
Targeting PI3K/AKT/mTOR and MAPK Signaling Pathways in Gastric Cancer

 , and
, and
Abstract
:1. Introduction
1.1. Gastric Cancer Incidence
1.2. Gastric Cancer Classification
2. Risk Factors
2.1. Helicobacter pylori
2.2. Diet
2.3. Systemic Diseases
2.4. Genetic Factors
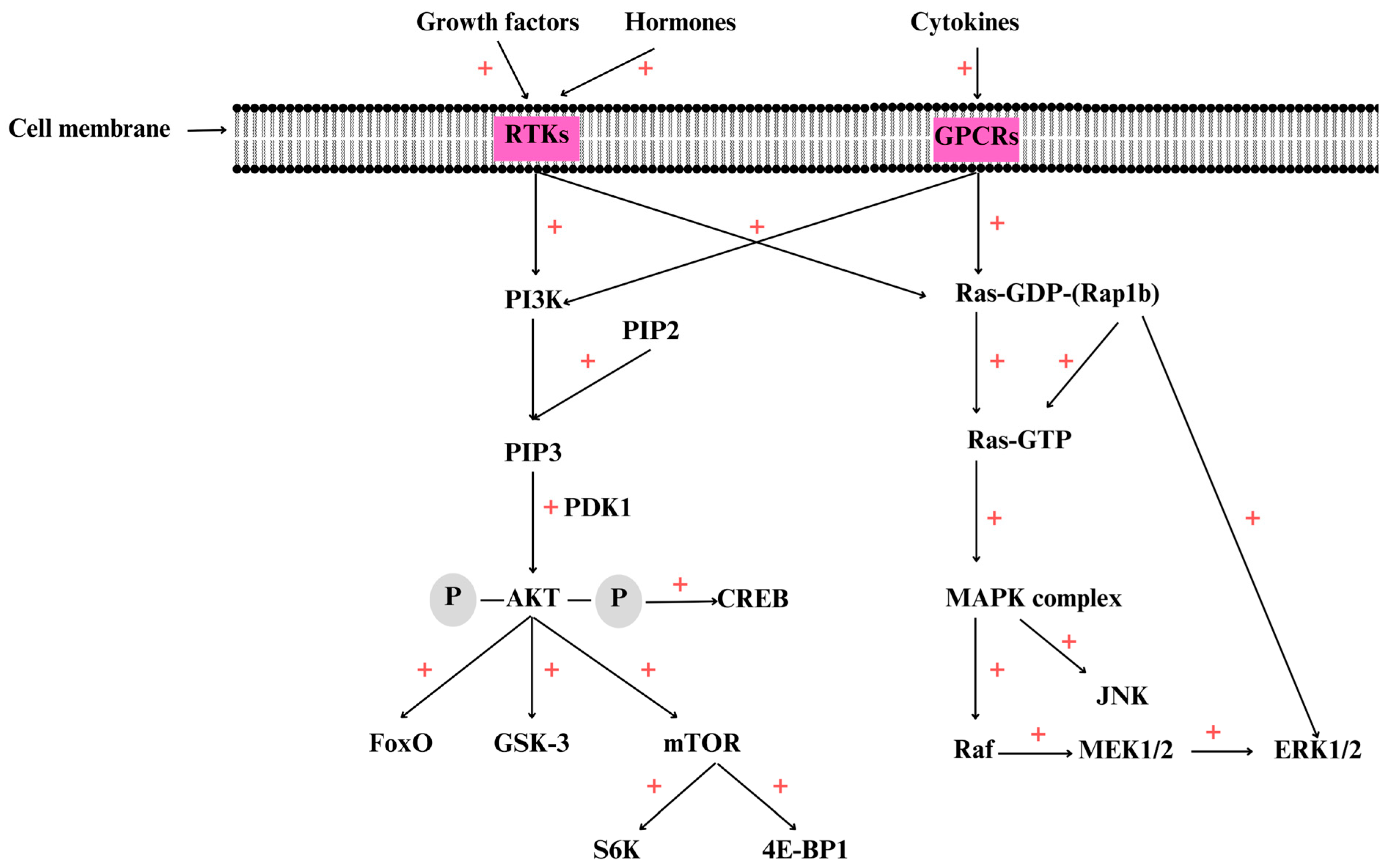
3. Overview of the PI3K/AKT/mTOR Signaling Pathway
4. Overview of the MAPK Signaling Pathway
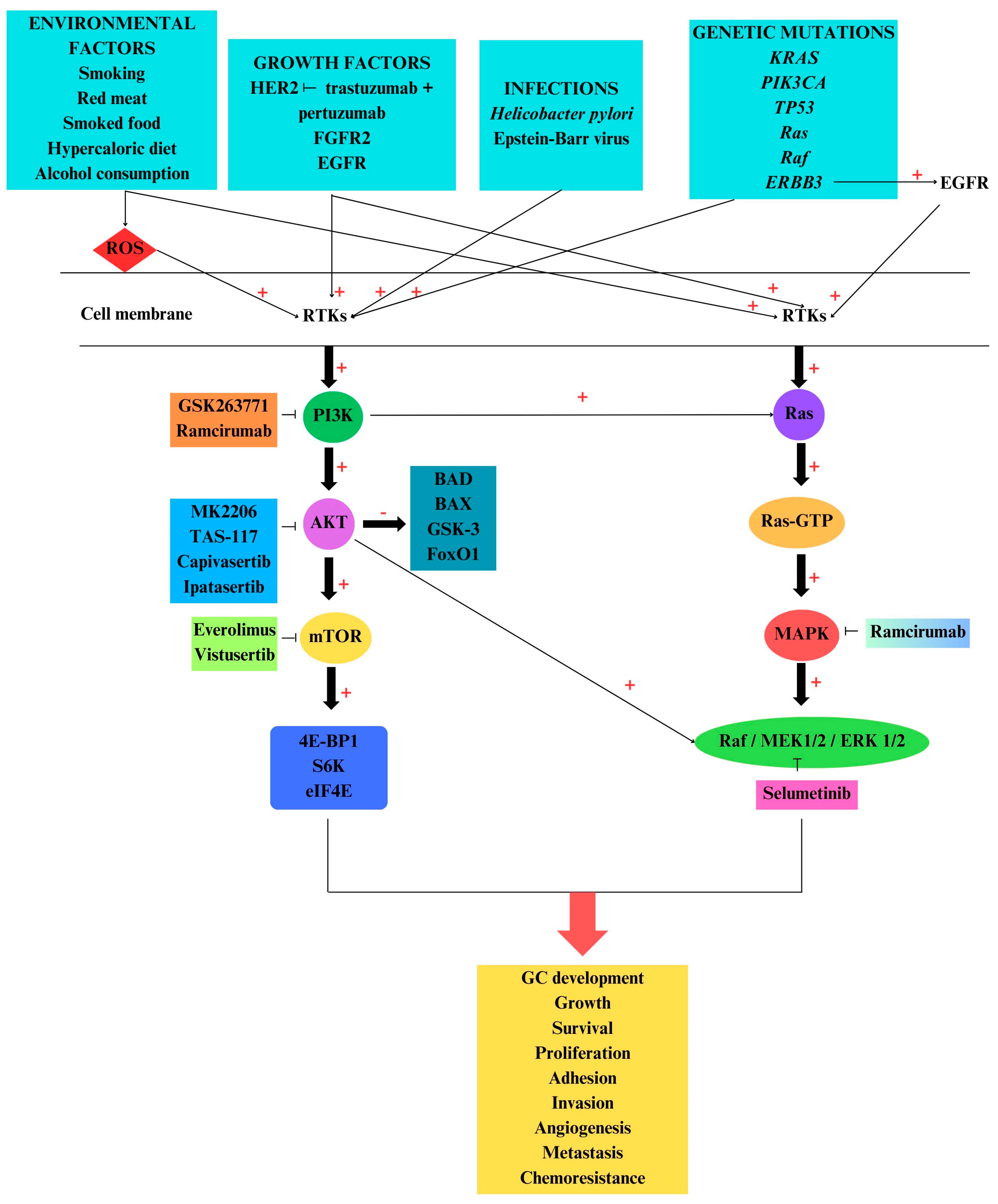
5. PI3K/AKT/mTOR and MAPK Pathways in Gastric Cancer
6. PI3K/AKT/mTOR and MAPK Inhibitors
6.1. Inhibitors Used in Clinical Trials
6.2. Inhibitors Used in In Vitro and In Vivo Studies
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Machlowska, J.; Baj, J.; Sitarz, M.; Maciejewski, R.; Sitarz, R. Gastric cancer: Epidemiology, risk factors, classification, genomic characteristics and treatment strategies. Int. J. Mol. Sci. 2020, 21, 4012. [Google Scholar] [CrossRef] [PubMed]
- Guggenheim, D.E.; Shah, M.A. Gastric cancer epidemiology and risk factors. J. Surg. Oncol. 2013, 107, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Bai, D.; Xiang, W.; Chen, X.; Hu, J. Risk factors of postoperative pulmonary infection of gastric cancer and perioperative intervention measures. Zhonghua Wei Chang. Wai Ke Za Zhi = Chin. J. Gastrointest. Surg. 2021, 24, 185–190. [Google Scholar]
- Yoon, H.; Kim, N. Diagnosis and management of high risk group for gastric cancer. Gut Liver 2015, 9, 5. [Google Scholar] [CrossRef]
- Zhang, K.; Hu, X.; Su, J.; Li, D.; Thakur, A.; Gujar, V.; Cui, H. Gastrointestinal Cancer Therapeutics via Triggering Unfolded Protein Response and Endoplasmic Reticulum Stress by 2-Arylbenzofuran. Int. J. Mol. Sci. 2024, 25, 999. [Google Scholar] [CrossRef]
- Yang, K.; Lu, L.; Liu, H.; Wang, X.; Gao, Y.; Yang, L.; Li, Y.; Su, M.; Jin, M.; Khan, S. A comprehensive update on early gastric cancer: Defining terms, etiology, and alarming risk factors. Expert Rev. Gastroenterol. Hepatol. 2021, 15, 255–273. [Google Scholar] [CrossRef]
- Shah, D.; Bentrem, D. Environmental and genetic risk factors for gastric cancer. J. Surg. Oncol. 2022, 125, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhang, Y.; Ding, M.; Xu, R. Long noncoding RNA TMPO-AS1/miR-126-5p/BRCC3 axis accelerates gastric cancer progression and angiogenesis via activating PI3K/Akt/mTOR pathway. J. Gastroenterol. Hepatol. 2021, 36, 1877–1888. [Google Scholar] [CrossRef]
- Thrift, A.P.; El-Serag, H.B. Burden of gastric cancer. Clin. Gastroenterol. Hepatol. 2020, 18, 534–542. [Google Scholar] [CrossRef]
- Esposito, T.; Pisanti, S.; Mauro, L.; Mencherini, T.; Martinelli, R.; Aquino, R.P. Activity of Colocasia esculenta (Taro) Corms against Gastric Adenocarcinoma Cells: Chemical Study and Molecular Characterization. Int. J. Mol. Sci. 2023, 25, 252. [Google Scholar] [CrossRef]
- Petryszyn, P.; Chapelle, N.; Matysiak-Budnik, T. Gastric cancer: Where are we heading? Dig. Dis. 2020, 38, 280–285. [Google Scholar] [CrossRef]
- Camargo, M.C.; Song, M.; Xu, X.; Zhao, I.; Sampson, J.N.; Etemadi, A.; Brenner, H.; Lee, H.W.; Trabert, B.; Holleczek, B. Urinary estrogen metabolites and gastric cancer risk among postmenopausal women. Cancer Rep. 2022, 5, e1574. [Google Scholar] [CrossRef]
- Imounga, L.M.; Plenet, J.; Belliardo, S.; Chine, E.C.; Louvel, D.; Cenciu, B.; Couppié, P.; Alsibai, K.D.; Nacher, M. Gastric cancer incidence and mortality in French Guiana: South American or French? J. Gastrointest. Cancer 2022, 53, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Gullo, I.; Grillo, F.; Mastracci, L.; Vanoli, A.; Carneiro, F.; Saragoni, L.; Limarzi, F.; Ferro, J.; Parente, P.; Fassan, M. Precancerous lesions of the stomach, gastric cancer and hereditary gastric cancer syndromes. Pathologica 2020, 112, 166. [Google Scholar] [CrossRef] [PubMed]
- Daniyal, M.; Ahmad, S.; Ahmad, M.; Asif, H.M.; Akram, M.; Rehman, S.U.; Sultana, S. Risk factors and epidemiology of gastric cancer in Pakistan. Asian Pac. J. Cancer Prev. 2015, 16, 4821–4824. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kim, N. Gastric cancer and family history. Korean J. Intern. Med. 2016, 31, 1042. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Park, J.Y.; Camargo, M.C.; Lunet, N.; Forman, D.; Soerjomataram, I. Is gastric cancer becoming a rare disease? A global assessment of predicted incidence trends to 2035. Gut 2020, 69, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, Y.; Kato, M.; Asaka, M. Geographic differences in gastric cancer incidence can be explained by differences between Helicobacter pylori strains. Intern. Med. 2008, 47, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Ke, J.; Ma, P.; Chen, J.; Qin, J.; Qian, H. LGR6 promotes the progression of gastric cancer through PI3K/AKT/mTOR pathway. OncoTargets Ther. 2018, 11, 3025–3033. [Google Scholar] [CrossRef]
- Karimi, P.; Islami, F.; Anandasabapathy, S.; Freedman, N.D.; Kamangar, F. Gastric cancer: Descriptive epidemiology, risk factors, screening, and prevention. Cancer Epidemiol. Biomark. Prev. 2014, 23, 700–713. [Google Scholar] [CrossRef]
- Akbarpour, E.; Sadjadi, A.; Derakhshan, M.; Roshandel, G.; Alimohammadian, M. Gastric cancer in Iran: An overview of risk factors and preventive measures. Arch. Iran. Med. 2021, 24, 556–567. [Google Scholar] [CrossRef]
- Thrift, A.P.; Nguyen, T.H. Gastric cancer epidemiology. Gastrointest. Endosc. Clin. 2021, 31, 425–439. [Google Scholar] [CrossRef]
- van Velzen, M.J.; Braemer, M.; Nieuwenhuijzen, G.A.; van Sandick, J.W.; Siersema, P.D.; Ruurda, J.P.; Verheij, M.; Spaander, M.C.; Beerepoot, L.V.; Mohammad, N.H. Incidence, Stage, Treatment, and Survival of Noncardia Gastric Cancer. JAMA Netw. Open 2023, 6, e2330018. [Google Scholar] [CrossRef]
- Xia, J.Y.; Aadam, A.A. Advances in screening and detection of gastric cancer. J. Surg. Oncol. 2022, 125, 1104–1109. [Google Scholar] [CrossRef]
- Crew, K.D.; Neugut, A.I. Epidemiology of gastric cancer. World J. Gastroenterol. WJG 2006, 12, 354. [Google Scholar] [CrossRef]
- Huang, R.J.; Hwang, J.H. Improving the early diagnosis of gastric cancer. Gastrointest. Endosc. Clin. 2021, 31, 503–517. [Google Scholar] [CrossRef]
- Riquelme, I.; Saavedra, K.; Espinoza, J.A.; Weber, H.; García, P.; Nervi, B.; Garrido, M.; Corvalán, A.H.; Roa, J.C.; Bizama, C. Molecular classification of gastric cancer: Towards a pathway-driven targeted therapy. Oncotarget 2015, 6, 24750. [Google Scholar] [CrossRef] [PubMed]
- Chia, N.-Y.; Tan, P. Molecular classification of gastric cancer. Ann. Oncol. 2016, 27, 763–769. [Google Scholar] [CrossRef]
- Moon, S.H.; Park, N.-S.; Noh, M.H.; Kim, Y.S.; Cheong, S.H.; Hur, D.Y. Olaparib-induced Apoptosis Through EBNA1-ATR-p38 MAPK Signaling Pathway in Epstein-Barr Virus-positive Gastric Cancer Cells. Anticancer Res. 2022, 42, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Mezynski, M.J.; Farrelly, A.M.; Cremona, M.; Carr, A.; Morgan, C.; Workman, J.; Armstrong, P.; McAuley, J.; Madden, S.; Fay, J. Targeting the PI3K and MAPK pathways to improve response to HER2-targeted therapies in HER2-positive gastric cancer. J. Transl. Med. 2021, 19, 184. [Google Scholar] [CrossRef] [PubMed]
- Osumi, H.; Kawachi, H.; Yoshio, T.; Fujisaki, J. Clinical impact of Epstein-Barr virus status on the incidence of lymph node metastasis in early gastric cancer. Dig. Endosc. 2020, 32, 316–322. [Google Scholar] [CrossRef]
- Jafarinezhad, S.; Assaran Darban, R.; Javid, H.; Hashemy, S.I. The SP/NK1R system promotes the proliferation of breast cancer cells through NF-κB-mediated inflammatory responses. Cell Biochem. Biophys. 2023, 81, 787–794. [Google Scholar] [CrossRef]
- Puliga, E.; Corso, S.; Pietrantonio, F.; Giordano, S. Microsatellite instability in Gastric Cancer: Between lights and shadows. Cancer Treat. Rev. 2021, 95, 102175. [Google Scholar] [CrossRef]
- Kim, Y.; Cho, M.-Y.; Kim, J.; Kim, S.N.; Oh, S.C.; Lee, K.-A. Profiling cancer-associated genetic alterations and molecular classification of cancer in Korean gastric cancer patients. Oncotarget 2017, 8, 69888. [Google Scholar] [CrossRef]
- Ang, T.L.; Fock, K.M. Clinical epidemiology of gastric cancer. Singap. Med. J. 2014, 55, 621. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.; Li, X.; Gong, M.; Wen, P.; Wen, J.; Xu, L.; Wang, G. SPI1 Mediates N-Myristoyltransferase 1 to Advance Gastric Cancer Progression via PI3K/AKT/mTOR Pathway. Can. J. Gastroenterol. Hepatol. 2023, 2023, 2021515. [Google Scholar] [CrossRef]
- He, P.; He, Y.; Ma, J.; Liu, Y.; Liu, C.; Baoping, Y.; Dong, W. Thymoquinone induces apoptosis and protective autophagy in gastric cancer cells by inhibiting the PI3K/Akt/mTOR pathway. Phytother. Res. 2023, 37, 3467–3480. [Google Scholar] [CrossRef]
- Zhou, D.; He, Y.; Li, H.; Huang, W. KLK6 mediates stemness and metabolism of gastric carcinoma cells via the PI3K/AKT/mTOR signaling pathway. Oncol. Lett. 2021, 22, 824. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liang, G.; Yang, S.; Sui, J.; Wu, W.; Xu, S.; Ye, Y.; Shen, B.; Zhang, X.; Zhang, Y. LncRNA-LOC101928316 contributes to gastric cancer progression through regulating PI3K-Akt-mTOR signaling pathway. Cancer Med. 2019, 8, 4428–4440. [Google Scholar] [CrossRef]
- Lee, Y.-C.; Chiang, T.-H.; Chou, C.-K.; Tu, Y.-K.; Liao, W.-C.; Wu, M.-S.; Graham, D.Y. Association between Helicobacter pylori eradication and gastric cancer incidence: A systematic review and meta-analysis. Gastroenterology 2016, 150, 1113–1124.e1115. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-J.; Zhao, H.-P.; Yu, Y.; Wang, J.-H.; Guo, L.; Liu, J.-Y.; Pu, J.; Lv, J. Updates on global epidemiology, risk and prognostic factors of gastric cancer. World J. Gastroenterol. 2023, 29, 2452. [Google Scholar] [CrossRef]
- den Hoed, C.M.; Kuipers, E.J. Gastric cancer: How can we reduce the incidence of this disease? Curr. Gastroenterol. Rep. 2016, 18, 34. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Yamamichi, N.; Kubota, D.; Shimamoto, T.; Nagao, S.; Sakuma, N.; Sakaguchi, Y.; Yakabi, S.; Tsuji, Y.; Wada, R. Risk factors for gastric cancer in Japan in the 2010s: A large, long-term observational study. Gastric Cancer 2022, 25, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Pasechnikov, V.; Chukov, S.; Fedorov, E.; Kikuste, I.; Leja, M. Gastric cancer: Prevention, screening and early diagnosis. World J. Gastroenterol. WJG 2014, 20, 13842. [Google Scholar] [CrossRef] [PubMed]
- Yang, L. Incidence and mortality of gastric cancer in China. World J. Gastroenterol. WJG 2006, 12, 17. [Google Scholar] [CrossRef] [PubMed]
- Imrie, C.; Rowland, M.; Bourke, B.; Drumm, B. Is Helicobacter pylori infection in childhood a risk factor for gastric cancer? Pediatrics 2001, 107, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Uno, Y. Prevention of gastric cancer by Helicobacter pylori eradication: A review from Japan. Cancer Med. 2019, 8, 3992–4000. [Google Scholar] [CrossRef] [PubMed]
- Abrams, J.A.; Gonsalves, L.; Neugut, A.I. Diverging trends in the incidence of reflux-related and Helicobacter pylori-related gastric cardia cancer. J. Clin. Gastroenterol. 2013, 47, 322. [Google Scholar] [CrossRef] [PubMed]
- Sekiguchi, M.; Oda, I.; Matsuda, T.; Saito, Y. Epidemiological trends and future perspectives of gastric cancer in Eastern Asia. Digestion 2022, 103, 22–28. [Google Scholar] [CrossRef]
- Huang, R.J.; Laszkowska, M.; In, H.; Hwang, J.H.; Epplein, M. Controlling Gastric Cancer in a World of Heterogeneous Risk. Gastroenterology 2023, 164, 736–751. [Google Scholar] [CrossRef]
- Lee, J.; Lim, J.W.; Kim, H. Astaxanthin inhibits matrix metalloproteinase expression by suppressing PI3K/AKT/mTOR activation in Helicobacter pylori-infected gastric epithelial cells. Nutrients 2022, 14, 3427. [Google Scholar] [CrossRef]
- Su, P.; Jiang, L.; Zhang, Y.; Yu, T.; Kang, W.; Liu, Y.; Yu, J. Crosstalk between tumor-associated macrophages and tumor cells promotes chemoresistance via CXCL5/PI3K/AKT/mTOR pathway in gastric cancer. Cancer Cell Int. 2022, 22, 290. [Google Scholar] [CrossRef]
- Liu, G.; Yang, S.; Liu, Y.; Xu, Y.; Qiu, H.; Sun, J.; Song, J.; Shi, L. The adenosine-A2a receptor regulates the radioresistance of gastric cancer via PI3K-AKT-mTOR pathway. Int. J. Clin. Oncol. 2022, 27, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Zheng, Z.-H.; Wan, T.; Wu, J.; Liao, C.-W.; Sun, X.-J. Vitexin inhibits gastric cancer growth and metastasis through HMGB1-mediated inactivation of the PI3K/AKT/mTOR/HIF-1α signaling pathway. J. Gastric Cancer 2021, 21, 439. [Google Scholar] [CrossRef]
- Tay, S.W.; Li, J.W.; Fock, K.M. Diet and cancer of the esophagus and stomach. Curr. Opin. Gastroenterol. 2021, 37, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Yusefi, A.R.; Lankarani, K.B.; Bastani, P.; Radinmanesh, M.; Kavosi, Z. Risk factors for gastric cancer: A systematic review. Asian Pac. J. Cancer Prev. APJCP 2018, 19, 591. [Google Scholar] [PubMed]
- Wu, X.; Chen, L.; Cheng, J.; Qian, J.; Fang, Z.; Wu, J. Effect of dietary salt intake on risk of gastric cancer: A systematic review and meta-analysis of case-control studies. Nutrients 2022, 14, 4260. [Google Scholar] [CrossRef] [PubMed]
- Maddineni, G.; Xie, J.J.; Brahmbhatt, B.; Mutha, P. Diet and carcinogenesis of gastric cancer. Curr. Opin. Gastroenterol. 2022, 38, 588–591. [Google Scholar] [CrossRef] [PubMed]
- Compare, D.; Rocco, A.; Nardone, G. Risk factors in gastric cancer. Eur. Rev. Med. Pharmacol. Sci. 2010, 14, 302–308. [Google Scholar]
- Kim, S.Y.; Kwak, J.H.; Eun, C.S.; Han, D.S.; Kim, Y.S.; Song, K.S.; Choi, B.Y.; Kim, H.J. Gastric Cancer Risk Was Associated with Dietary Factors Irritating the Stomach Wall: A Case–Control Study in Korea. Nutrients 2022, 14, 2233. [Google Scholar] [CrossRef]
- Yuan, P.; Lin, L.; Zheng, K.; Wang, W.; Wu, S.; Huang, L.; Wu, B.; Chen, T.; Li, X.; Cai, L. Risk factors for gastric cancer and related serological levels in Fujian, China: Hospital-based case–control study. BMJ Open 2020, 10, e042341. [Google Scholar] [CrossRef]
- Conti, C.B.; Agnesi, S.; Scaravaglio, M.; Masseria, P.; Dinelli, M.E.; Oldani, M.; Uggeri, F. Early gastric cancer: Update on prevention, diagnosis and treatment. Int. J. Environ. Res. Public Health 2023, 20, 2149. [Google Scholar] [CrossRef]
- Hu, K.; Wang, S.; Wang, Z.; Li, L.; Huang, Z.; Yu, W.; Chen, Z.; Wu, Q.-F. Clinicopathological risk factors for gastric cancer: A retrospective cohort study in China. BMJ Open 2019, 9, e030639. [Google Scholar] [CrossRef]
- Sun, D.; Zhang, M.; Wei, M.; Wang, Z.; Qiao, W.; Liu, P.; Zhong, X.; Liang, Y.; Chen, Y.; Huang, Y. Ox-LDL-mediated ILF3 overexpression in gastric cancer progression by activating the PI3K/AKT/mTOR signaling pathway. Aging (Albany NY) 2022, 14, 3887. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhang, J.; Zhou, Y.; Qiao, L. Obesity and gastric cancer. Front. Biosci.-Landmark 2012, 17, 2383–2390. [Google Scholar] [CrossRef]
- Pih, G.Y.; Gong, E.J.; Choi, J.Y.; Kim, M.-J.; Ahn, J.Y.; Choe, J.; Bae, S.E.; Chang, H.-S.; Na, H.K.; Lee, J.H. Associations of serum lipid level with gastric cancer risk, pathology, and prognosis. Cancer Res. Treat. Off. J. Korean Cancer Assoc. 2021, 53, 445–456. [Google Scholar] [CrossRef]
- Hui, Y.; Tu, C.; Liu, D.; Zhang, H.; Gong, X. Risk factors for gastric cancer: A comprehensive analysis of observational studies. Front. Public Health 2023, 10, 892468. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Li, X.; Ding, S.; Dai, D. Adherence to the Mediterranean Diet and Risk of Gastric Cancer: A Systematic Review and Meta-Analysis. Nutrients 2023, 15, 3826. [Google Scholar] [CrossRef]
- Liu, A.R.; He, Q.S.; Wu, W.H.; Du, J.L.; Kuo, Z.C.; Xia, B.; Tang, Y.; Yun, P.; Cheung, E.C.; Tang, Y.Z. Body composition and risk of gastric cancer: A population-based prospective cohort study. Cancer Med. 2021, 10, 2164–2174. [Google Scholar] [CrossRef]
- Akbari, A.; Ashtari, S.; Tabaiean, S.P.; Mehrdad-Majd, H.; Farsi, F.; Shojaee, S.; Agah, S. Overview of epidemiological characteristics, clinical features, and risk factors of gastric cancer in Asia-Pacific region. Asia-Pac. J. Clin. Oncol. 2022, 18, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Venerito, M.; Link, A.; Rokkas, T.; Malfertheiner, P. Gastric cancer–clinical and epidemiological aspects. Helicobacter 2016, 21, 39–44. [Google Scholar] [CrossRef]
- Kim, S.R.; Kim, K.; Lee, S.A.; Kwon, S.O.; Lee, J.-K.; Keum, N.; Park, S.M. Effect of red, processed, and white meat consumption on the risk of gastric cancer: An overall and dose–response meta-analysis. Nutrients 2019, 11, 826. [Google Scholar] [CrossRef]
- Wu, K.; Liu, L.; Shu, T.; Li, A.; Xia, D.; Sun, X. The relationship between processed meat, red meat, and risk of types of cancer: A Mendelian randomization study. Front. Nutr. 2022, 9, 942155. [Google Scholar] [CrossRef]
- Na, H.-K.; Lee, J.Y. Molecular basis of alcohol-related gastric and colon cancer. Int. J. Mol. Sci. 2017, 18, 1116. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Eshak, E.S.; Shirai, K.; Liu, K.; JY, D.; Iso, H.; Tamakoshi, A.; Group, J.S. Alcohol consumption and risk of gastric cancer: The Japan collaborative cohort study. J. Epidemiol. 2021, 31, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Carchman, E. Crohn’s disease and the risk of cancer. Clin. Colon Rectal Surg. 2019, 32, 305–313. [Google Scholar] [CrossRef]
- Mihai, D.A.; Stefan, D.S.; Stegaru, D.; Bernea, G.E.; Vacaroiu, I.A.; Papacocea, T.; Lupușoru, M.O.D.; Nica, A.E.; Stiru, O.; Dragos, D. Continuous glucose monitoring devices: A brief presentation. Exp. Ther. Med. 2022, 23, 174. [Google Scholar] [CrossRef]
- Tian, T.; Zhang, L.; Ma, X.; Zhou, J.; Shen, J. Diabetes mellitus and incidence and mortality of gastric cancer: A meta-analysis. Exp. Clin. Endocrinol. Diabetes 2011, 120, 217–223. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, Z. Diabetes Mellitus and Gastric Cancer: Correlation and Potential Mechanisms. J. Diabetes Res. 2023, 2023, 4388437. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J. Insulin resistance: A hidden risk factor for gastric cancer? Gut Liver 2019, 13, 133. [Google Scholar] [CrossRef]
- Wang, T.; Zhan, R.; Lu, J.; Zhong, L.; Peng, X.; Wang, M.; Tang, S. Grain consumption and risk of gastric cancer: A meta-analysis. Int. J. Food Sci. Nutr. 2020, 71, 164–175. [Google Scholar] [CrossRef]
- Gaesser, G.A. Whole grains, refined grains, and cancer risk: A systematic review of meta-analyses of observational studies. Nutrients 2020, 12, 3756. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Zhang, J.; Li, D.; Liu, Y.; Zhang, D.L.; Liu, C.F.; Wang, N.; Wu, S.R.; Lu, W.Q.; Guo, J.Z. Bile reflux is an independent risk factor for precancerous gastric lesions and gastric cancer: An observational cross-sectional study. J. Dig. Dis. 2021, 22, 282–290. [Google Scholar] [CrossRef]
- Malekzadeh, R.; Derakhshan, M.H.; Malekzadeh, Z. Gastric cancer in Iran: Epidemiology and risk factors. Arch. Iran. Med. 2009, 12, 576–583. [Google Scholar]
- Zeng, S.-X.; Liang, Y.-P.; Wu, X.-Y.; Tan, S.-W.; Liang, Q.; Wen, Z.-F.; Tao, J. Gastroesophageal reflux is associated with an increased risk of gastric cardiac polyps: A case-control study of 140 cases. Ann. Palliat. Med. 2021, 10, 7173–7183. [Google Scholar] [CrossRef]
- Blair, V.R.; McLeod, M.; Carneiro, F.; Coit, D.G.; D’Addario, J.L.; van Dieren, J.M.; Harris, K.L.; Hoogerbrugge, N.; Oliveira, C.; van der Post, R.S. Hereditary diffuse gastric cancer: Updated clinical practice guidelines. Lancet Oncol. 2020, 21, e386–e397. [Google Scholar] [CrossRef]
- Mankaney, G.; Macaron, C.; Burke, C.A. Refining risk factors for gastric cancer in patients with Lynch syndrome to optimize surveillance esophagogastroduodenoscopy. Clin. Gastroenterol. Hepatol. 2020, 18, 780–782. [Google Scholar] [CrossRef] [PubMed]
- Boland, C.R.; Yurgelun, M.B.; Mraz, K.A.; Boland, P.M. Managing gastric cancer risk in lynch syndrome: Controversies and recommendations. Fam. Cancer 2022, 21, 75–78. [Google Scholar] [CrossRef] [PubMed]
- van Beek, E.J.; Hernandez, J.M.; Goldman, D.A.; Davis, J.L.; McLaughlin, K.; Ripley, R.T.; Kim, T.S.; Tang, L.H.; Hechtman, J.F.; Zheng, J. Rates of TP53 mutation are significantly elevated in African American patients with gastric cancer. Ann. Surg. Oncol. 2018, 25, 2027–2033. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Xiong, X.; Han, L.; Zhang, L.; Gao, S.; Liu, L.; Wang, X.; Huang, C.; Tong, D.; Yang, J. SERPINA5 promotes tumour cell proliferation by modulating the PI3K/AKT/mTOR signalling pathway in gastric cancer. J. Cell. Mol. Med. 2022, 26, 4837–4846. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Shao, L.; Wang, Y.; Meng, Q.; Yu, J. Ketamine exhibits anti-gastric cancer activity via induction of apoptosis and attenuation of PI3K/Akt/mTOR. Arch. Med. Sci. 2020, 15, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Panneerpandian, P.; Ganesan, K. PI3K/AKT/mTOR inhibitors as potential extracellular matrix modulators for targeting EMT subtype gastric tumors. Med. Oncol. 2023, 40, 120. [Google Scholar] [CrossRef] [PubMed]
- Vara, J.Á.F.; Casado, E.; de Castro, J.; Cejas, P.; Belda-Iniesta, C.; González-Barón, M. PI3K/Akt signalling pathway and cancer. Cancer Treat. Rev. 2004, 30, 193–204. [Google Scholar] [CrossRef]
- Tewari, D.; Patni, P.; Bishayee, A.; Sah, A.N.; Bishayee, A. Natural products targeting the PI3K-Akt-mTOR signaling pathway in cancer: A novel therapeutic strategy. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2022; pp. 1–17. [Google Scholar]
- Shorning, B.Y.; Dass, M.S.; Smalley, M.J.; Pearson, H.B. The PI3K-AKT-mTOR pathway and prostate cancer: At the crossroads of AR, MAPK, and WNT signaling. Int. J. Mol. Sci. 2020, 21, 4507. [Google Scholar] [CrossRef]
- Xu, Z.; Han, X.; Ou, D.; Liu, T.; Li, Z.; Jiang, G.; Liu, J.; Zhang, J. Targeting PI3K/AKT/mTOR-mediated autophagy for tumor therapy. Appl. Microbiol. Biotechnol. 2020, 104, 575–587. [Google Scholar] [CrossRef]
- Akbarzadeh, M.; Mihanfar, A.; Akbarzadeh, S.; Yousefi, B.; Majidinia, M. Crosstalk between miRNA and PI3K/AKT/mTOR signaling pathway in cancer. Life Sci. 2021, 285, 119984. [Google Scholar] [CrossRef] [PubMed]
- Ratti, S.; Mauro, R.; Rocchi, C.; Mongiorgi, S.; Ramazzotti, G.; Gargiulo, M.; Manzoli, L.; Cocco, L.; Fiume, R. Roles of PI3K/AKT/mTOR Axis in Arteriovenous Fistula. Biomolecules 2022, 12, 350. [Google Scholar] [CrossRef]
- Sun, K.; Luo, J.; Guo, J.; Yao, X.; Jing, X.; Guo, F. The PI3K/AKT/mTOR signaling pathway in osteoarthritis: A narrative review. Osteoarthr. Cartil. 2020, 28, 400–409. [Google Scholar] [CrossRef]
- Li, Q.; Li, Z.; Luo, T.; Shi, H. Targeting the PI3K/AKT/mTOR and RAF/MEK/ERK pathways for cancer therapy. Mol. Biomed. 2022, 3, 47. [Google Scholar] [CrossRef]
- Glaviano, A.; Foo, A.S.; Lam, H.Y.; Yap, K.C.; Jacot, W.; Jones, R.H.; Eng, H.; Nair, M.G.; Makvandi, P.; Geoerger, B. PI3K/AKT/mTOR signaling transduction pathway and targeted therapies in cancer. Mol. Cancer 2023, 22, 138. [Google Scholar] [CrossRef]
- Basile, M.S.; Cavalli, E.; McCubrey, J.; Hernández-Bello, J.; Muñoz-Valle, J.F.; Fagone, P.; Nicoletti, F. The PI3K/Akt/mTOR pathway: A potential pharmacological target in COVID-19. Drug Discov. Today 2022, 27, 848–856. [Google Scholar] [CrossRef]
- Li, H.; Prever, L.; Hirsch, E.; Gulluni, F. Targeting PI3K/AKT/mTOR signaling pathway in breast cancer. Cancers 2021, 13, 3517. [Google Scholar] [CrossRef]
- Nepstad, I.; Hatfield, K.J.; Grønningsæter, I.S.; Reikvam, H. The PI3K-Akt-mTOR signaling pathway in human acute myeloid leukemia (AML) cells. Int. J. Mol. Sci. 2020, 21, 2907. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Pandolfi, P.P. The PTEN–PI3K axis in cancer. Biomolecules 2019, 9, 153. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Du, R.; Liu, W.; Huang, G.; Dong, Z.; Li, X. PI3K/Akt/mTOR signaling pathway: Role in esophageal squamous cell carcinoma, regulatory mechanisms and opportunities for targeted therapy. Front. Oncol. 2022, 12, 852383. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Wang, Y.; Zhou, C.; Mei, W.; Zeng, C. PI3K/Akt/mTOR pathway and its role in cancer therapeutics: Are we making headway? Front. Oncol. 2022, 12, 819128. [Google Scholar] [CrossRef] [PubMed]
- Bertacchini, J.; Heidari, N.; Mediani, L.; Capitani, S.; Shahjahani, M.; Ahmadzadeh, A.; Saki, N. Targeting PI3K/AKT/mTOR network for treatment of leukemia. Cell. Mol. Life Sci. 2015, 72, 2337–2347. [Google Scholar] [CrossRef] [PubMed]
- Hermida, M.A.; Kumar, J.D.; Leslie, N.R. GSK3 and its interactions with the PI3K/AKT/mTOR signalling network. Adv. Biol. Regul. 2017, 65, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Haybaeck, J.; Yang, Z. Therapeutic potential of PI3K/AKT/mTOR pathway in gastrointestinal stromal tumors: Rationale and progress. Cancers 2020, 12, 2972. [Google Scholar] [CrossRef] [PubMed]
- Fattahi, S.; Khalifehzadeh-Esfahani, Z.; Mohammad-Rezaei, M.; Mafi, S.; Jafarinia, M. PI3K/Akt/mTOR pathway: A potential target for anti-SARS-CoV-2 therapy. Immunol. Res. 2022, 70, 269–275. [Google Scholar] [CrossRef]
- Zhu, K.; Wu, Y.; He, P.; Fan, Y.; Zhong, X.; Zheng, H.; Luo, T. PI3K/AKT/mTOR-targeted therapy for breast cancer. Cells 2022, 11, 2508. [Google Scholar] [CrossRef]
- Li, X.; Wu, C.; Chen, N.; Gu, H.; Yen, A.; Cao, L.; Wang, E.; Wang, L. PI3K/Akt/mTOR signaling pathway and targeted therapy for glioblastoma. Oncotarget 2016, 7, 33440. [Google Scholar] [CrossRef] [PubMed]
- Porta, C.; Paglino, C.; Mosca, A. Targeting PI3K/Akt/mTOR signaling in cancer. Front. Oncol. 2014, 4, 64. [Google Scholar] [CrossRef] [PubMed]
- Iksen; Pothongsrisit, S.; Pongrakhananon, V. Targeting the PI3K/AKT/mTOR signaling pathway in lung cancer: An update regarding potential drugs and natural products. Molecules 2021, 26, 4100. [Google Scholar] [CrossRef] [PubMed]
- du Rusquec, P.; Blonz, C.; Frenel, J.S.; Campone, M. Targeting the PI3K/Akt/mTOR pathway in estrogen-receptor positive HER2 negative advanced breast cancer. Ther. Adv. Med. Oncol. 2020, 12, 1758835920940939. [Google Scholar] [CrossRef] [PubMed]
- Pungsrinont, T.; Kallenbach, J.; Baniahmad, A. Role of PI3K-AKT-mTOR pathway as a pro-survival signaling and resistance-mediating mechanism to therapy of prostate cancer. Int. J. Mol. Sci. 2021, 22, 11088. [Google Scholar] [CrossRef] [PubMed]
- Vander Broek, R.; Mohan, S.; Eytan, D.; Chen, Z.; Van Waes, C. The PI 3 K/A kt/m TOR axis in head and neck cancer: Functions, aberrations, cross-talk, and therapies. Oral Dis. 2015, 21, 815–825. [Google Scholar] [CrossRef]
- Zheng, W.; Wu, C.; Wu, X.; Cai, Y.; Liu, B.; Wang, C. Genetic variants of autophagy-related genes in the PI3K/Akt/mTOR pathway and risk of gastric cancer in the Chinese population. Gene 2021, 769, 145190. [Google Scholar] [CrossRef]
- Matsuoka, T.; Yashiro, M. The role of PI3K/Akt/mTOR signaling in gastric carcinoma. Cancers 2014, 6, 1441–1463. [Google Scholar] [CrossRef]
- Alves, C.L.; Ditzel, H.J. Drugging the PI3K/AKT/mTOR pathway in ER+ breast cancer. Int. J. Mol. Sci. 2023, 24, 4522. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, Y.; Li, P.; Jian, J.; Zhao, C.; Wen, G. Mitogen-activated protein kinase and substrate identification in plant growth and development. Int. J. Mol. Sci. 2022, 23, 2744. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- González-Coronel, J.M.; Rodríguez-Alonso, G.; Guevara-García, Á.A. A phylogenetic study of the members of the MAPK and MEK families across Viridiplantae. PLoS ONE 2021, 16, e0250584. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.Y.; Richardson, B.C. The MAPK signalling pathways and colorectal cancer. Lancet Oncol. 2005, 6, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Ronkina, N.; Gaestel, M. MAPK-activated protein kinases: Servant or partner? Annu. Rev. Biochem. 2022, 91, 505–540. [Google Scholar] [CrossRef] [PubMed]
- Burotto, M.; Chiou, V.L.; Lee, J.M.; Kohn, E.C. The MAPK pathway across different malignancies: A new perspective. Cancer 2014, 120, 3446–3456. [Google Scholar] [CrossRef]
- Degirmenci, U.; Wang, M.; Hu, J. Targeting aberrant RAS/RAF/MEK/ERK signaling for cancer therapy. Cells 2020, 9, 198. [Google Scholar] [CrossRef]
- Park, H.-B.; Baek, K.-H. E3 ligases and deubiquitinating enzymes regulating the MAPK signaling pathway in cancers. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2022, 1877, 188736. [Google Scholar] [CrossRef]
- Bardwell, A.J.; Frankson, E.; Bardwell, L. Selectivity of docking sites in MAPK kinases. J. Biol. Chem. 2009, 284, 13165–13173. [Google Scholar] [CrossRef]
- Mesquita, F.P.; Moreira-Nunes, C.A.; da Silva, E.L.; Lima, L.B.; Daniel, J.P.; Zuerker, W.J.; Brayner, M.; de Moraes, M.E.A.; Montenegro, R.C. MAPK14 (p38α) inhibition effects against metastatic gastric cancer cells: A potential biomarker and pharmacological target. Toxicol. In Vitro 2020, 66, 104839. [Google Scholar] [CrossRef]
- Kim, E.K.; Choi, E.-J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2010, 1802, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Mlakar, V.; Morel, E.; Mlakar, S.J.; Ansari, M.; Gumy-Pause, F. A review of the biological and clinical implications of RAS-MAPK pathway alterations in neuroblastoma. J. Exp. Clin. Cancer Res. 2021, 40, 189. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Xia, Y.; Lv, J.; Li, B.; Li, Y.; Wang, S.; Xuan, Z.; Xie, L.; Qiu, S.; He, Z. A novel protein encoded by circMAPK1 inhibits progression of gastric cancer by suppressing activation of MAPK signaling. Mol. Cancer 2021, 20, 66. [Google Scholar] [CrossRef]
- Inamdar, G.S.; Madhunapantula, S.V.; Robertson, G.P. Targeting the MAPK pathway in melanoma: Why some approaches succeed and other fail. Biochem. Pharmacol. 2010, 80, 624–637. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Shi, F.; Cheng, L.; She, J. Knockdown of Rap1b enhances apoptosis and autophagy in gastric cancer cells via the PI3K/Akt/mTOR pathway. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2016, 24, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Kwong, A.J.; Scheidt, K.A. Non-‘classical’MEKs: A review of MEK3-7 inhibitors. Bioorganic Med. Chem. Lett. 2020, 30, 127203. [Google Scholar] [CrossRef]
- Fehrenbacher, N.; Bar-Sagi, D.; Philips, M. Ras/MAPK signaling from endomembranes. Mol. Oncol. 2009, 3, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Molina, J.R.; Adjei, A.A. The ras/raf/mapk pathway. J. Thorac. Oncol. 2006, 1, 7–9. [Google Scholar] [CrossRef]
- Tasharrofi, B.; Ghafouri-Fard, S. Long non-coding RNAs as regulators of the mitogen-activated protein kinase (MAPK) pathway in cancer. Klin. Onkol. 2018, 31, 95–102. [Google Scholar] [CrossRef]
- Fattahi, S.; Amjadi-Moheb, F.; Tabaripour, R.; Ashrafi, G.H.; Akhavan-Niaki, H. PI3K/AKT/mTOR signaling in gastric cancer: Epigenetics and beyond. Life Sci. 2020, 262, 118513. [Google Scholar] [CrossRef]
- Zhao, W.; Zhao, X.; Xu, M.; Cheng, Z.; Zhang, Z. Knockdown of LINC01279 Suppresses Gastric Cancer Proliferation and Migration by Inhibiting PI3K/Akt/mTOR Signaling Pathway. J. Oncol. 2022, 2022, 6228982. [Google Scholar] [CrossRef]
- Gu, X.-J.; Li, Y.-J.; Wang, F.; Ye, T. MiR-30e-3p inhibits gastric cancer development by negatively regulating THO complex 2 and PI3K/AKT/mTOR signaling. World J. Gastrointest. Oncol. 2022, 14, 2170. [Google Scholar] [CrossRef]
- Li, H.; Zhao, S.; Shen, L.; Wang, P.; Liu, S.; Ma, Y.; Liang, Z.; Wang, G.; Lv, J.; Qiu, W. E2F2 inhibition induces autophagy via the PI3K/Akt/mTOR pathway in gastric cancer. Aging (Albany NY) 2021, 13, 13626. [Google Scholar] [CrossRef]
- Wang, N.; Dong, Q.; Zhou, X.-N. LMO4 promotes the invasion and proliferation of gastric cancer by activating PI3K-Akt-mTOR signaling. Am. J. Transl. Res. 2019, 11, 6534. [Google Scholar]
- Wang, R.; Ha, K.-Y.; Dhandapani, S.; Kim, Y.-J. Biologically synthesized black ginger-selenium nanoparticle induces apoptosis and autophagy of AGS gastric cancer cells by suppressing the PI3K/Akt/mTOR signaling pathway. J. Nanobiotechnol. 2022, 20, 441. [Google Scholar] [CrossRef]
- Khorasani, A.B.S.; Pourbagheri-Sigaroodi, A.; Pirsalehi, A.; Safaroghli-Azar, A.; Zali, M.R.; Bashash, D. The PI3K/Akt/mTOR signaling pathway in gastric cancer; from oncogenic variations to the possibilities for pharmacologic interventions. Eur. J. Pharmacol. 2021, 898, 173983. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.; Fujishita, T. Oncogenic roles of the PI3K/AKT/mTOR axis. Viruses Genes Cancer 2017, 407, 153–189. [Google Scholar]
- Kang, B.W.; Chau, I. Molecular target: Pan-AKT in gastric cancer. ESMO Open 2020, 5, e000728. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, X.; Xu, Y.; Li, J.; Zhang, X.; Peng, Z.; Hu, Y.; Zhao, X.; Dong, K.; Zhang, B. Mutations of PI3K-AKT-mTOR pathway as predictors for immune cell infiltration and immunotherapy efficacy in dMMR/MSI-H gastric adenocarcinoma. BMC Med. 2022, 20, 133. [Google Scholar] [CrossRef]
- Koh, V.; Chakrabarti, J.; Torvund, M.; Steele, N.; Hawkins, J.A.; Ito, Y.; Wang, J.; Helmrath, M.A.; Merchant, J.L.; Ahmed, S.A. Hedgehog transcriptional effector GLI mediates mTOR-Induced PD-L1 expression in gastric cancer organoids. Cancer Lett. 2021, 518, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Han, S.-H.; Lee, J.-H.; Woo, J.-S.; Jung, G.-H.; Jung, S.-H.; Han, E.-J.; Kim, B.; Cho, S.D.; Nam, J.S.; Che, J.H. Myricetin induces apoptosis and autophagy in human gastric cancer cells through inhibition of the PI3K/Akt/mTOR pathway. Heliyon 2022, 8, e09309. [Google Scholar] [CrossRef] [PubMed]
- Rong, L.; Li, Z.; Leng, X.; Li, H.; Ma, Y.; Chen, Y.; Song, F. Salidroside induces apoptosis and protective autophagy in human gastric cancer AGS cells through the PI3K/Akt/mTOR pathway. Biomed. Pharmacother. 2020, 122, 109726. [Google Scholar] [CrossRef] [PubMed]
- Xia, P.; Xu, X.-Y. PI3K/Akt/mTOR signaling pathway in cancer stem cells: From basic research to clinical application. Am. J. Cancer Res. 2015, 5, 1602. [Google Scholar] [PubMed]
- Yu, L.; Wei, J.; Liu, P. Attacking the PI3K/Akt/mTOR signaling pathway for targeted therapeutic treatment in human cancer. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2022; pp. 69–94. [Google Scholar]
- Stanland, L.J.; Ang, H.X.; Hoj, J.P.; Chu, Y.; Tan, P.; Wood, K.C.; Luftig, M.A. CBF-Beta Mitigates PI3K-Alpha–Specific Inhibitor Killing through PIM1 in PIK3CA-Mutant Gastric Cancer. Mol. Cancer Res. 2023, 21, 1148–1162. [Google Scholar] [CrossRef] [PubMed]
- Iranpour, M.; Nourian, M.; Saffari, S.; Samizadeh, E.; Mirghafori, M.; Iravani, S.; Ghafouri-Fard, S. PIK3CA mutation analysis in iranian patients with gastric cancer. Iran. Biomed. J. 2019, 23, 87. [Google Scholar] [CrossRef]
- Kipkeeva, F.; Muzaffarova, T.; Korotaeva, A.; Nikulin, M.; Grishina, K.; Mansorunov, D.; Apanovich, P.; Karpukhin, A. MicroRNA in gastric cancer development: Mechanisms and biomarkers. Diagnostics 2020, 10, 891. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yang, Z.; Xu, E.; Shen, X.; Wang, X.; Li, Z.; Yu, H.; Chen, K.; Hu, Q.; Xia, X. Apolipoprotein C-II induces EMT to promote gastric cancer peritoneal metastasis via PI3K/AKT/mTOR pathway. Clin. Transl. Med. 2021, 11, e522. [Google Scholar] [CrossRef]
- Yousefi, B.; Mohammadlou, M.; Abdollahi, M.; Salek Farrokhi, A.; Karbalaei, M.; Keikha, M.; Kokhaei, P.; Valizadeh, S.; Rezaiemanesh, A.; Arabkari, V. Epigenetic changes in gastric cancer induction by Helicobacter pylori. J. Cell. Physiol. 2019, 234, 21770–21784. [Google Scholar] [CrossRef]
- Seidlitz, T.; Schmäche, T.; Lee, J.H.; Qin, N.; Kochall, S.; Fohgrub, J.; Pauck, D.; Rothe, A.; Koo, B.K.; Weitz, J. Sensitivity towards HDAC inhibition is associated with RTK/MAPK pathway activation in gastric cancer. EMBO Mol. Med. 2022, 14, e15705. [Google Scholar] [CrossRef]
- Lin, Y.; Wu, Z.; Guo, W.; Li, J. Gene mutations in gastric cancer: A review of recent next-generation sequencing studies. Tumor Biol. 2015, 36, 7385–7394. [Google Scholar] [CrossRef]
- Shi, W.; Zhang, G.; Ma, Z.; Li, L.; Liu, M.; Qin, L.; Yu, Z.; Zhao, L.; Liu, Y.; Zhang, X. Hyperactivation of HER2-SHCBP1-PLK1 axis promotes tumor cell mitosis and impairs trastuzumab sensitivity to gastric cancer. Nat. Commun. 2021, 12, 2812. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Hormazabal, P.; Musleh, M.; Bustamante, M.; Stambuk, J.; Pisano, R.; Valladares, H.; Lanzarini, E.; Chiong, H.; Rojas, J.; Suazo, J. Polymorphisms in RAS/RAF/MEK/ERK pathway are associated with gastric cancer. Genes 2018, 10, 20. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Brant, R.; Sharpe, A.; Dry, J.R.; Hodgson, D.R.; Kilgour, E.; Kim, K.; Kim, S.T.; Park, S.H.; Kang, W.K. Correlation between MEK signature and Ras gene alteration in advanced gastric cancer. Oncotarget 2017, 8, 107492. [Google Scholar] [CrossRef] [PubMed]
- Ning, G.; Zhu, Q.; Kang, W.; Lee, H.; Maher, L.; Suh, Y.-S.; Michaud, M.; Silva, M.; Kwon, J.Y.; Zhang, C. A novel treatment strategy for lapatinib resistance in a subset of HER2-amplified gastric cancer. BMC Cancer 2021, 21, 923. [Google Scholar] [CrossRef] [PubMed]
- Lian, S.; Li, S.; Zhu, J.; Xia, Y.; Do Jung, Y. Nicotine stimulates IL-8 expression via ROS/NF-κB and ROS/MAPK/AP-1 axis in human gastric cancer cells. Toxicology 2022, 466, 153062. [Google Scholar] [CrossRef]
- Zhu, Q.; Guo, Y.; Chen, S.; Fu, D.; Li, Y.; Li, Z.; Ni, C. Irinotecan induces autophagy-dependent apoptosis and positively regulates ROS-related JNK-and P38-MAPK pathways in gastric cancer cells. OncoTargets Ther. 2020, 13, 2807–2817. [Google Scholar] [CrossRef]
- Shimizu, T.; Tolcher, A.; Papadopoulos, K.; Beeram, M.; Rasco, D.; Smith, L.; Gunn, S.; Smetzer, L.; Mays, T.; Kaiser, B. The clinical effect of the dual-targeting strategy involving PI3K/AKT/mTOR and RAS/MEK/ERK pathways in first-in-human phase I study: The START Center experience. J. Clin. Oncol. 2011, 29, 2502. [Google Scholar] [CrossRef]
- Jiang, C.; Ma, Z.; Zhang, G.; Yang, X.; Du, Q.; Wang, W. CSNK2A1 promotes gastric cancer invasion through the PI3K-Akt-mTOR signaling pathway. Cancer Manag. Res. 2019, 11, 10135–10143. [Google Scholar] [CrossRef]
- Arachchige Don, A.S.; Steven Zheng, X.F. Recent clinical trials of mTOR-targeted cancer therapies. Rev. Recent Clin. Trials 2011, 6, 24–35. [Google Scholar] [CrossRef]
- Apicella, M.; Corso, S.; Giordano, S. Targeted therapies for gastric cancer: Failures and hopes from clinical trials. Oncotarget 2017, 8, 57654. [Google Scholar] [CrossRef]
- Guan, W.-L.; He, Y.; Xu, R.-H. Gastric cancer treatment: Recent progress and future perspectives. J. Hematol. Oncol. 2023, 16, 57. [Google Scholar] [CrossRef]
- Hudis, C.; Swanton, C.; Janjigian, Y.Y.; Lee, R.; Sutherland, S.; Lehman, R.; Chandarlapaty, S.; Hamilton, N.; Gajria, D.; Knowles, J. A phase 1 study evaluating the combination of an allosteric AKT inhibitor (MK-2206) and trastuzumab in patients with HER2-positive solid tumors. Breast Cancer Res. 2013, 15, R110. [Google Scholar] [CrossRef]
- Akt Inhibitor MK2206, Lapatinib Ditosylate, and Trastuzumab in Treating Patients with Locally Advanced or Metastatic HER2-Positive Breast, Gastric, or Gastroesophageal Cancer That Cannot Be Removed By Surgery. ClinicalTrials.gov ID NCT01705340. Available online: https://clinicaltrials.gov/study/NCT01705340?cond=Gastric%20Cancer&term=PI3K%20inhibitors&limit=25&page=1&rank=3&tab=history (accessed on 9 December 2023).
- Ramanathan, R.K.; McDonough, S.L.; Kennecke, H.F.; Iqbal, S.; Baranda, J.C.; Seery, T.E.; Lim, H.J.; Hezel, A.F.; Vaccaro, G.M.; Blanke, C.D. Phase 2 study of MK-2206, an allosteric inhibitor of AKT, as second-line therapy for advanced gastric and gastroesophageal junction cancer: A SWOG cooperative group trial (S 1005). Cancer 2015, 121, 2193–2197. [Google Scholar] [CrossRef]
- Doi, T.; Muro, K.; Boku, N.; Yamada, Y.; Nishina, T.; Takiuchi, H.; Komatsu, Y.; Hamamoto, Y.; Ohno, N.; Fujita, Y. Multicenter phase II study of everolimus in patients with previously treated metastatic gastric cancer. J. Clin. Oncol. 2010, 28, 1904–1910. [Google Scholar] [CrossRef]
- Ohtsu, A.; Ajani, J.A.; Bai, Y.-X.; Bang, Y.-J.; Chung, H.-C.; Pan, H.-M.; Sahmoud, T.; Shen, L.; Yeh, K.-H.; Chin, K. Everolimus for previously treated advanced gastric cancer: Results of the randomized, double-blind, phase III GRANITE-1 study. J. Clin. Oncol. 2013, 31, 3935. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.-C.; Li, C.-P.; Yen, C.-J.; Hsu, C.; Lin, Y.-L.; Lin, Z.-Z.; Chen, L.-T.; Su, W.-C.; Chao, Y.; Yeh, K.-H. Phase II multicentered study of low-dose everolimus plus cisplatin and weekly 24-hour infusion of high-dose 5-fluorouracil and leucovorin as first-line treatment for patients with advanced gastric cancer. Oncology 2014, 87, 104–113. [Google Scholar] [CrossRef] [PubMed]
- RAD001 in Combination with Capecitabine and Oxaliplatin (XELOX) in Patients with Advanced Gastric Cancer: RAD001+XELOX. ClinicalTrials.gov ID NCT01049620. Available online: https://clinicaltrials.gov/study/NCT01049620?cond=Gastric%20Cancer&term=PI3K%20inhibitors&rank=2&tab=history (accessed on 9 December 2023).
- Santini, D.; Vincenzi, B.; Schiavon, G.; Di Seri, M.; Virzí, V.; Spalletta, B.; Caricato, M.; Coppola, R.; Tonini, G. Chronomodulated administration of oxaliplatin plus capecitabine (XELOX) as first line chemotherapy in advanced colorectal cancer patients: Phase II study. Cancer Chemother. Pharmacol. 2007, 59, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wei, Y.; Sun, X.; Liu, M.; Zhu, M.; Yuan, Y.; Zhang, J.; Dong, Y.; Hu, K.; Ma, S. JUNB mediates oxaliplatin resistance via the MAPK signaling pathway in gastric cancer by chromatin accessibility and transcriptomic analysis: JUNB mediates oxaliplatin resistance in gastric cancer. Acta Biochim. Biophys. Sin. 2023, 55, 1784. [Google Scholar] [CrossRef] [PubMed]
- Teng, F.; Xu, Z.; Chen, J.; Zheng, G.; Zheng, G.; Lv, H.; Wang, Y.; Wang, L.; Cheng, X. DUSP1 induces apatinib resistance by activating the MAPK pathway in gastric cancer. Oncol. Rep. 2018, 40, 1203–1222. [Google Scholar] [CrossRef]
- Lee, J.B.; Jung, M.; Beom, S.H.; Kim, G.M.; Kim, H.R.; Choi, H.J.; Sohn, J.H.; Ahn, J.B.; Rha, S.Y.; Chung, H.C. Phase 2 study of TAS-117, an allosteric akt inhibitor in advanced solid tumors harboring phosphatidylinositol 3-kinase/v-akt murine thymoma viral oncogene homolog gene mutations. Investig. New Drugs 2021, 39, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- PI3K Inhibitor BYL719 in Combination with the HSP90 Inhibitor AUY922 in Patients with Advanced or Metastatic Gastric Cancer. ClinicalTrials.gov ID NCT01613950. Available online: https://clinicaltrials.gov/study/NCT01613950?cond=Gastric%20Cancer&term=PI3K%20inhibitors&rank=1#more-information (accessed on 9 December 2023).
- PI3Kβ Selective Inhibitor with Paclitaxel, Advanced Gastric Adenocarcinoma ClinicalTrials.gov ID NCT02615730. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT02615730 (accessed on 9 December 2023).
- PANGEA-IMBBP: Personalized Antibodies for Gastro-Esophageal Adenocarcinoma—A 1st Pilot Metastatic Trial of Biologics beyond Progression. NCT02213289. Available online: https://clinicaltrials.gov/study/NCT02213289 (accessed on 9 December 2023).
- Lupușoru, G.; Ailincăi, I.; Frățilă, G.; Ungureanu, O.; Andronesi, A.; Lupușoru, M.; Banu, M.; Văcăroiu, I.; Dina, C.; Sinescu, I. Tumor lysis syndrome: An endless challenge in onco-nephrology. Biomedicines 2022, 10, 1012. [Google Scholar] [CrossRef]
- Lee, J.; Kim, S.T.; Kim, K.; Lee, H.; Kozarewa, I.; Mortimer, P.G.; Odegaard, J.I.; Harrington, E.A.; Lee, J.; Lee, T. Tumor genomic profiling guides patients with metastatic gastric cancer to targeted treatment: The VIKTORY umbrella trial. Cancer Discov. 2019, 9, 1388–1405. [Google Scholar] [CrossRef]
- Bang, Y.-J.; Kang, Y.-K.; Ng, M.; Chung, H.C.; Wainberg, Z.; Gendreau, S.; Chan, W.; Xu, N.; Maslyar, D.; Meng, R. A phase II, randomised study of mFOLFOX6 with or without the Akt inhibitor ipatasertib in patients with locally advanced or metastatic gastric or gastroesophageal junction cancer. Eur. J. Cancer 2019, 108, 17–24. [Google Scholar] [CrossRef]
- Kang, Y.; Rha, S.; Tassone, P.; Barriuso, J.; Yu, R.; Szado, T.; Garg, A.; Bang, Y. A phase IIa dose-finding and safety study of first-line pertuzumab in combination with trastuzumab, capecitabine and cisplatin in patients with HER2-positive advanced gastric cancer. Br. J. Cancer 2014, 111, 660–666. [Google Scholar] [CrossRef]
- Al-Batran, S.E.; Ducreux, M.; Ohtsu, A. mTOR as a therapeutic target in patients with gastric cancer. Int. J. Cancer 2012, 130, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Li, R.; Wei, P.-F.; Ou, L.; Li, M.; Bai, Y.; Luo, W.-J.; Fan, Z. Synergistic anticancer effects of everolimus (RAD001) and Rhein on gastric cancer cells via phosphoinositide-3-kinase (PI3K)/protein kinase B (AKT)/mammalian target of rapamycin (mTOR) pathway. Bioengineered 2022, 13, 6332–6342. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Lin, X.; Zhang, C.; Liu, Z.; Chen, Z.; Li, Z.; Wang, J.; Li, B.; Hu, Y.; Dong, B. Dual PI3K/mTOR inhibitor BEZ235 as a promising therapeutic strategy against paclitaxel-resistant gastric cancer via targeting PI3K/Akt/mTOR pathway. Cell Death Dis. 2018, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.S.; Yap, W.N.; Arfuso, F.; Kar, S.; Wang, C.; Cai, W.; Dharmarajan, A.M.; Sethi, G.; Kumar, A.P. Targeting the PI3K/Akt signaling pathway in gastric carcinoma: A reality for personalized medicine? World J. Gastroenterol. 2015, 21, 12261. [Google Scholar] [CrossRef] [PubMed]
- Bei, S.; Li, F.; Li, H.; Li, J.; Zhang, X.; Sun, Q.; Feng, L. Inhibition of gastric cancer cell growth by a PI3K-mTOR dual inhibitor GSK1059615. Biochem. Biophys. Res. Commun. 2019, 511, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Tang, J.; Fan, S.; Su, H.; Yu, R.; Zhang, Y.; Wu, H.; Lv, Y.; Zhang, S.; Zou, X. ABBV-744 induces autophagy in gastric cancer cells by regulating PI3K/AKT/mTOR/p70S6k and MAPK signaling pathways. Neoplasia 2023, 45, 100936. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Chen, S.; Wang, Z.; Li, S.; Fei, X.; Wang, T.; Zhang, M. KIRREL promotes the proliferation of gastric cancer cells and angiogenesis through the PI3K/AKT/mTOR pathway. J. Cell. Mol. Med. 2023, 28, e18020. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Venkatarame Gowda Saralamma, V.; Kim, S.M.; Ha, S.E.; Raha, S.; Lee, W.S.; Kim, E.H.; Lee, S.J.; Heo, J.D.; Kim, G.S. Pectolinarigenin induced cell cycle arrest, autophagy, and apoptosis in gastric cancer cell via PI3K/AKT/mTOR signaling pathway. Nutrients 2018, 10, 1043. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.-M.; Wang, X.-F.; Huang, Q.-X. Thymoquinone induces cytotoxicity and reprogramming of EMT in gastric cancer cells by targeting PI3K/Akt/mTOR pathway. J. Biosci. 2017, 42, 547–554. [Google Scholar] [CrossRef]
- Chen, M.; Lu, J.; Wei, W.; Lv, Y.; Zhang, X.; Yao, Y.; Wang, L.; Ling, T.; Zou, X. Effects of proton pump inhibitors on reversing multidrug resistance via downregulating V-ATPases/PI3K/Akt/mTOR/HIF-1α signaling pathway through TSC1/2 complex and Rheb in human gastric adenocarcinoma cells in vitro and in vivo. OncoTargets Ther. 2018, 11, 6705–6722. [Google Scholar] [CrossRef]
- Yao, C.; Liu, J.; Shao, L. Rapamycin inhibits the proliferation and apoptosis of gastric cancer cells by down regulating the expression of survivin. Hepato-Gastroenterol. 2011, 58, 1075–1080. [Google Scholar]
- Wang, R.Y.; Chen, X.W.; Zhang, W.W.; Jiang, F.; Liu, M.Q.; Shen, X.B. CYP2E1 changes the biological function of gastric cancer cells via the PI3K/Akt/mTOR signaling pathway. Mol. Med. Rep. 2020, 21, 842–850. [Google Scholar] [CrossRef]
- Liu, J.-Z.; Hu, Y.-L.; Feng, Y.; Guo, Y.-B.; Liu, Y.-F.; Yang, J.-L.; Mao, Q.-S.; Xue, W.-J. Rafoxanide promotes apoptosis and autophagy of gastric cancer cells by suppressing PI3K/Akt/mTOR pathway. Exp. Cell Res. 2019, 385, 111691. [Google Scholar] [CrossRef]
- Yuan, C.-X.; Zhou, Z.-W.; Yang, Y.-X.; He, Z.-X.; Zhang, X.; Wang, D.; Yang, T.; Pan, S.-Y.; Chen, X.-W.; Zhou, S.-F. Danusertib, a potent pan-Aurora kinase and ABL kinase inhibitor, induces cell cycle arrest and programmed cell death and inhibits epithelial to mesenchymal transition involving the PI3K/Akt/mTOR-mediated signaling pathway in human gastric cancer AGS and NCI-N78 cells. Drug Des. Dev. Ther. 2015, 9, 1293–1318. [Google Scholar]
- Li, L.; Zhang, S.; Xie, D.; Chen, H.; Zheng, X.; Pan, D. Dual inhibitor of PI3K and mTOR (NVP-BEZ235) augments the efficacy of fluorouracil on gastric cancer chemotherapy. OncoTargets Ther. 2018, 11, 6111–6118. [Google Scholar] [CrossRef]
- Kim, M.-Y.; Kruger, A.J.; Jeong, J.-Y.; Kim, J.; kyung Shin, P.; Kim, S.Y.; Cho, J.Y.; Hahm, K.B.; Hong, S.P. Combination Therapy with a PI3K/mTOR Dual Inhibitor and Chloroquine Enhances Synergistic Apoptotic Cell Death in Epstein–Barr Virus-Infected Gastric Cancer Cells. Mol. Cells 2019, 42, 448. [Google Scholar] [CrossRef]
- Wu, Y.-J.; Lin, S.-H.; Din, Z.-H.; Su, J.-H.; Liu, C.-I. Sinulariolide inhibits gastric cancer cell migration and invasion through downregulation of the EMT process and suppression of FAK/PI3K/AKT/mTOR and MAPKs signaling pathways. Mar. Drugs 2019, 17, 668. [Google Scholar] [CrossRef]
- Yang, L.; Liu, Y.-N.; Gu, Y.; Guo, Q. Deltonin enhances gastric carcinoma cell apoptosis and chemosensitivity to cisplatin via inhibiting PI3K/AKT/mTOR and MAPK signaling. World J. Gastrointest. Oncol. 2023, 15, 1739–1755. [Google Scholar] [CrossRef]
- Fukuoka, S.; Koga, Y.; Yamauchi, M.; Koganemaru, S.; Yasunaga, M.; Shitara, K.; Doi, T.; Yoshino, T.; Kuronita, T.; Elenbaas, B. p70S6K/Akt dual inhibitor DIACC3010 is efficacious in preclinical models of gastric cancer alone and in combination with trastuzumab. Sci. Rep. 2023, 13, 16017. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.-Q.; Zhang, T.; Xue, H.; Zuo, W.-B.; Li, Y.-N.; Zhao, Y.; Sun, G.; Fu, Z.-R.; Zhang, Q. Calycosin Induces Gastric Cancer Cell Apoptosis via the ROS-Mediated MAPK/STAT3/NF-κB Pathway. OncoTargets Ther. 2021, 14, 2505–2517. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.Q.; Feng, Y.Y.; Luo, Y.H.; Zhai, Y.Q.; Ju, X.Y.; Feng, Y.C.; Wang, J.R.; Yu, C.Q.; Jin, C.H. Glycitein induces reactive oxygen species-dependent apoptosis and G0/G1 cell cycle arrest through the MAPK/STAT3/NF-κB pathway in human gastric cancer cells. Drug Dev. Res. 2019, 80, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ren, F.; Tian, Z.; Song, W.; Cheng, B.; Feng, Z. Osthole synergizes with HER2 inhibitor, trastuzumab in HER2-overexpressed N87 gastric cancer by inducing apoptosis and inhibition of AKT-MAPK pathway. Front. Pharmacol. 2018, 9, 1392. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Shi, L.; Ma, H.; Li, H.; Li, Y.; Lu, Y.; Wang, Q.; Li, W. Dihydroartemisinin induces apoptosis in human gastric cancer cell line BGC-823 through activation of JNK1/2 and p38 MAPK signaling pathways. J. Recept. Signal Transduct. 2017, 37, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.-N.; Luo, Y.-H.; Liu, S.-B.; Xu, W.-T.; Zhang, Y.; Zhang, T.; Xue, H.; Zuo, W.-B.; Li, Y.-N.; Wang, C.-Y. Zeaxanthin induces apoptosis via ROS-regulated MAPK and AKT signaling pathway in human gastric cancer cells. OncoTargets Ther. 2020, 13, 10995–11006. [Google Scholar] [CrossRef]
- Capasso, A.; Gambardella, V.; Martinelli, E.; Troiani, T.; Nappi, A.; Sforza, V.; Manzo, A.; Martini, G.; Della Corte, C.; Napolitano, S. Cell Growth Inhibition of Her2 Positive Trastuzumab Resistant Gastric Cancer Cell Lines By Combined Inhibition of Pi3K/Akt/Mtor and Mapk. Ann. Oncol. 2014, 25, iv220. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Trial Type | Drug | Target | Reference |
|---|---|---|---|
| Phase I | Trastuzumab | HER2 | [174] |
| MK-2206 | AKT 1 and AKT 2 | ||
| Phase I | MK-2206+ | AKT | [175] |
| lapatinib ditosylate + trastuzumab | HER2 | ||
| Phase 2 | MK-2206 | AKT | [176] |
| Phase II | Everolimus (RAD 0001) | mTOR | [177] |
| International, double-blind, phase III | Everolimus (RAD001) | mTOR | [178] |
| Phase II | Everolimus + cisplatin + 5-fluorouracil Everolimus + cisplatin, 5-fluorouracil + leucovorin (advanced GC) | mTOR | [179] |
| Single-center, phase 2 | TAS-117 | AKT | [184] |
| Phase IB, multicenter | BYL719 | PI3K | [185] |
| Phase Ib/IIa, open-label, non-randomized | GSK2636771 + paclitaxel | PI3Kβ | [186] |
| First pilot metastatic | Cytotherapy + ramucirumab | PI3K and MAPK | [187] |
| ABT-806 | EGFR | ||
| standard cytotherapy + trastuzumab | HER 2 | ||
| VIKTORY | capivasertib | AKT, PI3K | [189] |
| vistusertib | TORC | ||
| selumetinib | MEK | ||
| capivasertib + paclitaxel | PI3K/AKT | ||
| The randomized, double-blind, placebo-controlled, multicenter, phase II trial | Ipatasertib + mFOLFOX6 | AKT | [190] |
| A randomized, multicenter, open-label phase IIa | Pertuzumab + trastuzumab | HER2 | [191] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morgos, D.-T.; Stefani, C.; Miricescu, D.; Greabu, M.; Stanciu, S.; Nica, S.; Stanescu-Spinu, I.-I.; Balan, D.G.; Balcangiu-Stroescu, A.-E.; Coculescu, E.-C.; et al. Targeting PI3K/AKT/mTOR and MAPK Signaling Pathways in Gastric Cancer. Int. J. Mol. Sci. 2024, 25, 1848. https://doi.org/10.3390/ijms25031848
Morgos D-T, Stefani C, Miricescu D, Greabu M, Stanciu S, Nica S, Stanescu-Spinu I-I, Balan DG, Balcangiu-Stroescu A-E, Coculescu E-C, et al. Targeting PI3K/AKT/mTOR and MAPK Signaling Pathways in Gastric Cancer. International Journal of Molecular Sciences. 2024; 25(3):1848. https://doi.org/10.3390/ijms25031848
Chicago/Turabian StyleMorgos, Diana-Theodora, Constantin Stefani, Daniela Miricescu, Maria Greabu, Silviu Stanciu, Silvia Nica, Iulia-Ioana Stanescu-Spinu, Daniela Gabriela Balan, Andra-Elena Balcangiu-Stroescu, Elena-Claudia Coculescu, and et al. 2024. "Targeting PI3K/AKT/mTOR and MAPK Signaling Pathways in Gastric Cancer" International Journal of Molecular Sciences 25, no. 3: 1848. https://doi.org/10.3390/ijms25031848