
Opioid Analgesic as a Positive Allosteric Modulator of Acid-Sensing Ion Channels
 , , and
, , and 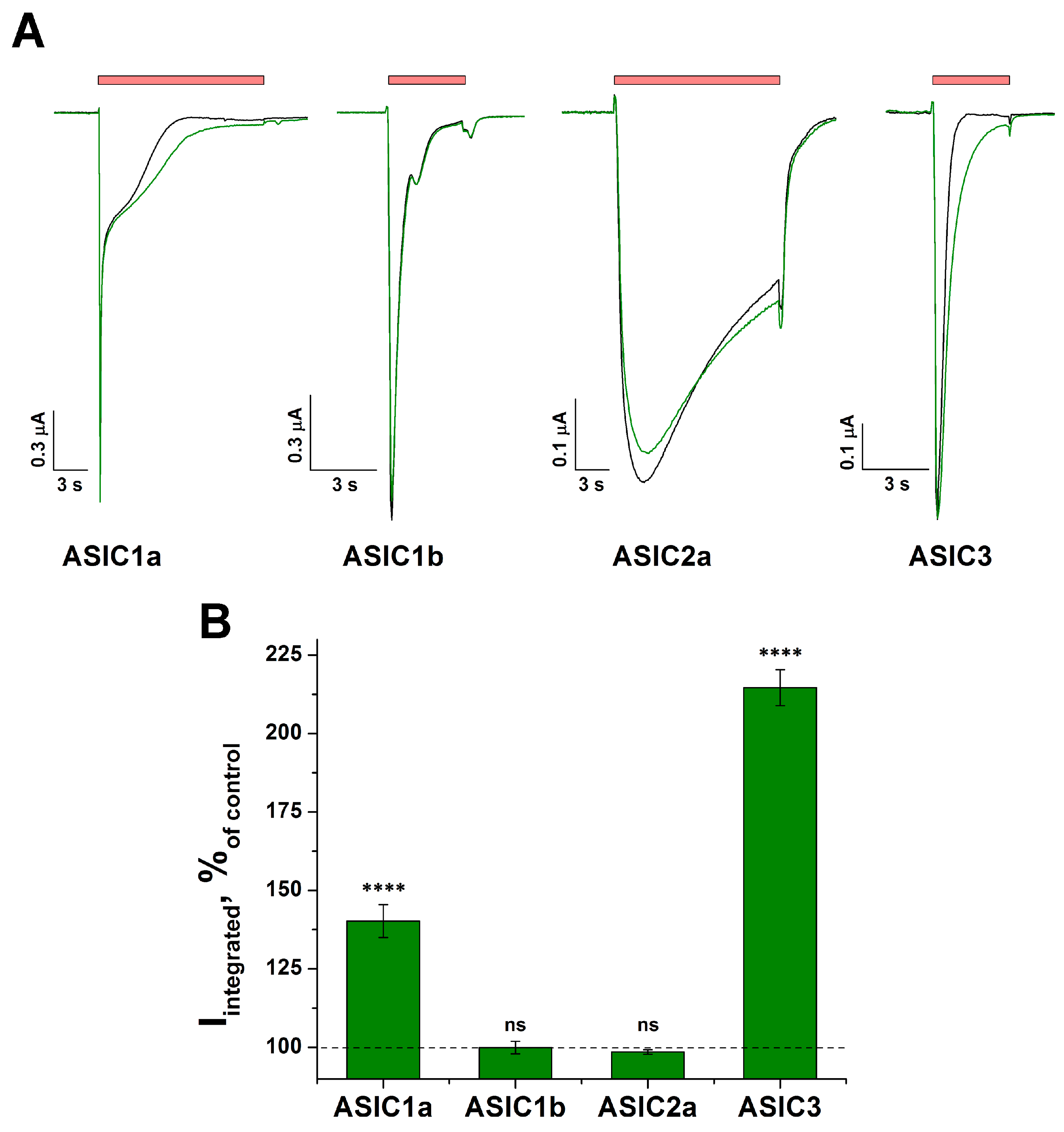{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
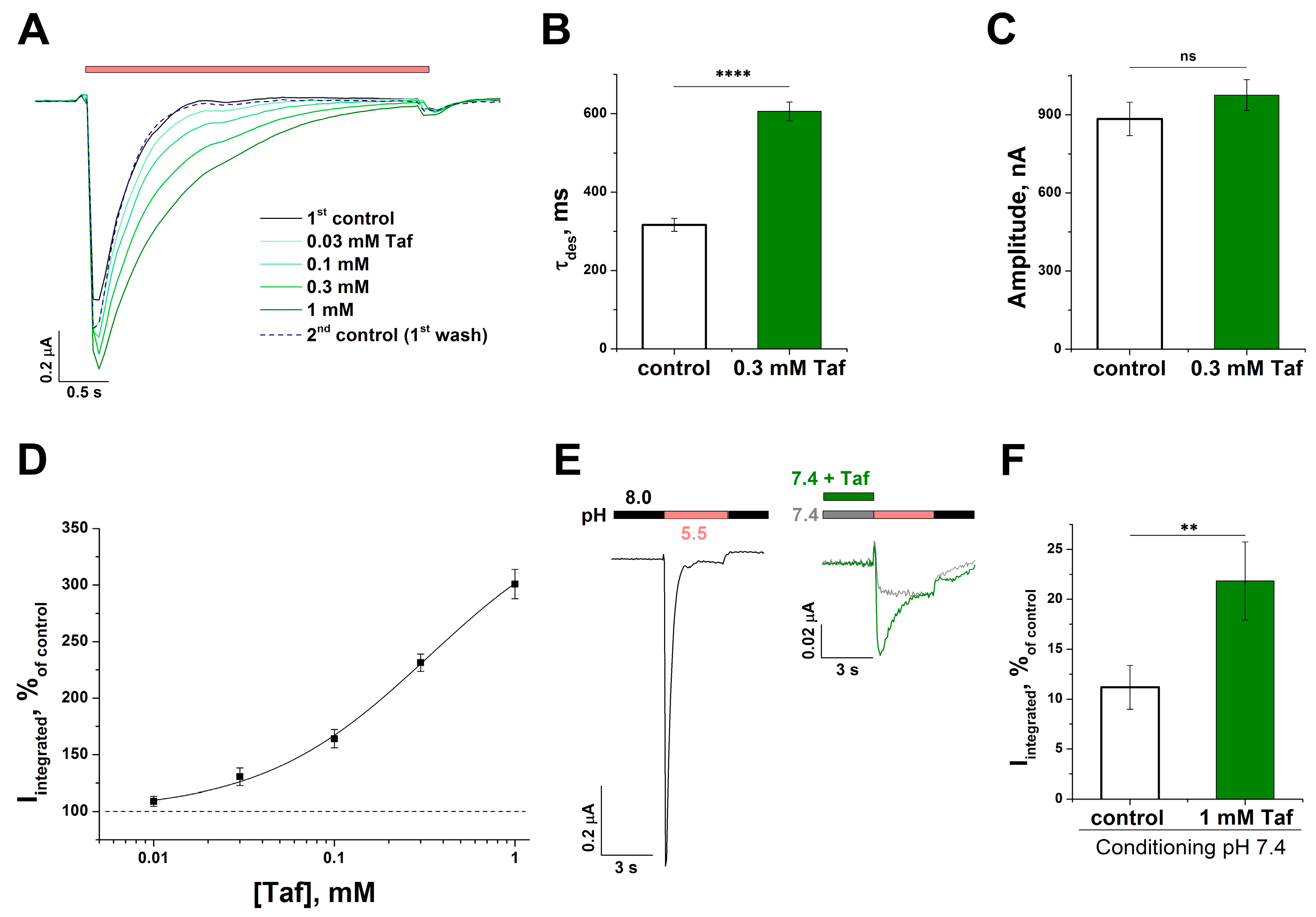
2.1. The Action of Tafalgin on Homomeric ASIC Channels
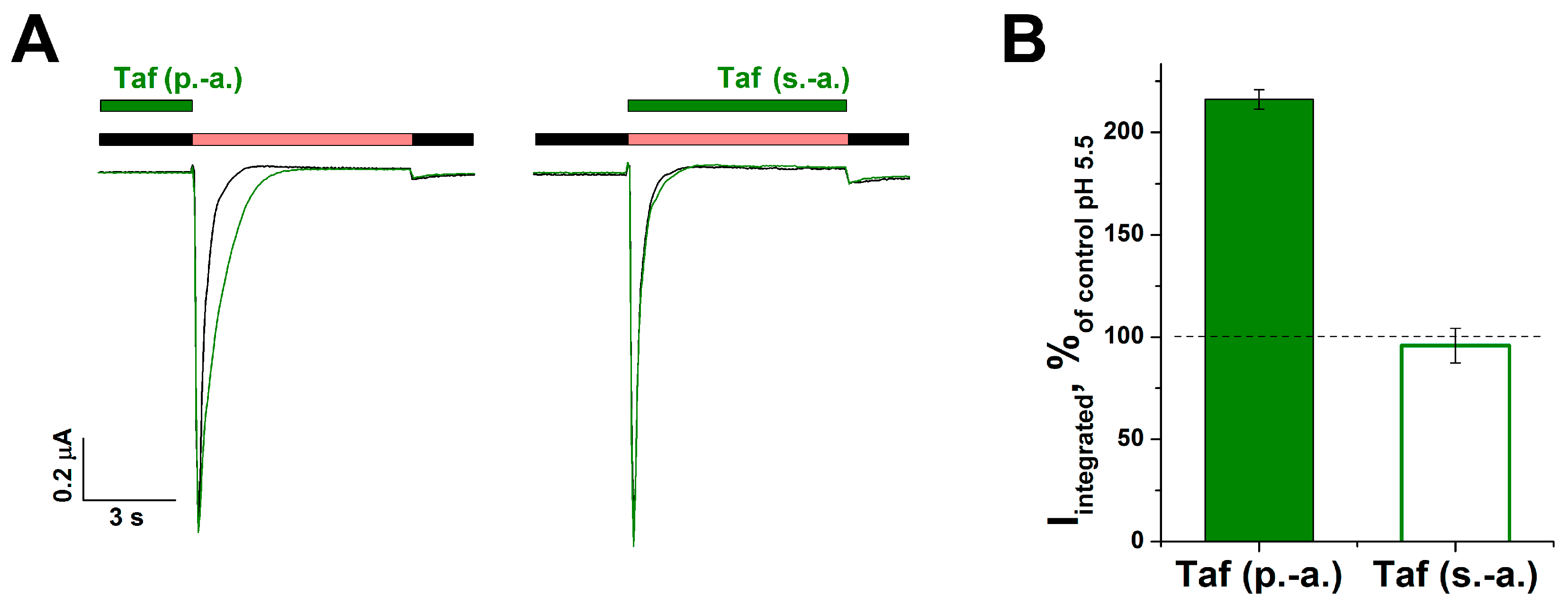
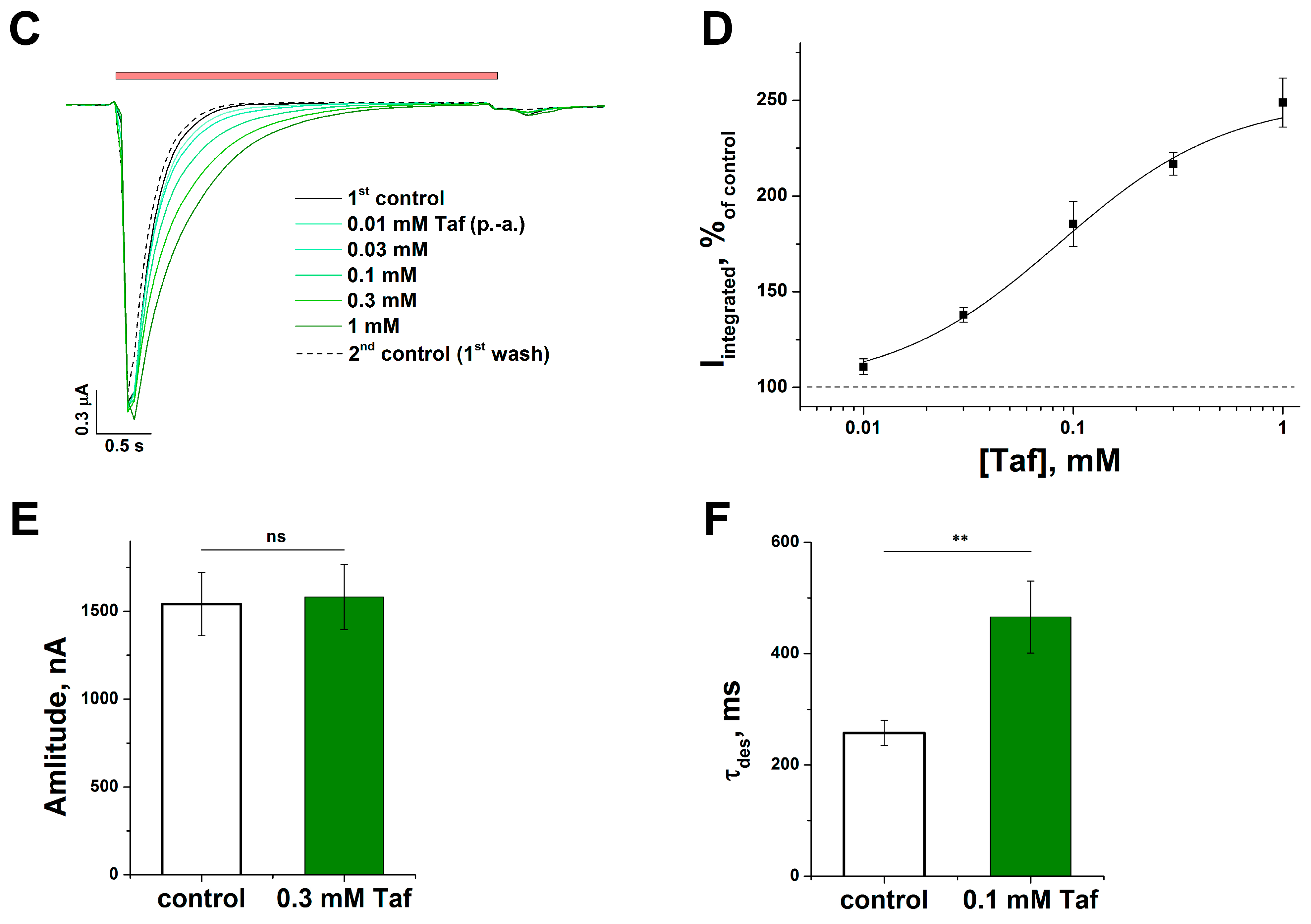
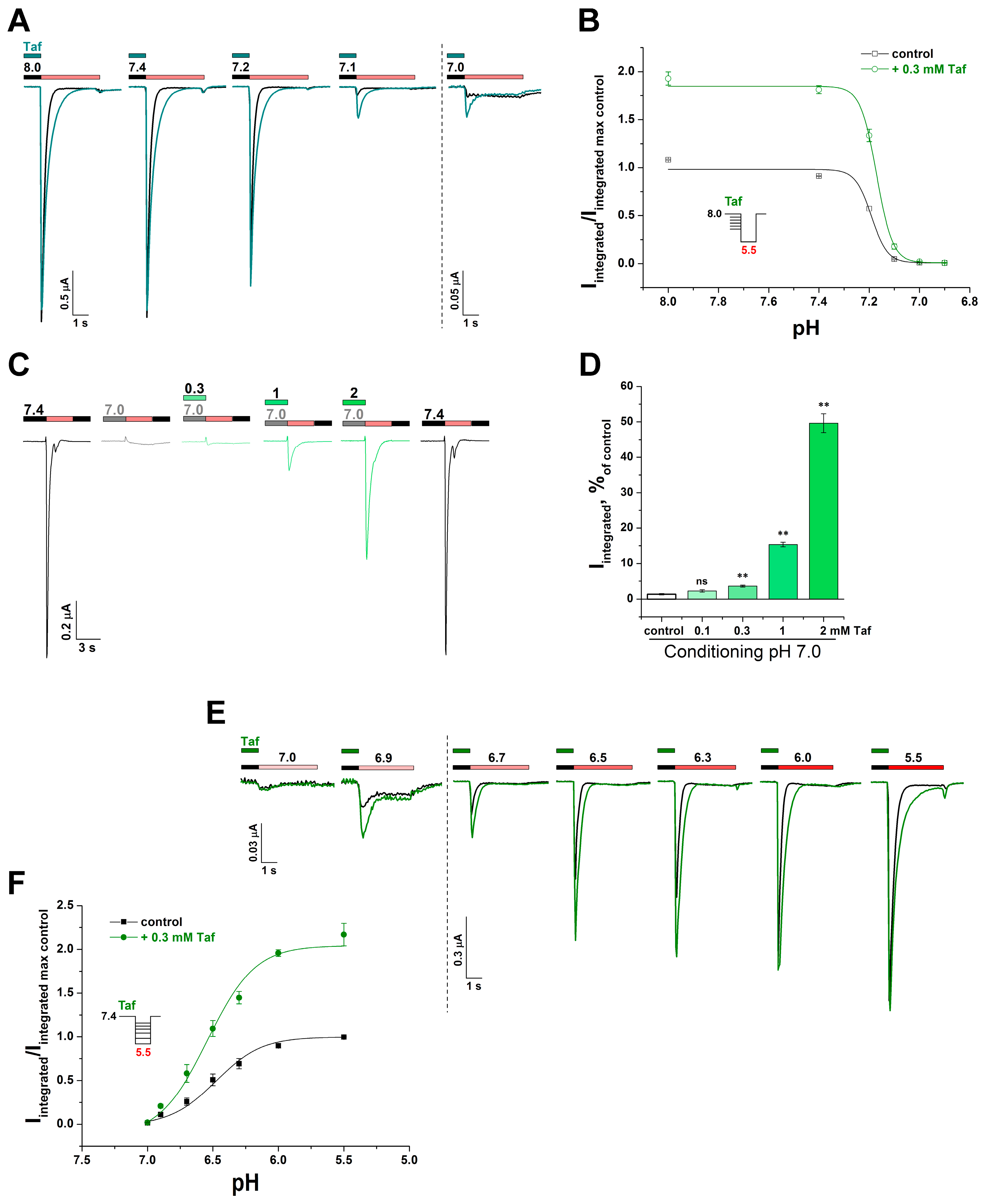
2.2. The Mechanism of the Tafalgin Action on the Rat ASIC3 Channel
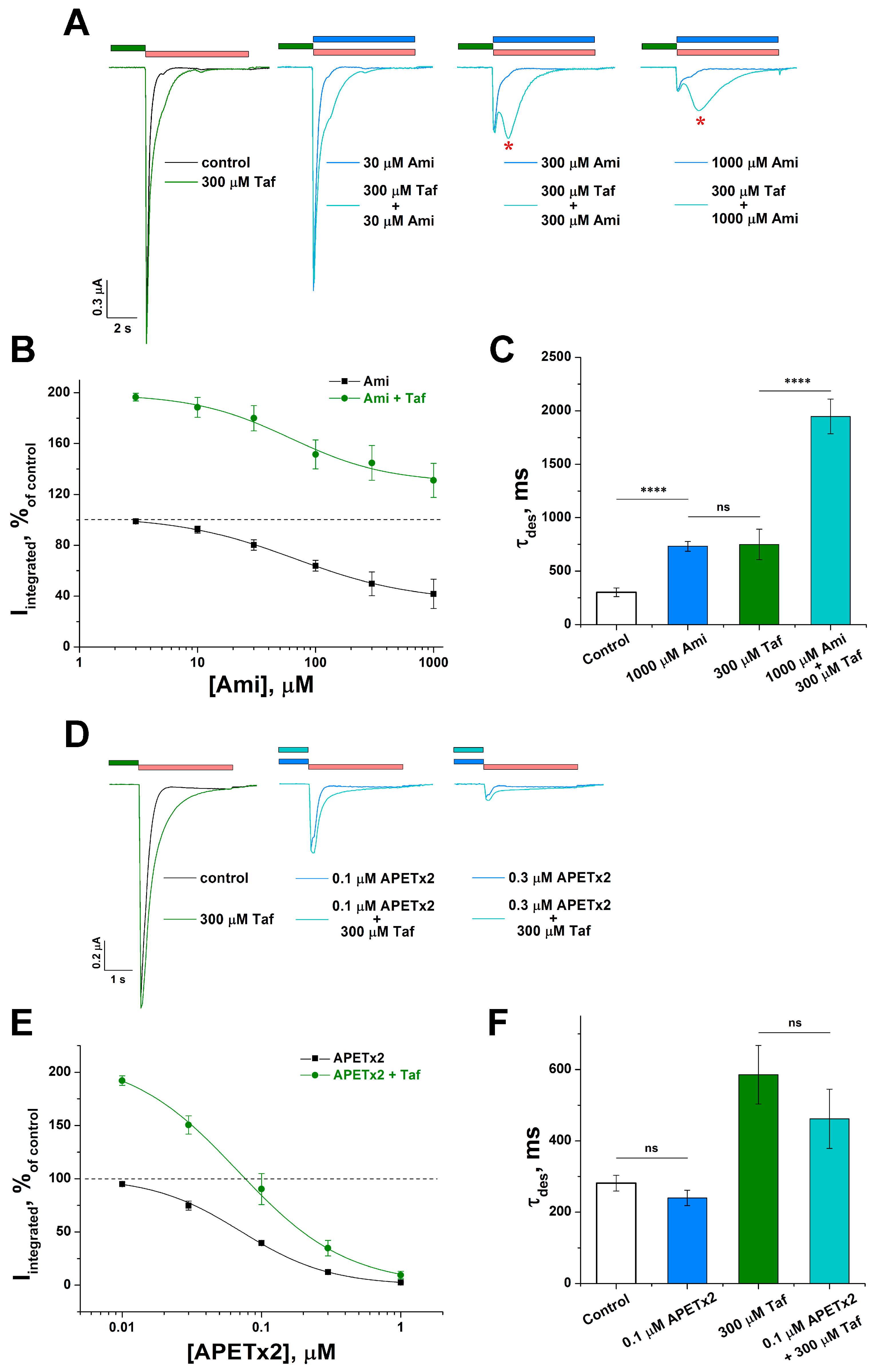
2.3. Cooperative Action of Tafalgin with ASIC3 Channel Antagonists
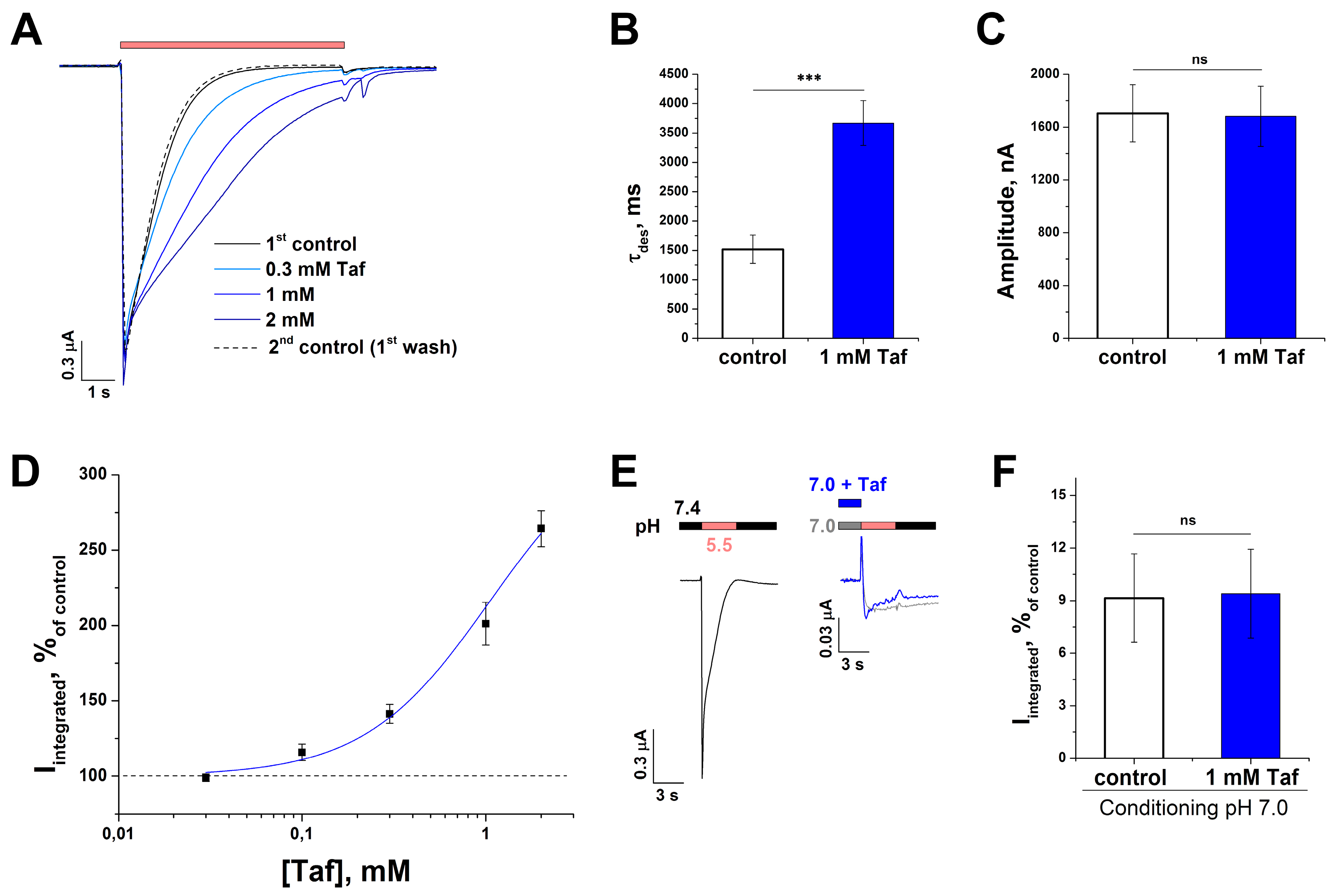
2.4. Effect of Tafalgin on Human ASIC3 Channels
2.5. Effect of Tafalgin on ASIC1a Channels
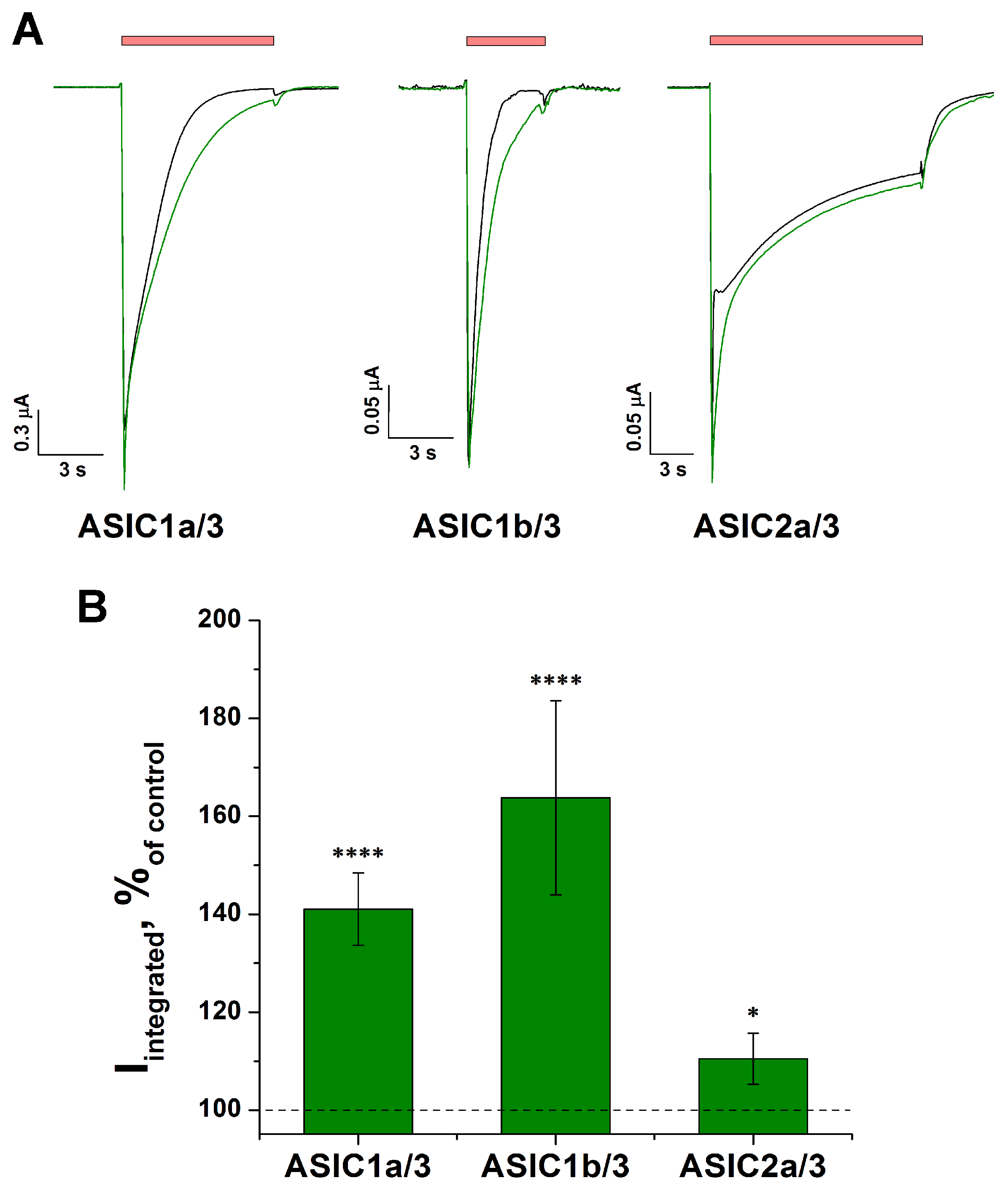
2.6. Effect of Tafalgin on Heteromeric Rat ASIC Channels
3. Discussion
4. Materials and Methods
4.1. Synthesis and Purification of Tafalgin
4.2. Chemical Reagents and ASIC Ligands
4.3. Xenopus laevis Oocyte Isolation and mRNA Injection
4.4. Two-Electrode Voltage-Clamp Recordings
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pergolizzi, J.; Böger, R.H.; Budd, K.; Dahan, A.; Erdine, S.; Hans, G.; Kress, H.G.; Langford, R.; Likar, R.; Raffa, R.B.; et al. Opioids and the management of chronic severe pain in the elderly: Consensus statement of an international expert panel with focus on the six clinically most often used world health organization step III opioids (Buprenorphine, Fentanyl, Hydromorphone, Met. Pain Pract. 2008, 8, 287–313. [Google Scholar] [CrossRef]
- Pasternak, G.W. Pharmacological mechanisms of opioid analgesics. Clin. Neuropharmacol. 1993, 16, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Harrison, C.; Lambert, D.G.; Smart, D. Stimulatory effects of opioids. Br. J. Anaesth. 1998, 81, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Al-Hasani, R.; Bruchas, M.R. Molecular mechanisms of opioid receptor-dependent signaling and behavior. Anesthesiology 2011, 115, 1363–1381. [Google Scholar] [CrossRef] [PubMed]
- Deval, E.; Lingueglia, E. Acid-Sensing Ion Channels and nociception in the peripheral and central nervous systems. Neuropharmacology 2015, 94, 49–57. [Google Scholar] [CrossRef]
- Boscardin, E.; Alijevic, O.; Hummler, E.; Frateschi, S.; Kellenberger, S. The function and regulation of acid-sensing ion channels (ASICs) and the epithelial Na + channel (ENaC): IUPHAR Review 19. Br. J. Pharmacol. 2016, 173, 2671–2701. [Google Scholar] [CrossRef]
- Kellenberger, S.; Schild, L. International Union of Basic and Clinical Pharmacology. XCI. Structure, Function, and Pharmacology of Acid-Sensing Ion Channels and the Epithelial Na+ Channel. Pharmacol. Rev. 2015, 67, 1–35. [Google Scholar] [CrossRef]
- Gründer, S.; Pusch, M. Biophysical properties of acid-sensing ion channels (ASICs). Neuropharmacology 2015, 94, 9–18. [Google Scholar] [CrossRef]
- Osmakov, D.I.; Khasanov, T.A.; Andreev, Y.A.; Lyukmanova, E.N.; Kozlov, S.A. Animal, Herb, and Microbial Toxins for Structural and Pharmacological Study of Acid-Sensing Ion Channels. Front. Pharmacol. 2020, 11, 991. [Google Scholar] [CrossRef]
- Du, J.; Reznikov, L.R.; Price, M.P.; Zha, X.-m.; Lu, Y.; Moninger, T.O.; Wemmie, J.A.; Welsh, M.J. Protons are a neurotransmitter that regulates synaptic plasticity in the lateral amygdala. Proc. Natl. Acad. Sci. USA 2014, 111, 8961–8966. [Google Scholar] [CrossRef]
- Ruan, N.; Tribble, J.; Peterson, A.M.; Jiang, Q.; Wang, J.Q.; Chu, X.-P. Acid-Sensing Ion Channels and Mechanosensation. Int. J. Mol. Sci. 2021, 22, 4810. [Google Scholar] [CrossRef] [PubMed]
- Dulai, J.S.; Smith, E.S.J.; Rahman, T. Acid-sensing ion channel 3: An analgesic target. Channels 2021, 15, 94–127. [Google Scholar] [CrossRef] [PubMed]
- Farrag, M.; Drobish, J.K.; Puhl, H.L.; Kim, J.S.; Herold, P.B.; Kaufman, M.P.; Ruiz-Velasco, V. Endomorphins potentiate acid-sensing ion channel currents and enhance the lactic acid-mediated increase in arterial blood pressure: Effects amplified in hindlimb ischaemia. J. Physiol. 2017, 595, 7167–7183. [Google Scholar] [CrossRef] [PubMed]
- Zaremba, M.; Ruiz-Velasco, V. Opioid-Mediated Modulation of Acid-Sensing Ion Channel Currents in Adult Rat Sensory Neurons. Mol. Pharmacol. 2019, 95, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Qiu, C.-Y.; Qiu, F.; Liu, T.-T.; Qu, Z.-W.; Liu, Y.-M.; Hu, W.-P. Morphine inhibits acid-sensing ion channel currents in rat dorsal root ganglion neurons. Brain Res. 2014, 1554, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Vick, J.S.; Askwith, C.C. ASICs and neuropeptides. Neuropharmacology 2015, 94, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Borg, C.B.; Braun, N.; Heusser, S.A.; Bay, Y.; Weis, D.; Galleano, I.; Lund, C.; Tian, W.; Haugaard-Kedström, L.M.; Bennett, E.P.; et al. Mechanism and site of action of big dynorphin on ASIC1a. Proc. Natl. Acad. Sci. USA 2020, 117, 7447–7454. [Google Scholar] [CrossRef] [PubMed]
- Leisle, L.; Margreiter, M.; Ortega-Ramírez, A.; Cleuvers, E.; Bachmann, M.; Rossetti, G.; Gründer, S. Dynorphin Neuropeptides Decrease Apparent Proton Affinity of ASIC1a by Occluding the Acidic Pocket. J. Med. Chem. 2021, 64, 13299–13311. [Google Scholar] [CrossRef]
- Kosorukov, V.S.; Abuzarova, G.R.; Zakharochkina, E.R.; Gamzeleva, O.Y.; Yatsenko, K.A. Tafalgin is a Russian innovative tetrapeptide pharmaceutical for subcutaneous injection: Review of the results of phase I and II clinical trials. Head Neck Tumors 2022, 12, 89–107. [Google Scholar] [CrossRef]
- Baron, A.; Lingueglia, E. Pharmacology of acid-sensing ion channels-Physiological and therapeutical perspectives. Neuropharmacology 2015, 94, 19–35. [Google Scholar] [CrossRef]
- Waldmann, R.; Bassilana, F.; de Weille, J.; Champigny, G.; Heurteaux, C.; Lazdunski, M. Molecular Cloning of a Non-inactivating Proton-gated Na + Channel Specific for Sensory Neurons. J. Biol. Chem. 1997, 272, 20975–20978. [Google Scholar] [CrossRef] [PubMed]
- Diochot, S.; Baron, A.; Rash, L.D.; Deval, E.; Escoubas, P.; Scarzello, S.; Salinas, M.; Lazdunski, M. A new sea anemone peptide, APETx2, inhibits ASIC3, a major acid-sensitive channel in sensory neurons. EMBO J. 2004, 23, 1516–1525. [Google Scholar] [CrossRef]
- Jensen, J.E.; Cristofori-Armstrong, B.; Anangi, R.; Rosengren, K.J.; Lau, C.H.Y.; Mobli, M.; Brust, A.; Alewood, P.F.; King, G.F.; Rash, L.D. Understanding the Molecular Basis of Toxin Promiscuity: The Analgesic Sea Anemone Peptide APETx2 Interacts with Acid-Sensing Ion Channel 3 and hERG Channels via Overlapping Pharmacophores. J. Med. Chem. 2014, 57, 9195–9203. [Google Scholar] [CrossRef] [PubMed]
- Osmakov, D.I.; Koshelev, S.G.; Andreev, Y.A.; Kozlov, S.A. Endogenous isoquinoline alkaloids agonists of acid-sensing ion channel type 3. Front. Mol. Neurosci. 2017, 10, 282. [Google Scholar] [CrossRef] [PubMed]
- Wemmie, J.A.; Chen, J.; Askwith, C.C.; Hruska-Hageman, A.M.; Price, M.P.; Nolan, B.C.; Yoder, P.G.; Lamani, E.; Hoshi, T.; Freeman, J.H.; et al. The Acid-Activated Ion Channel ASIC Contributes to Synaptic Plasticity, Learning, and Memory. Neuron 2002, 34, 463–477. [Google Scholar] [CrossRef]
- Wemmie, J.A.; Askwith, C.C.; Lamani, E.; Cassell, M.D.; Freeman, J.H.; Welsh, M.J. Acid-Sensing Ion Channel 1 Is Localized in Brain Regions with High Synaptic Density and Contributes to Fear Conditioning. J. Neurosci. 2003, 23, 5496–5502. [Google Scholar] [CrossRef] [PubMed]
- Kalinovskii, A.P.; Pushkarev, A.P.; Mikhailenko, A.D.; Kudryavtsev, D.S.; Belozerova, O.A.; Shmygarev, V.I.; Yatskin, O.N.; Korolkova, Y.V.; Kozlov, S.A.; Osmakov, D.I.; et al. Dual Modulator of ASIC Channels and GABAA Receptors from Thyme Alters Fear-Related Hippocampal Activity. Int. J. Mol. Sci. 2023, 24, 13148. [Google Scholar] [CrossRef] [PubMed]
- Duan, B.; Wang, Y.-Z.; Yang, T.; Chu, X.-P.; Yu, Y.; Huang, Y.; Cao, H.; Hansen, J.; Simon, R.P.; Zhu, M.X.; et al. Extracellular Spermine Exacerbates Ischemic Neuronal Injury through Sensitization of ASIC1a Channels to Extracellular Acidosis. J. Neurosci. 2011, 31, 2101–2112. [Google Scholar] [CrossRef]
- Vergo, S.; Craner, M.J.; Etzensperger, R.; Attfield, K.; Friese, M.A.; Newcombe, J.; Esiri, M.; Fugger, L. Acid-sensing ion channel 1 is involved in both axonal injury and demyelination in multiple sclerosis and its animal model. Brain 2011, 134, 571–584. [Google Scholar] [CrossRef]
- Heusser, S.A.; Pless, S.A. Acid-sensing ion channels as potential therapeutic targets. Trends Pharmacol. Sci. 2021, 42, 1035–1050. [Google Scholar] [CrossRef]
- Yu, Y.; Chen, Z.; Li, W.-G.; Cao, H.; Feng, E.-G.; Yu, F.; Liu, H.; Jiang, H.; Xu, T.-L. A Nonproton Ligand Sensor in the Acid-Sensing Ion Channel. Neuron 2010, 68, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Bohlen, C.J.; Chesler, A.T.; Sharif-Naeini, R.; Medzihradszky, K.F.; Zhou, S.; King, D.; Sánchez, E.E.; Burlingame, A.L.; Basbaum, A.I.; Julius, D. A heteromeric Texas coral snake toxin targets acid-sensing ion channels to produce pain. Nature 2011, 479, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Reimers, C.; Lee, C.-H.; Kalbacher, H.; Tian, Y.; Hung, C.-H.; Schmidt, A.; Prokop, L.; Kauferstein, S.; Mebs, D.; Chen, C.-C.; et al. Identification of a cono-RFamide from the venom of Conus textile that targets ASIC3 and enhances muscle pain. Proc. Natl. Acad. Sci. USA 2017, 114, E3507–E3515. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, T.W.; Askwith, C.C. Dynorphin Opioid Peptides Enhance Acid-Sensing Ion Channel 1a Activity and Acidosis-Induced Neuronal Death. J. Neurosci. 2009, 29, 14371–14380. [Google Scholar] [CrossRef] [PubMed]
- Deval, E.; Noël, J.; Lay, N.; Alloui, A.; Diochot, S.; Friend, V.; Jodar, M.; Lazdunski, M.; Lingueglia, E. ASIC3, a sensor of acidic and primary inflammatory pain. EMBO J. 2008, 27, 3047–3055. [Google Scholar] [CrossRef]
- Sluka, K.A.; Gregory, N.S. The dichotomized role for acid sensing ion channels in musculoskeletal pain and inflammation. Neuropharmacology 2015, 94, 58–63. [Google Scholar] [CrossRef]
- Diochot, S.; Baron, A.; Salinas, M.; Douguet, D.; Scarzello, S.; Dabert-Gay, A.S.; Debayle, D.; Friend, V.; Alloui, A.; Lazdunski, M.; et al. Black mamba venom peptides target acid-sensing ion channels to abolish pain. Nature 2012, 490, 552–555. [Google Scholar] [CrossRef]
- Chang, C.-T.; Fong, S.W.; Lee, C.-H.; Chuang, Y.-C.; Lin, S.-H.; Chen, C.-C. Involvement of Acid-Sensing Ion Channel 1b in the Development of Acid-Induced Chronic Muscle Pain. Front. Neurosci. 2019, 13, 1247. [Google Scholar] [CrossRef]
- Matasic, D.S.; Holland, N.; Gautam, M.; Gibbons, D.D.; Kusama, N.; Harding, A.M.S.; Shah, V.S.; Snyder, P.M.; Benson, C.J. Paradoxical Potentiation of Acid-Sensing Ion Channel 3 (ASIC3) by Amiloride via Multiple Mechanisms and Sites Within the Channel. Front. Physiol. 2021, 12, 1755. [Google Scholar] [CrossRef]
- Gei, L.; Yan, Y.; Xing, W.; Li, Q.; Chen, X.; Yan, F.; Wang, Y.; Cao, Y.; Jiang, W.; RiQi, E.; et al. Amiloride alleviates morphine tolerance by suppressing ASIC3-dependent neuroinflammation in the spinal cord. Eur. J. Pharmacol. 2023, 936, 176173. [Google Scholar] [CrossRef]
- Salinas, M.; Lazdunski, M.; Lingueglia, E. Structural Elements for the Generation of Sustained Currents by the Acid Pain Sensor ASIC3. J. Biol. Chem. 2009, 284, 31851–31859. [Google Scholar] [CrossRef] [PubMed]
- Logashina, Y.A.; Korolkova, Y.V.; Maleeva, E.E.; Osmakov, D.I.; Kozlov, S.A.; Andreev, Y.A. Refolding of disulfide containing peptides in fusion with thioredoxin. Mendeleev Commun. 2020, 30, 214–216. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osmakov, D.I.; Onoprienko, L.V.; Kalinovskii, A.P.; Koshelev, S.G.; Stepanenko, V.N.; Andreev, Y.A.; Kozlov, S.A. Opioid Analgesic as a Positive Allosteric Modulator of Acid-Sensing Ion Channels. Int. J. Mol. Sci. 2024, 25, 1413. https://doi.org/10.3390/ijms25031413
Osmakov DI, Onoprienko LV, Kalinovskii AP, Koshelev SG, Stepanenko VN, Andreev YA, Kozlov SA. Opioid Analgesic as a Positive Allosteric Modulator of Acid-Sensing Ion Channels. International Journal of Molecular Sciences. 2024; 25(3):1413. https://doi.org/10.3390/ijms25031413
Chicago/Turabian StyleOsmakov, Dmitry I., Lyudmila V. Onoprienko, Aleksandr P. Kalinovskii, Sergey G. Koshelev, Vasiliy N. Stepanenko, Yaroslav A. Andreev, and Sergey A. Kozlov. 2024. "Opioid Analgesic as a Positive Allosteric Modulator of Acid-Sensing Ion Channels" International Journal of Molecular Sciences 25, no. 3: 1413. https://doi.org/10.3390/ijms25031413