
Advance Research on the Pre-Harvest Sprouting Trait in Vegetable Crop Seeds
 and
and
Abstract
:1. Introduction
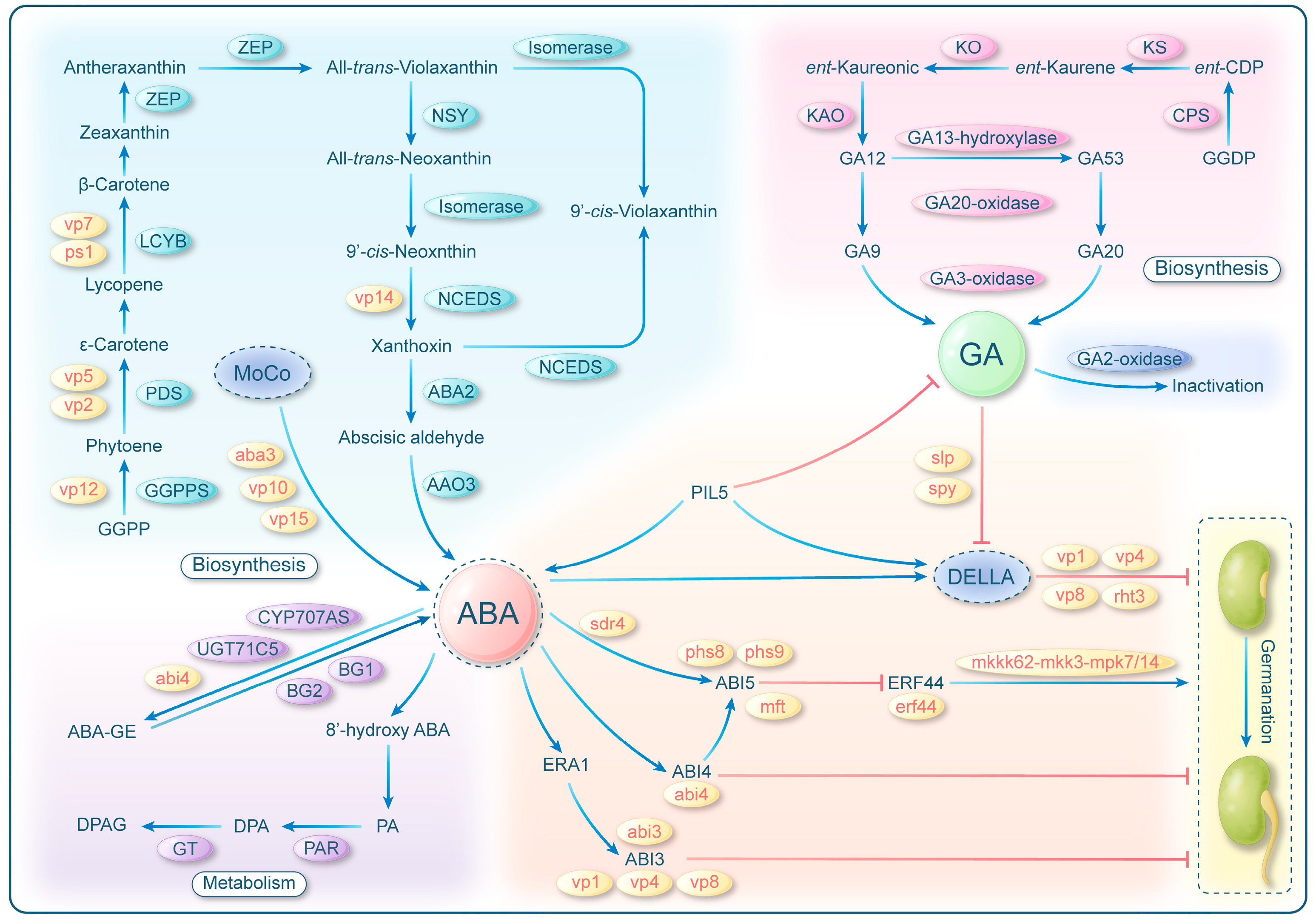
2. Pre-Harvest Sprouting Tightly Regulated by Plant Hormones
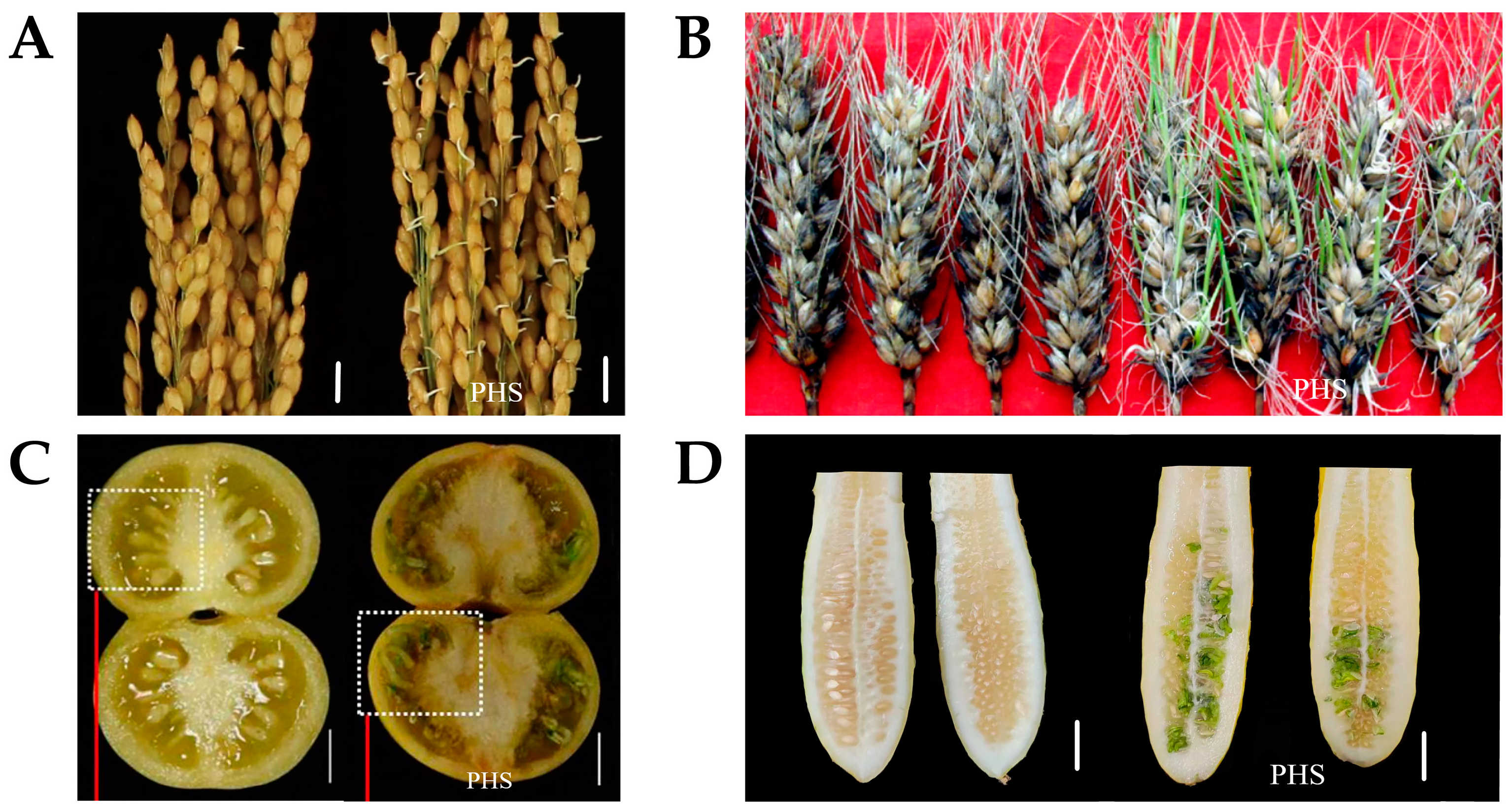
3. Pre-Harvest Sprouting as a Quantitative Trait Affected by Various Factors
3.1. Plant Hormone-Related Genes Play a Significant Role in the Pre-Harvest Sprouting Trait
3.2. Physiological Factors Influencing Pre-Harvest Sprouting in Vegetable Crops through Plant Hormones
3.3. Environmental Factors Affect Pre-Harvest Sprouting by Affecting the Contents of Plant Hormones
4. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pang, J.; Ma, D.; Huo, Z.; Li, S. Effects of Growth Time and Afterrepining on Maturation and Germination-in-fruit of Cucumber Seeds. In Proceedings of the 4th Youth Symposium of Chinese Horticultural Society, Harbin, China, August 2000; pp. 407–410. [Google Scholar]
- Zhang, C.; Zhou, L.; Lu, Y.; Yang, Y.; Feng, L.; Hao, W.; Li, Q.; Fan, X.; Zhao, D.; Liu, Q. Changes in the physicochemical properties and starch structures of rice grains upon pre-harvest sprouting. Carbohydr. Polym. 2020, 234, 115893. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Feng, F.; Xu, X.; Chen, W.; Zhou, H.; Ye, C. The Effect of Vivipary on Fruit Quality of Jackfruit. Southeast Hortic. 2021, 9, 1–4. [Google Scholar]
- Xu, F.; Tang, J.; Wang, S.; Cheng, X.; Wang, H.; Ou, S.; Gao, S.; Li, B.; Qian, Y.; Gao, C.; et al. Antagonistic control of seed dormancy in rice by two bHLH transcription factors. Nat. Genet. 2022, 12, 1972–1982. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Qu, B.; Li, H.; Zuo, D.; Zhao, Z.; Liao, Y. A maize viviparous 1 gene increases seed dormancy and preharvest sprouting tolerance in transgenic wheat. J. Cereal Sci. 2012, 2, 166–173. [Google Scholar] [CrossRef]
- Yao, M.; Chen, W.; Kong, J.; Zhang, X.; Shi, N.; Zhong, S.; Ma, P.; Gallusci, P.; Jackson, S.; Liu, Y.; et al. METHYLTRANSFERASE1 and Ripening Modulate Vivipary during Tomato Fruit Development. Plant Physiol. 2020, 183, 1883–1897. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Li, H.; Ma, D. Progress on Preharvest Sproution of Vegetable Crops. Tianjin Agric. Sci. 2002, 03, 42–46. [Google Scholar]
- Tian, X. Evaluation and Transcriptome Analysis of Seed Germination in Melon of Cucumber Germplasm Resources. Ph.D. Thesis, Northeast Agricultural University, Harbin, China, 2021. [Google Scholar]
- Guan, W.; Cao, M.; Li, S.; Wang, H.; Yang, R.; Deng, Q. Preliminary study on inhibition of preharvest cucumber seed germination by exogenous ABA. Bull. Agric. Sci. Technol. 2016, 05, 122–125. [Google Scholar]
- Mo, Y.; Wang, L.; Zhou, Y.; Sun, Q.; He, W.; Chen, H.; Xiao, L.; Wang, R. Physiological characteristics and related gene expression of vivipary in cucumber seeds. J. South. Agric. 2020, 51, 2090–2096. [Google Scholar]
- Li, C. Study on the Vivipary Chracteristics of Summer Squash Seed. Ph.D. Thesis, Shandong Agricultural University, Taian, China, 2017. [Google Scholar]
- Feng, F. Genetic Studies and QTL Mapping of Seed Vivipary in Brassica napus L. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2009. [Google Scholar]
- Groot, S.C.M. Karssen Dormancy and Germination of Abscisic Acid-Deficient Tomato Seeds—Studies with the Sitiens Mutant. Plant Physiol. 1992, 99, 952–958. [Google Scholar] [CrossRef]
- Wang, X. Physiological Mechanism of Vivipary in One Rin Mutant Tomato Accession and Role of Rin in Vivipary. Ph.D. Thesis, Northeast Agricultural University, Harbin, China, 2016. [Google Scholar]
- Sun, G.; Zhang, X.; Xiao, S. Maternal Effect on Sensitivity of Embryo to ABA and Grain Dormancy in Wheat. J. Triticeae Crops 2005, 25, 37–41. [Google Scholar]
- Zhang, L. Study on the Seed Vivipary Eharacteris tics of Eomm on Beall (Pllaseolu svulgaris L.). Ph.D. Thesis, Zhejiang University, Hangzhou, China, 2007. [Google Scholar]
- Jin, Y.; Zhu, H.; Zhang, M.; Hui, M. Preliminary Studies on Occurrence Time, Position and nheritance Genetics of Seed Vivipary in Chinese Cabbage (Brassica rapa). North Hortic. 2016, 09, 24–27. [Google Scholar]
- He, X. Mapping and Mining of Major Genes for Seed Vivipary Trait in Chinese Cabbage. Ph.D. Thesis, Northwest A&F University, Yangling, China, 2021. [Google Scholar]
- Cao, M.; Wang, H.; Deng, Q.; Yang, R.; Li, S. Quantitative Genetic Analysis of Pre-harvest Sprouting in Cucumber. Chian Veg. 2018, 01, 34–38. [Google Scholar]
- Cao, M.; Li, S.; Deng, Q.; Wang, H.; Yang, R. Identification of a major-effect QTL associated with pre-harvest sprouting in cucumber (Cucumis sativus L.) using the QTL-seq method. BMC Genom. 2021, 22, 2491. [Google Scholar] [CrossRef] [PubMed]
- Penfield, S. Seed dormancy and germination. Curr. Biol. 2017, 17, 874–878. [Google Scholar] [CrossRef] [PubMed]
- Hable, W.E.; Oishi, K.K.; Schumaker, K.S. Viviparous-5 encodes phytoene desaturase, an enzyme essential for abscisic acid (ABA) accumulation and seed development in maize. Mol. Gen. Genet. 1998, 257, 167–176. [Google Scholar] [CrossRef]
- Singh, M.; Lewis, P.E.; Hardeman, K.; Bai, L.; Rose, J.K.C.; Mazourek, M.; Chomet, P.; Brutnell, T.P. Activator mutagenesis of the pink scutellum1/viviparous7 locus of maize. Plant Cell 2003, 15, 874–884. [Google Scholar] [CrossRef]
- Maluf, M.P.; Saab, I.N.; Wurtzel, E.T.; Sachs, M.M. The viviparous12 maize mutant is deficient in abscisic acid, carotenoids, and chlorophyll synthesis. J. Experin. Botan. 1997, 48, 1259–1268. [Google Scholar] [CrossRef]
- Bittner, F.; Oreb, M.; Mendel, R.R. ABA3 is a molybdenum cofactor sulfurase required for activation of aldehyde oxidase and xanthine dehydrogenase in Arabidopsis thaliana. J. Biol. Chem. 2001, 276, 40381–40384. [Google Scholar] [CrossRef]
- Xiong, L.M.; Ishitani, M.; Lee, H.; Zhu, J.K. The Arabidopsis LOS5/ABA3 locus encodes a molybdenum cofactor sulfurase and modulates cold stress-responsive and osmotic stress-responsive gene expression. Plant Cell 2001, 13, 2063–2083. [Google Scholar]
- Leydecker, M.T.; Moureanx, T.; Kraepiel, Y.; Schnorr, K.; Caboche, M. Molybdenum Cofactor Mutants, Specifically Impaired in Xanthine Dehydrogenase-Activity and Absisic-Acid Biosynthesis, Simultaneously Overexpress Nitrate Reductase. Plant Physiol. 1995, 107, 1427–1431. [Google Scholar] [CrossRef]
- Sagi, M.; Scazzocchio, C.; Fluhr, R. The absence of molybdenum cofactor sulfuration is the primary cause of the flacca phenotype in tomato plants. Plant J. 2002, 31, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.C.; Schwartz, S.H.; Zeevaart, J.A.D.; McCarty, D.R. Genetic control of abscisic acid biosynthesis in maize. Proc. Natl. Acad. Sci. USA 1997, 94, 12235–12240. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Latshaw, S.; Sato, Y.; Settles, A.M.; Koch, K.E.; Hannah, L.C.; Kojima, M.; Sakakibara, H.; McCarty, D.R. The maize Viviparous8 locus, encoding a putative ALTERED MERISTEM PROGRAM1-like peptidase, regulates abscisic acid accumulation and coordinates embryo and endosperm development. Plant Physiol. 2008, 146, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Mccarty, D.R.; Hattori, T.; Carson, C.B.; Vasil, V.; Lazar, M.; Vasil, I.K. The VIVIPAROUS-1 Developmental Gene of Mazie Encodes a Novel Transcriptional Activator. Cell 1991, 66, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Hoecker, U.; Vasil, I.K.; Mccarty, D.R. Integrated control of seed maturation and germinationprograms by activator and repressor functions of Viviparous-1 of maize. Genes Dev. 1995, 20, 2459. [Google Scholar] [CrossRef] [PubMed]
- Mccarty, D.R.; Carson, C.B.; Stinard, P.S.; Robertson, D.S. Molecular Analysis of viviparous-1: An Abscisic Acid-Insensitive Mutant of Maize. Plant Cell 1989, 5, 523–532. [Google Scholar]
- Wang, R.; Chen, Y.; Sun, M.; Zhang, X.; Du, Y.; Zheng, J. Genetic analysis and causal gene identification of maize viviparous mutant vp-like8. Acta Agron. Sin. 2019, 45, 656–661. [Google Scholar] [CrossRef]
- Chu, L. Effect and Mechanism of AP2/ERF Teanscription Factor OsERF44 on Panicle Sprouting in Rice. Ph.D. Thesis, Yangzhou University, Yangzhou, China, 2021. [Google Scholar]
- Bentsink, L.; Jowett, J.; Hanhart, C.J.; Koornneef, M. Cloning of DOG1, a quantitative trait locus controlling seed dormancy in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 17042–17047. [Google Scholar] [CrossRef]
- Sugimoto, K.; Takeuchi, Y.; Ebana, K.; Miyao, A.; Hirochika, H.; Hara, N.; Ishiyama, K.; Kobayashi, M.; Ban, Y.; Hattori, T.; et al. Molecular cloning of Sdr4, a regulator involved in seed dormancy and domestication of rice. Proc. Natl. Acad. Sci. USA 2010, 107, 5792–5797. [Google Scholar] [CrossRef]
- Du, L.; Xu, F.; Fang, J.; Gao, S.; Tang, J.; Fang, S.; Wang, H.; Tong, H.; Zhang, F.; Chu, J.; et al. Endosperm sugar accumulation caused by mutation of PHS8/ISA1 leads to pre-harvest sprouting in rice. Plant J. 2018, 95, 545–556. [Google Scholar] [CrossRef]
- Xu, F.; Tang, J.; Gao, S.; Chang, X.; Du, L.; Chu, C. Control of rice pre-harvest sprouting by glutaredoxin-mediated abscisic acid signaling. Plant J. 2019, 100, 1036–1051. [Google Scholar] [CrossRef] [PubMed]
- Xi, W.; Liu, C.; Hou, X.; Yu, H. Motherof FT and TFL1 Regulates Seed Germination through a Negative Feedback Loop Modulating ABA Signaling in Arabidopsis. Plant Cell 2010, 22, 1733–1748. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, B.; Zang, Q. Progress in Seed Vivipary Mechanism. Chin. J. Cell Biol. 2007, 05, 701–705. [Google Scholar]
- Dai, C.; Xue, H. Rice early flowering1, a CKI, phosphorylates DELLA protein SLR1 to negatively regulate gibberellin signalling. EMBO J. 2010, 29, 1916–1927. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Zhang, H.; Wang, S.; Chen, M.; Wu, Y.; Tang, S.; Liu, C.; Feng, Y.; Cao, X.; Xie, Q. ABI4 Regulates Primary Seed Dormancy by Regulating the Biogenesis of Abscisic Acid and Gibberellins in Arabidopsis. PLoS Genet. 2013, 9, e1003577. [Google Scholar] [CrossRef] [PubMed]
- Dill, A.; Thomas, S.G.; Hu, J.H.; Steber, C.M.; Sun, T.P. The Arabidopsis F-box protein SLEEPY1 targets gibberellin signaling repressors for gibberellin-induced degradation. Plant Cell 2004, 16, 1392–1405. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, S.E.; Binkowski, K.A.; Olszewski, N.E. SPINDLY, a tetratricopeptide repeat protein involved in gibberellin signal transduction Arabidopsis. Proc. Natl. Acad. Sci. USA 1996, 93, 9292–9296. [Google Scholar] [CrossRef] [PubMed]
- Lang, J.; Fu, Y.; Zhou, Y.; Cheng, M.; Deng, M.; Li, M.; Zhu, T.; Yang, J.; Guo, X.; Gui, L.; et al. Myb10-D confers PHS-3D resistance to pre-harvest sprouting by regulating NCED in ABA biosynthesis pathway of wheat. New Phytol. 2021, 230, 1940–1952. [Google Scholar] [CrossRef]
- Bi, H.H.; Sun, Y.W.; Xiao, Y.G.; Xia, L.Q. Characterization of DFR allelic variations and their associations with pre-harvest sprouting resistance in a set of red-grained Chinese wheat germplasm. Euphytica 2014, 195, 197–207. [Google Scholar] [CrossRef]
- Masaharu, S.; Mark, S.A.; Chi-Wah, T. The maize viviparous15 locus encodes the molybdopterin synthase small submit. Pant J. 2010, 2, 264–274. [Google Scholar]
- Zhou, G.; Wu, S.; Chen, D.; Wu, X.; Cai, Q. Polyphenols and phytohormones profiling of pre-harvest sprouting resistant and susceptible wheat genotypes. SN Appl. Sci. 2023, 5, 249. [Google Scholar] [CrossRef]
- Sohn, S.-I.; Pandian, S.; Kumar, T.S.; Zoclanclounon, Y.A.B.; Muthuramalingam, P.; Shilpha, J.; Satish, L.; Ramesh, M. Seed Dormancy and Pre-Harvest Sprouting in Rice—An Updated Overview. Int. J. Mol. Sci. 2021, 22, 11804. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.X.; Chen, W. Study on the physiological biochemistry of pre-harvest sprouting and scanning electron microscopy of glume in rice. Chin. Agric. Sci. Bull. 2008, 1, 75–80. [Google Scholar]
- Xu, J. Analysis the Cuticular Wax and Influences of Tissue Culture Condition on Cuticular Wax and Water Saving of Tobacco Leaf. Ph.D. Thesis, Northwest A&F University, Yangling, China, 2014. [Google Scholar]
- Flintham, J.; Adlam, R.; Bassoi, M.; Holdsworth, M.; Gale, M. Mapping genes for resistance to sprouting damage in wheat. Euphytica 2002, 126, 39–45. [Google Scholar] [CrossRef]
- Shen, Z.; Yu, S.; Wu, S. Study on Pre-Harvest Sprouting Resistance in Wheat Cultivars. Sci. Agric. Sin. 1991, 05, 44–50. [Google Scholar]
- Zeevaart, J.A.D.; Creelman, R.A. Metabolism and Physiology of Abscisic-Acid. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1988, 39, 439–473. [Google Scholar] [CrossRef]
- Welbaum, G.E.; Bradford, K.J. Water Relations of Seed Development and Germination in Muskmelon (Cucumis melo L.) V. Water Relations of Imbibition and Germination. Plant Physiol. 1990, 92, 1046–1052. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Chen, X.; Han, Y. Progress on Preharvest Sprouting Resistance in White Wheat. J. Triticeae Crops 2000, 02, 84–87. [Google Scholar]
- Liu, X.; Yang, Q.; Zhang, H.Q.; He, J.W.; Yan, Y.T.; Ju, H. Influencing factors of the vivipary occurrence of rapeseed. J. Hunan Agric. Univ. Nat. Sci. 2022, 48, 8–12. [Google Scholar]
- Liu, Z.; Zou, Y.; Dong, X.; Wei, J.; Xu, C.; Mi, W.; Xu, M.; Fang, X.; Cao, X.; Zheng, G.; et al. Germinating seed can sense low temperature for the floral transition and vernalization of winter rapeseed (Brassica rapa). Plant Sci. 2021, 307, 110900. [Google Scholar] [CrossRef]
- Nee, G.; Xiang, Y.; Soppe, W.J.J. The release of dormancy, a wake-up call for seeds to germinate. Curr. Opin. Plant Biol. 2017, 35, 8–14. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, Y.; Hu, G.; Wang, X.; Chen, H.; Shi, Q.; Xiang, J.; Zhang, Y.; Zhu, D.; Zhang, Y. Reduced bioactive gibberellin content in rice seeds under low temperature leads to decreased sugar consumption and low seed germination rates. Plant Physiol. Biochem. 2018, 133, 1–10. [Google Scholar] [CrossRef]
- Obroucheva, N.V.; Sinkevich, I.A.; Lityagina, S.V.; Novikova, G.V. Water relations in germinating seeds. Russ. J. Plant Physiol. 2017, 64, 625–633. [Google Scholar] [CrossRef]
- Chen, B.; Liu, J. Research Progress of Rie Vivipary and Its Regulation. Seed 2017, 36, 49–55. [Google Scholar]
- Sussex, I.M. Growth and metabolism of the embryo and attached seeding of the viviparous mangrove Rhizophora mangle. Am. J. Bot. 1975, 9, 948–953. [Google Scholar] [CrossRef]
- Dong, Y.; Xu, H.; Zhang, H.; Zhang, H.; Wang, F.; Gu, N.; Zhu, Y. Dynamic profile of genes related to seed dormancy under high humidity condition during late stage of rice grain filling. J. Zhejiang Agric. Univ. 2022, 34, 1103–1113. [Google Scholar]
- Toh, S.; Imamura, A.; Watanabe, A.; Nakabayashi, K.; Okamoto, M.; Jikumaru, Y.; Hanada, A.; Aso, Y.; Ishiyama, K.; Tamura, N.; et al. High temperature-induced abscisic acid biosynthesis and its role in the inhibition of gibberellin action in Arabidopsis seeds. Plant Physiol. 2008, 146, 1368–1385. [Google Scholar] [CrossRef]
- Barrero, J.M.; Downie, A.B.; Xu, Q.; Gubler, F. A Role for Barley CRYPTOCHROME1 in Light Regulation of Grain Dormancy and Germination. Plant Cell 2014, 26, 1094–1104. [Google Scholar] [CrossRef]
- Hai, H.; Sechet, J.; Bailly, C.; Leymarie, J.; Corbineau, F. Inhibition of germination of dormant barley (Hordeum vulgare L.) grains by blue light as related to oxygen and hormonal regulation. Plant Cell Environ. 2014, 37, 1393–1403. [Google Scholar]
- Yang, L.; Liu, S.; Lin, R. Advances in Light and Hormones in Regulating Seed Dormancy and Germination. Chin. Bull. Bot. 2019, 5, 569–581. [Google Scholar]
- Ghassemian, M.; Nambara, E.; Cutler, S.; Kawaide, H.; Kamiya, Y.; McCourt, P. Regulation of abscisic acid signaling by the ethylene response pathway in arabidopsis. Plant Cell 2000, 12, 1117–1126. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Wu, X.; Li, X.; Wim, J.J.S.; Cao, H.; Liu, Y. Overexpression of Taetr1-1 promotes enhanced seed dormancy and ethylene insensitivity in wheat. Planta 2023, 258, 56. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Candidate Gene with Physical Position (bp)/Regulatory Gene | Function | Reference |
|---|---|---|---|
| Chinese cabbage | QTL (Chr3 9,859,000–10,632,000) | - | He, 2021 [18] |
| Cucumber | QTL (Chr4 19,973,692–20,505,510) | - | Cao et al., 2021 [20] |
| Oilseed rape | QTLs (qPHS-2-a, qPHS-2-b, qPHS-2-c, qPHS-2-d, qPHS-7) | - | Feng, 2009 [12] |
| Tomato | SlRIN (NC_015442.3) | Keep osmolality in fruit | Wang, 2016 [14] |
| Tomato | SlMET1(NC_015448.3) | Influences the expression of ABA-related genes | Yao et al., 2020 [6] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qu, Y.; Zhang, Y.; Zhang, Z.; Fan, S.; Qi, Y.; Wang, F.; Wang, M.; Feng, M.; Liu, X.; Ren, H. Advance Research on the Pre-Harvest Sprouting Trait in Vegetable Crop Seeds. Int. J. Mol. Sci. 2023, 24, 17171. https://doi.org/10.3390/ijms242417171
Qu Y, Zhang Y, Zhang Z, Fan S, Qi Y, Wang F, Wang M, Feng M, Liu X, Ren H. Advance Research on the Pre-Harvest Sprouting Trait in Vegetable Crop Seeds. International Journal of Molecular Sciences. 2023; 24(24):17171. https://doi.org/10.3390/ijms242417171
Chicago/Turabian StyleQu, Yixin, Yaqi Zhang, Zhongren Zhang, Shanshan Fan, Yu Qi, Fang Wang, Mingqi Wang, Min Feng, Xingwang Liu, and Huazhong Ren. 2023. "Advance Research on the Pre-Harvest Sprouting Trait in Vegetable Crop Seeds" International Journal of Molecular Sciences 24, no. 24: 17171. https://doi.org/10.3390/ijms242417171