
Molecular Genetic Characteristics of the Hoxc13 Gene and Association Analysis of Wool Traits

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
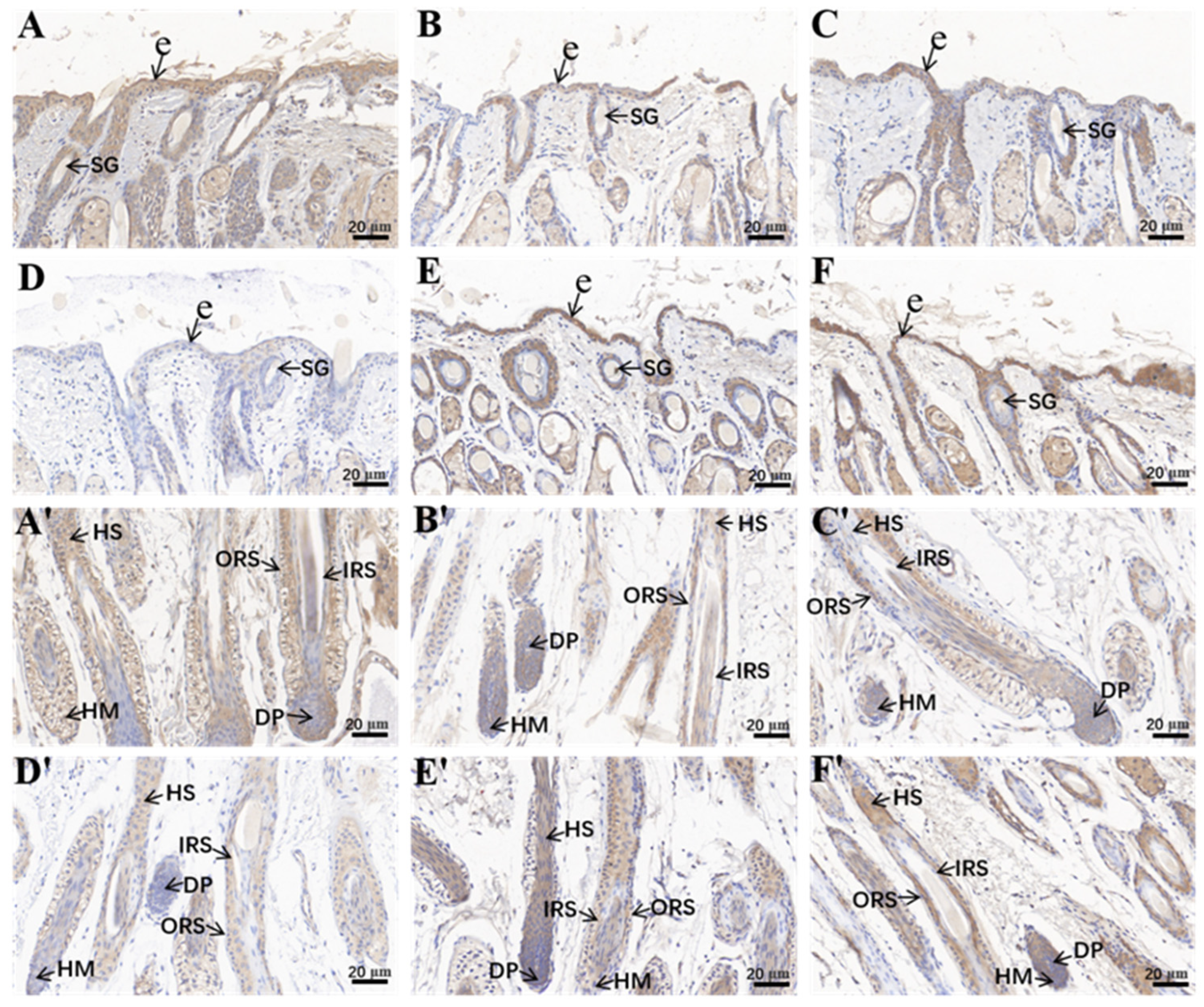
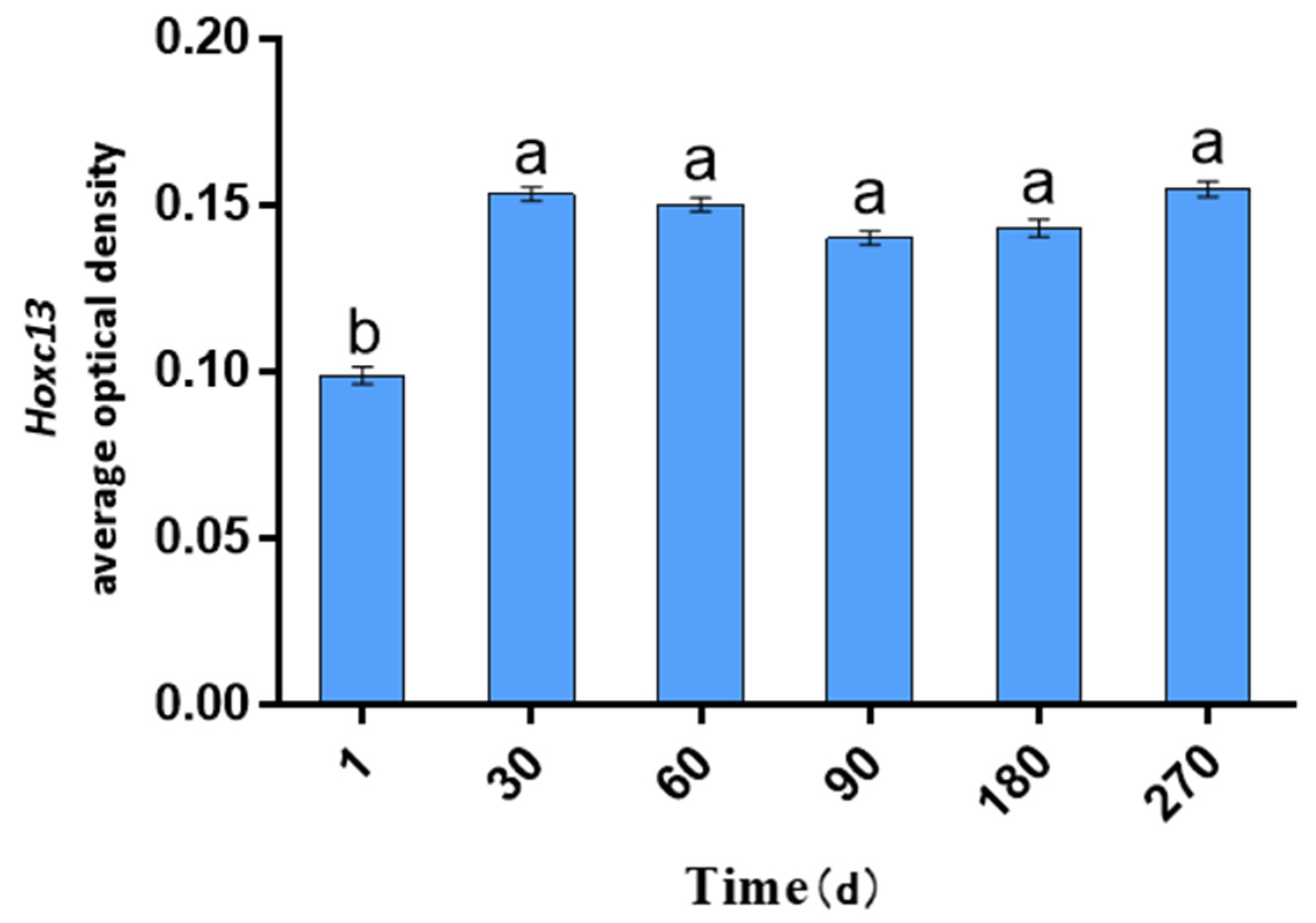
2.1. AOD Values of Hoxc13 in Different Stages of Skin Follicle Cycle Development
2.2. Distribution and Localization of Hoxc13 in Different Stages of Hair Follicle Cycle Development in the Skin
2.3. PCR Amplification Product Electrophoresis, Product Sequencing Results, and SNP Loci Detection Results
2.4. Genotype Frequencies, Gene Frequencies, and Population Genetic Diversity at Each Locus of the Hoxc13 Gene
2.5. Haplotype Analysis in the Hoxc13 Gene of Gansu Alpine Fine-Wool Sheep
2.6. Results of RT-qPCR for Different Genotypes of Two SNPs in the Hoxc13 Gene of Gansu Alpine Fine-Wool Sheep
3. Discussion
3.1. Protein Expression and Localization Study of Hoxc13 in Skin Tissues of Different Ages
3.2. Identification of Hoxc13 Polymorphisms and Association Analysis of Wool Traits
4. Materials and Methods
4.1. Animal Samples
4.2. Immunohistochemical Analysis
4.3. Immunofluorescence Analysis
4.4. PCR Amplification and Genotyping
4.5. RT-qPCR Analysis
4.6. Measurements and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Müller-Röver, S.; Handjiski, B.; van der Veen, C.; Eichmüller, S.; Foitzik, K.; McKay, I.A.; Stenn, K.S.; Paus, R. A comprehensive guide for the accurate classification of murine hair follicles in distinct hair cycle stages. J. Investig. Dermatol. 2001, 117, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Sennett, R.; Rendl, M.J.S.I.C.; Biology, D. Mesenchymal-epithelial interactions during hair follicle morphogenesis and cycling. Semin. Cell Dev. Biol. 2012, 23, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, K.; Toyoshima, K.E.; Tsuji, T. Functional hair follicle regeneration by the rearrangement of stem cells. Methods Mol. Biol. 2017, 1597, 117–134. [Google Scholar] [PubMed]
- Tsai, S.Y.; Sennett, R.; Rezza, A.; Clavel, C.; Grisanti, L.; Zemla, R.; Najam, S.; Rendl, M.J.D.B. Wnt/β-catenin signaling in dermal condensates is required for hair follicle formation. Dev. Biol. 2014, 385, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, A.; Krishnaswami, S.R.; Cowing-Zitron, C.; Hung, N.J.; Reilly-Rhoten, H.; Burns, J.; Yu, B.D. Negative regulation of shh levels by kras and fgfr2 during hair follicle development. Dev. Biol. 2013, 373, 373–382. [Google Scholar] [CrossRef]
- Mizutari, K.; Fujioka, M.; Hosoya, M.; Bramhall, N.; Okano, H.; Okano, H.; Edge, A.S. Notch inhibition induces cochlear hair cell regeneration and recovery of hearing after acoustic trauma. Neuron 2013, 77, 58–69. [Google Scholar] [CrossRef]
- Li, J.; Jiang, T.X.; Chuong, C.M. Many paths to alopecia via compromised regeneration of hair follicle stem cells. J. Investig. Dermatol. 2013, 133, 1450–1452. [Google Scholar] [CrossRef]
- Godwin, A.R.; Capecchi, M.R. Hoxc13 mutant mice lack external hair. Genes Dev. 1998, 12, 11–20. [Google Scholar] [CrossRef]
- Stelnicki, E.J.; Kömüves, L.G.; Kwong, A.O.; Holmes, D.; Klein, P.; Rozenfeld, S.; Lawrence, H.J.; Adzick, N.S.; Harrison, M.; Largman, C. Hox homeobox genes exhibit spatial and temporal changes in expression during human skin development. J. Investig. Dermatol. 1998, 110, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Kasiri, S.; Ansari, K.I.; Hussain, I.; Bhan, A.; Mandal, S.S. Antisense oligonucleotide mediated knockdown of hoxc13 affects cell growth and induces apoptosis in tumor cells and over expression of hoxc13 induces 3d-colony formation. RSC Adv. 2013, 3, 3260–3269. [Google Scholar] [CrossRef] [PubMed]
- Thummel, R.; Li, L.; Tanase, C.; Sarras, M.P.; Godwin, A.R. Differences in expression pattern and function between zebrafish hoxc13 orthologs: Recruitment of hoxc13b into an early embryonic role. Dev. Biol. 2004, 274, 318–333. [Google Scholar] [CrossRef] [PubMed]
- Tkatchenko, A.V.; Visconti, R.P.; Shang, L.; Papenbrock, T.; Pruett, N.D.; Ito, T.; Ogawa, M.; Awgulewitsch, A. Overexpression of hoxc13 in differentiating keratinocytes results in downregulation of a novel hair keratin gene cluster and alopecia. Development 2001, 128, 1547–1558. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Chen, Q.; Shi, L.; Lee, M.; Giehl, K.A.; Tang, Z.; Wang, H.; Jie, Z.; Yin, J.; Wu, L.; et al. Loss-of-function mutations in hoxc13 cause pure hair and nail ectodermal dysplasia. Am. J. Hum. Genet. 2012, 91, 906–911. [Google Scholar] [CrossRef]
- Han, K.; Liang, L.; Li, L.; Ouyang, Z.; Zhao, B.; Wang, Q.; Liu, Z.; Zhao, Y.; Ren, X.; Jiang, F.; et al. Generation of hoxc13 knockout pigs recapitulates human ectodermal dysplasia-9. Hum. Mol. Genet. 2017, 26, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Humbatova, A.; Maroofian, R.; Romano, M.T.; Tafazzoli, A.; Behnam, M.; Dilaver, N.; Nouri, N.; Salehi, M.; Wolf, S.; Frank, J.; et al. An insertion mutation in hoxc13 underlies pure hair and nail ectodermal dysplasia with lacrimal duct obstruction. Br. J. Dermatol. 2018, 178, e265–e267. [Google Scholar] [CrossRef]
- Sulayman, A.; Tian, K.; Huang, X.; Tian, Y.; Xu, X.; Fu, X.; Zhao, B.; Wu, W.; Wang, D.; Yasin, A.; et al. Genome-wide identification and characterization of long non-coding rnas expressed during sheep fetal and postnatal hair follicle development. Sci. Rep. 2019, 9, 8501. [Google Scholar] [CrossRef]
- Zhang, R.; Wu, H.; Lian, Z.J.G. Bioinformatics analysis of evolutionary characteristics and biochemical structure of fgf5 gene in sheep. Gene 2019, 702, 123–132. [Google Scholar] [CrossRef]
- Wang, J.; Yu, P.; Wang, H.; He, Y. Hoxc13 and hsp27 expression in skin and the periodic growth of secondary fiber follicles from longdong cashmere goats raised in different production systems. Anat. Rec. 2018, 301, 742–752. [Google Scholar] [CrossRef]
- He, Y.; Luo, Y.; Cheng, L.; Wang, J.; Liu, X.; Shao, B.; Cui, Y. Determination of secondary follicle characteristics, density, activity, and hoxc13 expression pattern of hexi cashmere goats breed. Anat. Rec. 2015, 298, 1796–1803. [Google Scholar] [CrossRef]
- He, Y.; Liu, X.; De, J.; Kang, S.; Munday, J.S. Altered hypoxia-induced and heat shock protein immunostaining in secondary hair follicles associated with changes in altitude and temperature in tibetan cashmere goats. Animals 2021, 11, 2798. [Google Scholar] [CrossRef]
- Jeong, K.H.; Joo, H.J.; Kim, J.E.; Park, Y.M.; Kang, H.J.C.; Dermatology, E. Effect of mycophenolic acid on proliferation of dermal papilla cells and induction of anagen hair follicles. Clin. Exp. Dermatol. 2015, 40, 894–902. [Google Scholar] [CrossRef]
- Morgan, B.A. The dermal papilla: An instructive niche for epithelial stem and progenitor cells in development and regeneration of the hair follicle. Cold Spring Harb. Perspect. Med. 2014, 4, a015180. [Google Scholar] [CrossRef]
- Farooq, M.; Kurban, M.; Fujimoto, A.; Fujikawa, H.; Abbas, O.; Nemer, G.; Saliba, J.; Sleiman, R.; Tofaili, M.; Kibbi, A.G.; et al. A homozygous frameshift mutation in the hoxc13 gene underlies pure hair and nail ectodermal dysplasia in a syrian family. Hum. Mutat. 2013, 34, 578–581. [Google Scholar]
- Lee, S.T.; Choi, K.W.; Yeo, H.T.; Kim, J.W.; Ki, C.S.; Cho, Y.D. Identification of an arg35x mutation in the pdcd10 gene in a patient with cerebral and multiple spinal cavernous malformations. J. Neurol. Sci. 2008, 267, 177–181. [Google Scholar] [CrossRef]
- Sharma, Y.; Miladi, M.; Dukare, S.; Boulay, K.; Caudron-Herger, M.; Groß, M.; Backofen, R.; Diederichs, S. A pan-cancer analysis of synonymous mutations. Nat. Commun. 2019, 10, 2569. [Google Scholar] [CrossRef]
- Sauna, Z.E.; Kimchi-Sarfaty, C. Understanding the contribution of synonymous mutations to human disease. Nat. Rev. Genet. 2011, 12, 683–691. [Google Scholar] [CrossRef]
- Shim, J.; Park, J.; Abudureyimu, G.; Kim, M.H.; Shim, J.S.; Jang, K.T.; Kwon, E.J.; Jang, H.S.; Yeo, E.; Lee, J.H.; et al. Comparative spatial transcriptomic and single-cell analyses of human nail units and hair follicles demonstrate transcriptional similarities between the onychodermis and follicular dermal papilla. J. Investig. Dermatol. 2022, 142, 3146–3157. [Google Scholar] [CrossRef]
- Qiu, W.; Lei, M.; Tang, H.; Yan, H.; Wen, X.; Zhang, W.; Tan, R.; Wang, D.; Wu, J. Hoxc13 is a crucial regulator of murine hair cycle. Cell Tissue Res. 2016, 364, 149–158. [Google Scholar] [CrossRef]
- Naeem, M.; John, P.; Ali, G.; Ahmad, W. Pure hair-nail ectodermal dysplasia maps to chromosome 12p11.1-q21.1 in a consanguineous pakistani family. Clin. Exp. Dermatol. 2007, 32, 502–505. [Google Scholar] [CrossRef]
- Perez, C.J.; Mecklenburg, L.; Fernandez, A.; Cantero, M.; de Souza, T.A.; Lin, K.; Dent, S.Y.R.; Montoliu, L.; Awgulewitsch, A.; Benavides, F. Naked (n) mutant mice carry a nonsense mutation in the homeobox of hoxc13. Exp. Dermatol. 2022, 31, 330–340. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, H.; Lei, C.; Pan, C.; Chen, H.; Lin, Q.; Lan, X. Effects of genetic variations within goat pitx2 gene on growth traits and mrna expression. Anim. Biotechnol. 2020, 31, 107–114. [Google Scholar] [CrossRef]
- Cai, H.; Lan, X.; Li, A.; Zhou, Y.; Sun, J.; Lei, C.; Zhang, C.; Chen, H. Snps of bovine hgf gene and their association with growth traits in nanyang cattle. Res. Vet. Sci. 2013, 95, 483–488. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, X.; Li, F.; La, Y.; Li, G.; Zhang, Y.; Li, X.; Zhao, Y.; Song, Q.; Wang, W. The association of polymorphisms in the ovine ppargc1b and zeb2 genes with body weight in hu sheep. Anim. Biotechnol. 2022, 33, 90–97. [Google Scholar] [CrossRef]
- Zhang, Z.; He, X.; Liu, Q.; Tang, J.; Di, R.; Chu, M. Tgif1 and sf1 polymorphisms are associated with litter size in small tail han sheep. Reprod. Domest. Anim. 2020, 55, 1145–1153. [Google Scholar] [CrossRef]
- Zhao, L.; Li, F.; Yuan, L.; Zhang, X.; Zhang, D.; Li, X.; Zhang, Y.; Zhao, Y.; Song, Q.; Wang, J.; et al. Expression of ovine ctnna3 and cap2 genes and their association with growth traits. Gene 2022, 807, 145949. [Google Scholar] [CrossRef]
- Wang, W.; La, Y.; Li, F.; Liu, S.; Pan, X.; Li, C.; Zhang, X. Molecular characterization and expression profiles of the ovine lhβ gene and its association with litter size in chinese indigenous small-tailed han sheep. Animals 2020, 10, 460. [Google Scholar] [CrossRef]
- Awgulewitsch, A.; Tkatchenko, A.V. Hoxc13 Transgenic Mice Exhibiting Hair Loss and Ichthyosis-Like Syndrome. AU20010022721, 25 June 2001. [Google Scholar]
- Khan, A.K.; Muhammad, N.; Aziz, A.; Genetics, S. A novel mutation in homeobox DNA binding domain of hoxc13 gene underlies pure hair and nail ectodermal dysplasia (ectd9) in a pakistani family. BMC Med. Genet. 2017, 18, 42. [Google Scholar] [CrossRef]
- Ali, R.H.; Habib, R.; Ud-Din, N.; Khan, M.N.; Ansar, M.; Ahmad, W. Novel mutations in the gene hoxc13 underlying pure hair and nail ectodermal dysplasia in consanguineous families. Br. J. Dermatol. 2013, 169, 478–480. [Google Scholar] [CrossRef]
- Li, X.; Orseth, M.L.; Smith, J.M.; Brehm, M.A.; Agim, N.G.; Glass, D.A., 2nd. A novel homozygous missense mutation in hoxc13 leads to autosomal recessive pure hair and nail ectodermal dysplasia. Pediatr. Dermatol. 2017, 34, 172–175. [Google Scholar] [CrossRef]
- Shanhe, W.; Zhixin, L.; Yuelang, Z.; Dan, Y.; Wei, G.; Xin, W. The inconsistent regulation of hoxc13 on different keratins and the regulation mechanism on hoxc13 in cashmere goat (Capra hircus). BMC Genom. 2018, 19, 630. [Google Scholar]
- Sun, H.; He, Z.; Xi, Q.; Zhao, F.; Hu, J.; Wang, J.; Liu, X.; Zhao, Z.; Li, M.; Luo, Y.; et al. Lef1 and dlx3 may facilitate the maturation of secondary hair follicles in the skin of gansu alpine merino. Genes 2022, 13, 1326. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days | Corneum | Inner Root Sheath | Outer Root Sheath | Hair Medulla | Dermal Papillae |
|---|---|---|---|---|---|
| 1 | +++ | ++++ | ++++ | + | ++++ |
| 30 | ++ | ++++ | ++++ | + | ++++ |
| 60 | ++ | ++++ | ++++ | + | ++++ |
| 90 | ++ | ++++ | ++++ | + | ++++ |
| 180 | ++ | ++++ | ++++ | + | ++++ |
| 270 | ++ | ++++ | ++++ | + | ++++ |
| Gene | SNP Site | Number | Genotype Frequency (Number) | Allele Frequency | ||
|---|---|---|---|---|---|---|
| Hoxc13 | c.405 T > G | 310 | TT | TG | T | G |
| 0.8613 (267) | 0.1387 (43) | 0.9306 | 0.0694 | |||
| c.673 A > G | 310 | AA | AG | A | G | |
| 0.8613 (267) | 0.1387 (43) | 0.9306 | 0.0694 | |||
| Gene | SNP Site | Ho | He | Ne | PIC | Hardy–Weinberg p-Value |
|---|---|---|---|---|---|---|
| Hoxc13 | c.405 T > G | 0.8708 | 0.1292 | 1.1484 | 0.1209 | p > 0.05 |
| c.673 A > G | 0.8708 | 0.1292 | 1.1484 | 0.1209 | p > 0.05 |
| Gene | SNP Site | Trait | Genotype (Number) | |
|---|---|---|---|---|
| Hoxc13 | c.405 T > G | TT (267) | TG (43) | |
| Mean Fiber Diameter (μm) | 22.182 ± 0.172 | 22.384 ± 0.408 | ||
| Mean Staple Length (mm) | 80.019 ± 1.297 b | 94.114 ± 3.606 a | ||
| Mean Fiber Curve (n/mm) | 105.359 ± 0.802 | 105.514 ± 2.490 | ||
| Comfort Factor (%) | 88.241 ± 0.780 | 85.890 ± 3.120 | ||
| Fiber Shift | 12.245 ± 0.257 | 13.533 ± 0.881 | ||
| Strength (cN/dT) | 14.332 ± 0.416 | 14.847 ± 1.329 | ||
| c.673 A > G | AA (267) | AG (43) | ||
| Mean Fiber Diameter (μm) | 22.274 ± 0.174 | 21.887 ± 0.372 | ||
| Mean Staple Length (mm) | 79.717 ± 1.256 b | 95.941 ± 3.986 a | ||
| Mean Fiber Curve (n/mm) | 105.359 ± 0.791 | 109.169 ± 3.478 | ||
| Comfort Factor (%) | 87.793 ± 0.790 | 90.375 ± 3.163 | ||
| Fiber Shift | 12.433 ± 0.252 | 11.518 ± 1.156 | ||
| Strength (cN/dT) | 14.353 ± 0.410 | 14.381 ± 1.610 | ||
| Haplotype | SNP1 | SNP2 | Frequency | Haplotype Combination | Frequency |
|---|---|---|---|---|---|
| H1 (TA) | T | A | 0.913 | H1H1 | 0.836 |
| H2 (GG) | G | G | 0.051 | H1H2 | 0.047 |
| H3 (GA) | G | A | 0.018 | ||
| H4 (TG) | T | G | 0.018 |
| Haplotype Combination | Mean Fiber Diameter (μm) | Mean Staple Length (mm) | Mean Fiber Curve (n/mm) | Comfort Factor (%) | Fiber Shift | Strength (cN/dT) |
|---|---|---|---|---|---|---|
| H1H1 | 22.161 ± 0.168 | 79.970 ± 1.272 b | 105.143 ± 0.799 | 88.403 ± 0.767 | 12.296 ± 0.263 | 14.293 ± 0.427 |
| H1H2 | 22.332 ± 0.617 | 100.794 ± 4.062 a | 112.740 ± 4.611 | 85.600 ± 4.611 | 12.808 ± 1.376 | 16.551 ± 2.157 |
| Gene | GenBank Accession No. | Primer Sequence (5′–3′) | Product Length (bp) | Annealing Temperature (°C) | Application |
|---|---|---|---|---|---|
| Hoxc13 | ENSOARG00020018195 | F: GGCGGTGGAAACACCAGGAG | 1007 | 60 | PCR |
| R: TCCATCTGCAGCCCAGCAAAG | |||||
| Hoxc13 | AY_266017.1 | F: GATAGTCAGGTGTACTGCTC | 132 | 60 | RT-qPCR |
| R: CTGCGTACTCCTTCTCTAGC | |||||
| β–actin | NM_001009784 | F: AGCCTTCCTTCCTGGGCATGGA | 113 | 60 | |
| R: GGACAGCACCGTGTTGGCGTAGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, H.; He, Z.; Zhao, F.; Hu, J.; Wang, J.; Liu, X.; Zhao, Z.; Li, M.; Luo, Y.; Li, S. Molecular Genetic Characteristics of the Hoxc13 Gene and Association Analysis of Wool Traits. Int. J. Mol. Sci. 2024, 25, 1594. https://doi.org/10.3390/ijms25031594
Sun H, He Z, Zhao F, Hu J, Wang J, Liu X, Zhao Z, Li M, Luo Y, Li S. Molecular Genetic Characteristics of the Hoxc13 Gene and Association Analysis of Wool Traits. International Journal of Molecular Sciences. 2024; 25(3):1594. https://doi.org/10.3390/ijms25031594
Chicago/Turabian StyleSun, Hongxian, Zhaohua He, Fangfang Zhao, Jiang Hu, Jiqing Wang, Xiu Liu, Zhidong Zhao, Mingna Li, Yuzhu Luo, and Shaobin Li. 2024. "Molecular Genetic Characteristics of the Hoxc13 Gene and Association Analysis of Wool Traits" International Journal of Molecular Sciences 25, no. 3: 1594. https://doi.org/10.3390/ijms25031594