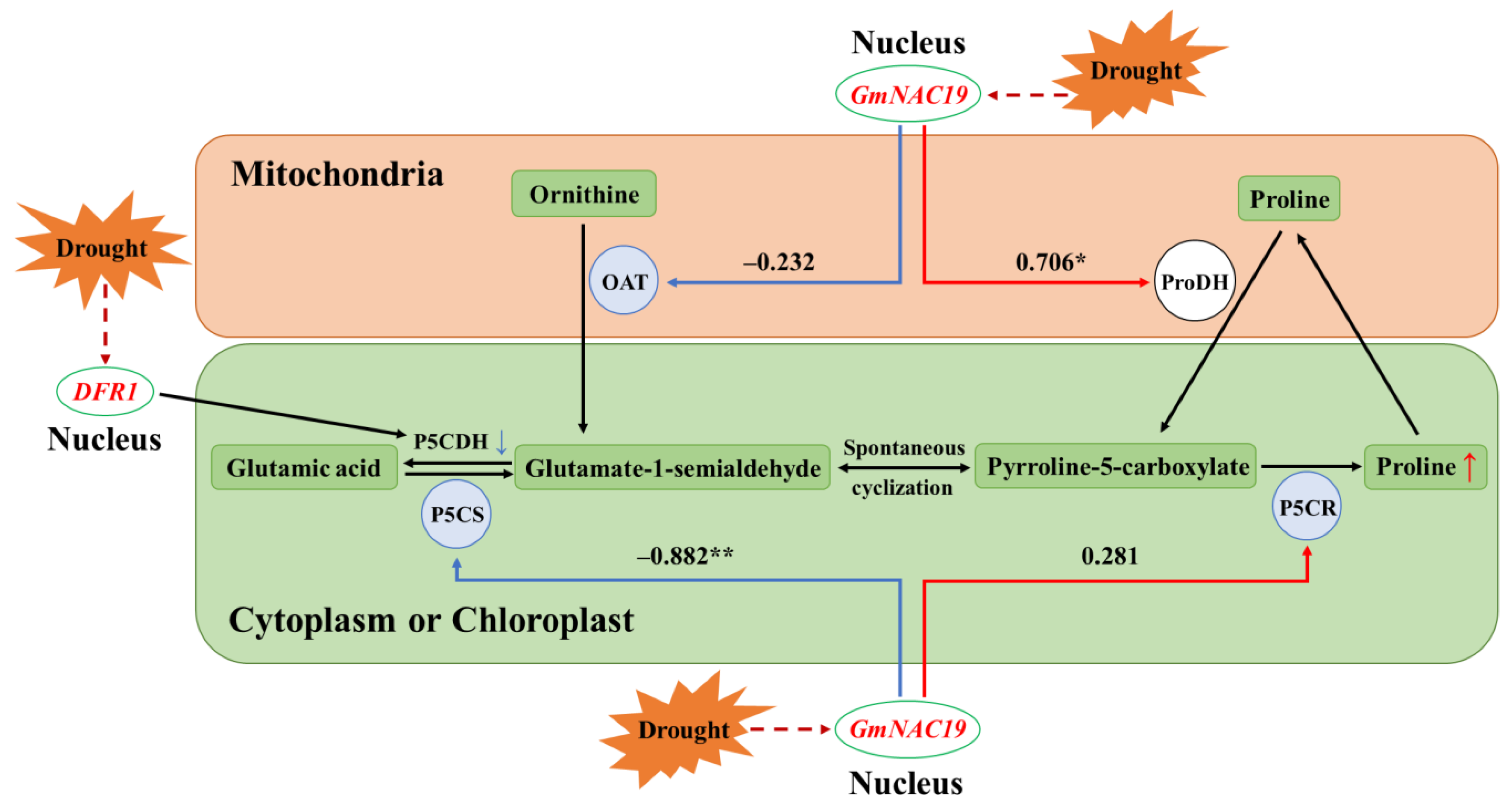
- Expression of four key genes involved in proline metabolic pathway in transgenic soybean hairy roots with GmNAC19 under drought stress simulated by 20% PEG6000
- Correlation analysis between gene expression levels of GmNAC19 and four key genes involved in proline metabolic pathway
3. Discussion
3.1. Gene Expression Patterns of Soybean Transcription Factor GmNAC19
3.2. Enhanced Drought Tolerance Conferred by Soybean Transcription Factor Gene GmNAC19
3.3. Variations in Physiological and Biochemical Indices Related to Drought Tolerance Conferred by the Gene GmNAC19
3.4. Proline Metabolism in Transgenic Soybean and Arabidopsis thaliana with GmNAC19
4. Materials and Methods
4.1. Plant and Microbial Materials
4.2. Cultivation and Treatment of Soybean Materials
4.3. Cultivation and Treatment of Arabidopsis thaliana
4.4. Fluorescence-Based Quantitative Real-Time PCR
4.5. Validation of Drought Resistance in Saccharomyces cerevisiae
4.6. Determination of Physiological and Biochemical Indicators Related to Drought Tolerance
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Q.; Shi, X.; Ai, L.; Tian, X.; Zhang, H.; Tian, J.; Wang, Q.; Zhang, M.; Cui, S.; Yang, C. Genome-wide identification of genes encoding SWI/SNF components in soybean and the functional characterization of GmLFR1 in drought-stressed plants. Front. Plant Sci. 2023, 14, 1176376. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Tripathi, M.K.; Tiwari, S.; Tripathi, N.; Gupta, N.; Sharma, A. Morphological and physiological performance of Indian soybean [Glycine max (L.) Merrill] genotypes in respect to drought. Legume Res. 2021, 105, 1–9. [Google Scholar] [CrossRef]
- Joram, W.; YU, Q.; Ratemo, B. Breeding and genetic engineering of drought-resistant crops. Int. J. Agric. Crop Sci. 2016, 9, 7–12. [Google Scholar]
- Wang, X.; Wu, Z.; Zhou, Q.; Wang, X.; Song, S.; Dong, S. Physiological response of soybean plants to water deficit. Front. Plant Sci. 2022, 12, 809692. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wan, C.; Wang, Y.; Chen, H.; Zhou, Z.; Fu, H.; Sosebee, R.E. The characteristics of Na+, K+ and free proline distribution in several drought-resistant plants of the Alxa desert, China. J. Arid. Environ. 2004, 56, 525–539. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Van Breusegem, F. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef]
- Vadez, V.; Grondin, A.; Chenu, K.; Henry, A.; Laplaze, L.; Millet, E.J.; Carminati, A. Crop traits and production under drought. Nat Rev Earth Environ. 2024. [Google Scholar] [CrossRef]
- Arya, H.; Singh, M.B.; Bhalla, P.L. Towards developing drought-smart soybeans. Front. Plant Sci. 2021, 12, 750664. [Google Scholar] [CrossRef]
- Hoang, X.L.T.; Nhi, D.N.H.; Thu, N.B.A.; Thao, N.P.; Tran, L.S.P. Transcription factors and their roles in signal transduction in plants under abiotic stresses. J. Turbul. 2017, 18, 483–497. [Google Scholar] [CrossRef]
- Hussain, Q.; Asim, M.; Zhang, R.; Khan, R.; Farooq, S.; Wu, J. Transcription factors interact with ABA through gene expression and signaling pathways to mitigate drought and salinity stress. Biomolecules 2021, 11, 1159. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Koyama, H.; Bhati, K.K.; Alok, A. The biotechnological importance of the plant-specific NAC transcription factor family in crop improvement. J. Plant Res. 2021, 134, 475–495. [Google Scholar] [CrossRef]
- Chen, K.; Tang, W.; Zhou, Y.; Chen, J.; Xu, Z.; Ma, R.; Dong, Y.; Ma, Y.; Chen, M. AP2/ERF transcription factor GmDREB1 confers drought tolerance in transgenic soybean by interacting with GmERFs. Plant Physiol. Bioch 2022, 170, 287–295. [Google Scholar] [CrossRef]
- Kerr, T.C.C.; Abdel-Mageed, H.; Aleman, L.; Lee, J.; Payton, P.; Cryer, D.; Allen, R.D. Ectopic expression of two AREB/ABF orthologs increases drought tolerance in cotton (Gossypium hirsutum). Plant Cell Environ. 2018, 41, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Radani, Y.; Li, R.; Korboe, H.M.; Ma, H.; Yang, L. Transcriptional and post-translational regulation of plant bHLH transcription factors during the response to environmental stresses. Plants 2023, 12, 2113. [Google Scholar] [CrossRef]
- Wang, X.; Niu, Y.; Zheng, Y. Multiple functions of MYB transcription factors in abiotic stress responses. Int. J. Mol. Sci. 2021, 22, 6125. [Google Scholar] [CrossRef]
- Qian, C.; Chen, Z.; Liu, Q.; Mao, W.; Chen, Y.; Tian, W.; Liu, Y.; Han, J.; Ouyang, X.; Huang, X. Coordinated transcriptional regulation by the UV-B photoreceptor and multiple transcription factors for plant UV-B responses. Mol. Plant 2020, 13, 777–792. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Y.; Zhang, D.; Tang, X.; Li, Z.; Shen, C.; Han, X.; Deng, W.; Yin, W.; Xia, X. PtrWRKY75 overexpression reduces stomatal aperture and improves drought tolerance by salicylic acid-induced reactive oxygen species accumulation in poplar. Environ. Exp. Bot. 2020, 176, 104117. [Google Scholar] [CrossRef]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Skriver, K. NAC transcription factors: Structurally distinct, functionally diverse. Trends Plant Sci. 2005, 10, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, R.; Jiang, Q.; Sun, X.; Zhang, H.; Hu, Z. GmNAC06, a NAC domain transcription factor enhances salt stress tolerance in soybean. Plant Mol. Biol. 2021, 105, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Munir, N.; Chen, Y.K.; Chen, X.H.; Nawaz, M.A.; Iftikhar, J.; Rizwan, H.M.; Shen, X.; Lin, Y.L.; Xu, X.H.; Lai, Z.X. Genome-wide identification and comprehensive analyses of NAC transcription factor gene family and expression patterns during somatic embryogenesis in Dimocarpus longan Lour. Plant Physiol. Biochem. 2020, 157, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Kjaersgaard, T.; Jensen, M.K.; Christiansen, M.W.; Gregersen, P.; Kragelund, B.B.; Skriver, K. Senescence-associated barley NAC (NAM, ATAF1,2, CUC) transcription factor interacts with radical-induced cell death 1 through a disordered regulatory domain. J. Biol. Chem. 2011, 286, 35418–35429. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, Q.; Xiong, L.; Lou, Z. A structural view of the conserved domain of rice stress-responsive NAC1. Protein Cell 2011, 2, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Seo, P.J.; Kim, S.-G.; Park, C.-M. Membrane-bound transcription factors in plants. Trends Plant Sci. 2008, 13, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Luo, F.; Zhong, Y.; He, J.; Li, L. Modulation of NAC transcription factor NST1 activity by XYLEM NAC DOMAIN 1 regulates secondary cell wall formation in Arabidopsis. J. Exp. Bot. 2020, 71, 1449–1458. [Google Scholar] [CrossRef]
- Ma, J.; Wang, L.Y.; Dai, J.X.; Wang, Y.; Lin, D. The NAC-Type transcription factor CaNAC46 regulates the salt and drought tolerance of transgenic Arabidopsis thaliana. BMC Plant Biol. 2021, 21, 11. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, S.; Ma, X.; Wang, Y.; Kong, F.; Meng, Q. A stress-associated NAC transcription factor (SlNAC35) from tomato plays a positive role in biotic and abiotic stresses. Physiol. Plant 2016, 158, 45–64. [Google Scholar] [CrossRef]
- Jia, D.; Jiang, Q.; Van, N.S.; Gong, X.; Ma, F. An apple (Malus domestica) NAC transcription factor enhances drought tolerance in transgenic apple plants. Plant Physiol. Biochem. 2019, 139, 504–512. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, Y.; Li, B.; Chang, J.; Chen, M.; Li, K.; Yang, G.; He, G. TaNAC29, a NAC transcription factor from wheat, enhances salt and drought tolerance in transgenic Arabidopsis. BMC Plant Biol. 2015, 15, 268. [Google Scholar] [CrossRef]
- Hong, Y.; Zhang, H.; Huang, L.; Li, D.; Song, F. Overexpression of a stress-responsive NAC transcription factor gene ONAC022 improves drought and salt tolerance in rice. Front. Plant Sci. 2016, 7, 4. [Google Scholar] [CrossRef]
- Ju, Y.L.; Yue, X.F.; Min, Z.; Wang, X.H.; Fang, Y.L.; Zhang, J.X. VvNAC17, a novel stress-responsive grapevine (Vitis vinifera L.) NAC transcription factor, increases sensitivity to abscisic acid and enhances salinity, freezing, and drought tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2020, 146, 98–111. [Google Scholar] [CrossRef]
- Yu, X.; Liu, Y.; Wang, S.; Tao, Y.; Wang, Z.; Shu, Y.; Peng, H.; Mijiti, A.; Wang, Z.; Zhang, H. CarNAC4, a NAC-type chickpea transcription factor conferring enhanced drought and salt stress tolerances in Arabidopsis. Plant Cell Rep. 2016, 35, 613–627. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Wang, H.; Cai, J.; Bi, Y.; Li, D.; Song, F. Rice NAC transcription factor ONAC066 functions as a positive regulator of drought and oxidative stress response. BMC Plant Biol. 2019, 19, 278. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yang, X.; Tang, M.; Wang, Y.; Zhang, Q.; Li, H.; Zhou, Y.; Sun, F.; Cui, X. Molecular characterization and drought resistance of GmNAC3 transcription factor in Glycine max (L.) Merr. Int. J. Mol. Sci. 2022, 23, 12378. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y. Cloning and Functional Study of Soybean Transcription Factor GmNAC4 Gene. Master’s Thesis, Jilin Agricultural University, Jilin, China, 2022. [Google Scholar]
- Yang, C.; Huang, Y.; Lv, W.; Zhang, Y.; Bhat, J.A.; Kong, J.; Xing, H.; Zhao, J.; Zhao, T. GmNAC8 acts as a positive regulator in soybean drought stress. Plant Sci. 2020, 293, 110442. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, C.; Li, L.; Reynolds, M.; Mao, X.; Jing, R. Exploitation of drought tolerance-related genes for crop improvement. Int. J. Mol. Sci. 2021, 22, 10265. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, T.; Shinozaki, K. Research on plant abiotic stress responses in the post-genome era: Past, present and future. Plant J. 2010, 61, 1041–1052. [Google Scholar] [CrossRef]
- Nakashima, K.; Takasaki, H.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. NAC transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta 2012, 1819, 97–103. [Google Scholar] [CrossRef]
- Li, D.P.; Peng, S.B.; Chen, S.W.; Li, Z.Y.; Ren, B.; Yang, G.Y. Identification and characterization of 5 walnut MYB genes in response to drought stress involved in ABA signaling. Physiol. Mol. Biol. Plants 2021, 27, 1323–1335. [Google Scholar] [CrossRef]
- Li, H.J.; Chen, J.; Zhao, Q.; Han, Y.L.; Li, L.; Sun, C.Y.; Wang, K.Y.; Wang, Y.F.; Zhao, M.Z.; Chen, P.; et al. Basic leucine zipper (bZIP) transcription factor genes and their response to drought stress in ginseng. Genomics 2021, 22, 316. [Google Scholar] [PubMed]
- Yang, S.Q.; Xu, K.; Chen, S.J.; Li, T.I.; Xia, H.; Chen, L.; Liu, H.Y.; Luo, L.J. A stress-responsive bZIP transcription factor OsbZIP62 improves drought and oxidative tolerance in rice. Plant Biol. 2019, 19, 260. [Google Scholar] [CrossRef] [PubMed]
- Huysmans, M.; Buono, R.; Andrade, S.; Noemi, R.; Marta, C. NAC Transcription factors ANAC087 and ANAC046 control distinct aspects of programmed cell Death in the Arabidopsis columella and kateral root cap. Plant Cell 2018, 30, 2197–2213. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Chai, S.C.; Mcintyre, C.L.; Xue, G.P. Overexpression of a predominantly root-expressed NAC transcription factor in wheat roots enhances root length, biomass and drought tolerance. Plant Cell Rep. 2018, 37, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Zhang, Y.F.; Nie, L.Z. Sunflower HaDREBA5 gene cloning and its response to biotic and abiotic stress. J. Agricul Biotechnol. 2021, 29, 900–914. [Google Scholar]
- Ren, D.Y.; Zhao, H.Y.; Liu, Y.M.; Gao, X.Q.; Sun, Y.D.; Yang, G.Y. Cloning and drought resistance function analysis of walnut translation initiation factor JreIF1A. J. Northwest A&F Univ. 2019, 34, 103–108. [Google Scholar]
- Bian, S.; Jin, D.; Sun, G.; Shan, B.; Zhou, H.; Wang, J.; Zhai, L.; Li, X. Characterization of the soybean R2R3-MYB transcription factor GmMYB81 and its functional roles under abiotic stresses. Gene 2020, 753, 144803. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Dong, Y.; Yang, X.; Guo, D.; Qian, X.; Yan, F.; Wang, Y.; Li, J.; Wang, Q. Functional activation of a novel R2R3-MYB protein gene, GmMYB68, confers salt-alkali resistance in soybean (Glycine max L.). Genome 2020, 63, 13–26. [Google Scholar] [CrossRef]
- Long, H.; He, T.; Farrar, S.; Ji, L.B.; Liu, T.Y.; Ma, X. Antioxidants, aintain cellular redox homeostasis by elimination of reactive oxygen species. Cell Physiol. Biochem. 2017, 44, 532–553. [Google Scholar]
- Ayala, A.; Mu, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Xia, Z.; Cai, Z.; Li, L.; Cheng, Y.; Liu, J.; Nian, H. GmWRKY16 enhances drought and salt tolerance through an ABA-mediated pathway in Arabidopsis thaliana. Front. Plant Sci. 2019, 9, 1979. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.S.; Wang, M.; Gong, X.Q.; Liu, J.H. The transcription factor FcWRKY40 of Fortunella crassifolia functions positively in salt tolerance through modulation of ion homeostasis and proline biosynthesis by directly regulating SOS2 and P5CS1 homologs. New Phytol. 2018, 3, 972–989. [Google Scholar] [CrossRef] [PubMed]
- Lv, K.; Wei, H.; Liu, G.A. R2R3-MYB transcription factor gene, BpMYB123, regulates BpLEA14 to improve drought tolerance in Betula platyphylla. Front. Plant Sci. 2021, 10, 791390. [Google Scholar] [CrossRef]
- Wang, B.; Zhong, Z.; Wang, X.; Han, X.Y.; Yu, D.S.; Wang, C.G.; Song, W.Q.; Zheng, X.L.; Chen, C.B.; Zhang, Y. Knockout of the OsNAC006 transcription factor causes drought and heat sensitivity in rice. Int. J. Mol. Sci. 2020, 21, 2288. [Google Scholar] [CrossRef]
- Wang, F.; Wang, W.J.; Sun, L.J.; Song, S.X. The molecular cloning and functional characterization of ChNAC1, a NAC transcription factor in Cerasus humilis. Plant Growth Regul. 2019, 89, 331–343. [Google Scholar] [CrossRef]
- Mijiti, M.; Wang, Y.C.; Wang, L.Q.; Habuding, X. Tamarix hispida NAC transcription factor ThNAC4 confers salt and drought stress tolerance to transgenic tamarix and Arabidopsis. Plants 2022, 11, 2647. [Google Scholar] [CrossRef]
- Jian, W.; Zheng, Y.X.; Zheng, G.; Yu, T.T.; Cao, H.H.; Chen, Y.; Cui, Q.Y.; Xu, C.; Li, Z.G. SlNAC6, A NAC transcription factor, is involved in drought stress response and reproductive process in tomato. J. Plant Physiol. 2021, 264, 153483. [Google Scholar] [CrossRef]
- Sadeghipour, O. Cadmium toxicity alleviates by seed priming with proline or glycine betaine in cowpea (Vigna unguiculata (L.) Walp.). Egypt. J. Agron. 2020, 42, 163–170. [Google Scholar] [CrossRef]
- Adejumo, S.A.; Oniosun, B.; Akpoilih, O.A.; Adeseko, A.; Arowo, D.O. Anatomical changes, osmolytes accumulation and distribution in the native plants growing on pb-contaminated sites. Environ. Geochem. Health 2021, 43, 1537–1549. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Kumar, V.; Burritt, D.; Fujita, M.; Mäkelä, P. Osmoprotectant-Mediated Abiotic Stress Tolerance in Plants Recent Advances and Future Perspectives: Recent Advances and Future Perspectives; Spring Nature: Cham, Switzerland, 2019; pp. 41–72. [Google Scholar]
- Yin, H. Functional Analysis of Soybean GmRIQ2 Gene under Stress. Master’s Thesis, Northeast Agricultural University, Harbin, China, 2022. [Google Scholar]
- Ren, Y.B.; Miao, M.; Meng, Y.; Cao, J.; Fan, T.; Yue, J.; Xiao, F.; Liu, Y.; Cao, S. DFR1-mediated inhibition of proline degradation pathway regulates drought and freezing tolerance in Arabidopsis. Cell Rep. 2018, 23, 3960–3974. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.P.; Liang, R.Q.; Zhang, L.Q.; Chen, X.Q.; Zhang, X.D.; Wang, G.Y.; Sun, Z.Y. Enhancing the drought resistance of perennial ryegrass by the CBF1 gene, a stress resistant regulatory transcription factor. Acta Agriculturae Boreali-Sinica 2006, 1, 14–18. [Google Scholar]
- Hosseinifard, M.; Stefaniak, S.; Ghorbani, J.M.; Soltani, E.; Wojtyla, Ł.; Garnczarska, M. Contribution of exogenous proline to abiotic stresses tolerance in plants: A review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Székely, G.; Abrahám, E.; Cséplo, A.; Rigó, G.; Zsigmond, L.; Csiszár, J.; Ayaydin, F.; Strizhov, N.; Jásik, J.; Schmelzer, E. Duplicated P5CS genes of Arabidopsis play distinct roles in stress regulation and developmental control of proline biosynthesis. Plant J. 2008, 53, 11–28. [Google Scholar] [CrossRef]
- Sun, S.; Liu, X.; Zhang, T.; Yang, H.; Yu, B. Functional Characterization of the Transcription Factor GsWRKY23 Gene from Glycine soja in Overexpressed Soybean Composite Plants and Arabidopsis under Salt Stress. Plants 2023, 12, 3030. [Google Scholar] [CrossRef]
- Matthews, B.F.; Youssef, R.M. Agrobacterium rhizogenes-Based Transformation of Soybean Roots to Form Composite Plants. Bio-protocol 2016, 6, e1708. [Google Scholar] [CrossRef]
- Wang, Y.J.; Wang, Y.; Zhang, Q.; Fan, H.; Wang, X.; Wang, J.; Zhou, Y.; Chen, Z.; Sun, F.; Cui, X. Saline-alkali soil property improved by the synergistic effects of Priestia aryabhattai JL-5, Staphylococcus pseudoxylosus XW-4, leymus chinensis and soil microbiota. Int. J. Mol. Sci. 2023, 24, 7737. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.W.; Wang, L.; Wen, H.C.; Liu, B.X.; Li, H.J. Physiological and biochemical responses of branches of different mulberry species under low temperature stress. Non-Wood For. Res. 2023, 41, 136–149. [Google Scholar]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transgenic Line | 100-Seed Weight | WT Line | 100-Seed Weight |
|---|---|---|---|
| OE-1 | 2.32 ± 0.02 ** | WT-3 | 1.21 ± 0.01 |
| OE-6 | 3.31 ± 0.03 ** | WT-4 | 1.62 ± 0.02 |
| OE-7 | 2.62 ± 0.03 * | WT-6 | 1.61 ± 0.02 |
| OE-9 | 2.53 ± 0.03 * | WT-7 | 1.61 ± 0.02 |
| OE-12 | 2.21 ± 0.02 * | WT-8 | 1.33 ± 0.01 |
| Group | P5CS | OAT | P5CR | ProDH |
|---|---|---|---|---|
| Control | 0.732 * | 0.791 * | 0.776 * | −0.143 |
| GmNAC19-OE | −0.882 ** | −0.232 | 0.281 | 0.706 * |
| Gene | Coding Sequence (bp) | Length in Amino Acid | Chromosome | NAM Domain Amino Acid Location | POD Activity | CAT Activity | SOD Activity | MDA Content | Chlorophyll Content |
|---|---|---|---|---|---|---|---|---|---|
| GmNAC3 | 1452 | 483 | n/a | 6–132 | + | + | + | − | n/a |
| GmNAC4 | 1650 | 549 | 20 | 20–146 | − | + | + | − | n/a |
| GmNAC8 | 1092 | 363 | n/a | 10–139 | + | n/a | n/a | n/a | n/a |
| GmNAC19 | 807 | 268 | 13 | 10–133 | + | + | + | − | + |
| Primer | Sequence (5′→3′) |
|---|---|
| pYES2-GmNAC19-F | CGTTACTAGTGGATCCATGGCCGCAGCAACACA |
| pYES2-GmNAC19-R | AGGGAATATTAAGCTTTCAGAAGGGCCTGGAGAG |
| GmNAC19-3301-F | TCCAGCTCCAGGATCCATGGCCGCAGCAACACA |
| GmNAC19-3301-R | TCAGAAGGGCCTGGAGAGGAGAAAGCTTGGATCC |
| qGmNAC19-F | ATGGCCGCAGCAACACAACT |
| qGmNAC19-R | ATACCACTCTTTCTCTCCGT |
| qGmEF1A-F | TGCAAAGGAGGCTGCTAACT |
| qGmEF1A-R | CAGCATCACCGTTCTTCAAA |
| qGmP5CS-F | TCACTCGCCAAGATGGAAGG |
| qGmP5CS-R | ACTTGCGGCTTCTGAAGGTC |
| qGmP5CR-F | GGGTTCCGTGGAACACTGAT |
| qGmP5CR-R | AGCTCGAAAAGACTGTTATGGC |
| qGmProDH-F | GGTGTCGACAAAGAGGCTG |
| qGmProDH-R | GCGTCTTCCACACCGTACA |
| qGmδ-OAT-F | AGGGTTTGCAGAGGAAGTAGG |
| qGmδ-OAT-R | CAGAGGTTCCCTTTGCCTGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, X.; Tang, M.; Li, L.; Chang, J.; Yang, X.; Chang, H.; Zhou, J.; Liu, M.; Wang, Y.; Zhou, Y.; et al. Expression Patterns and Molecular Mechanisms Regulating Drought Tolerance of Soybean [Glycine max (L.) Merr.] Conferred by Transcription Factor Gene GmNAC19. Int. J. Mol. Sci. 2024, 25, 2396. https://doi.org/10.3390/ijms25042396
Cui X, Tang M, Li L, Chang J, Yang X, Chang H, Zhou J, Liu M, Wang Y, Zhou Y, et al. Expression Patterns and Molecular Mechanisms Regulating Drought Tolerance of Soybean [Glycine max (L.) Merr.] Conferred by Transcription Factor Gene GmNAC19. International Journal of Molecular Sciences. 2024; 25(4):2396. https://doi.org/10.3390/ijms25042396
Chicago/Turabian StyleCui, Xiyan, Minghao Tang, Lei Li, Jiageng Chang, Xiaoqin Yang, Hongli Chang, Jiayu Zhou, Miao Liu, Yan Wang, Ying Zhou, and et al. 2024. "Expression Patterns and Molecular Mechanisms Regulating Drought Tolerance of Soybean [Glycine max (L.) Merr.] Conferred by Transcription Factor Gene GmNAC19" International Journal of Molecular Sciences 25, no. 4: 2396. https://doi.org/10.3390/ijms25042396