

High-Density Chitosan Induces a Biochemical and Molecular Response in Coffea arabica during Infection with Hemileia vastatrix
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Biological Effectiveness Test
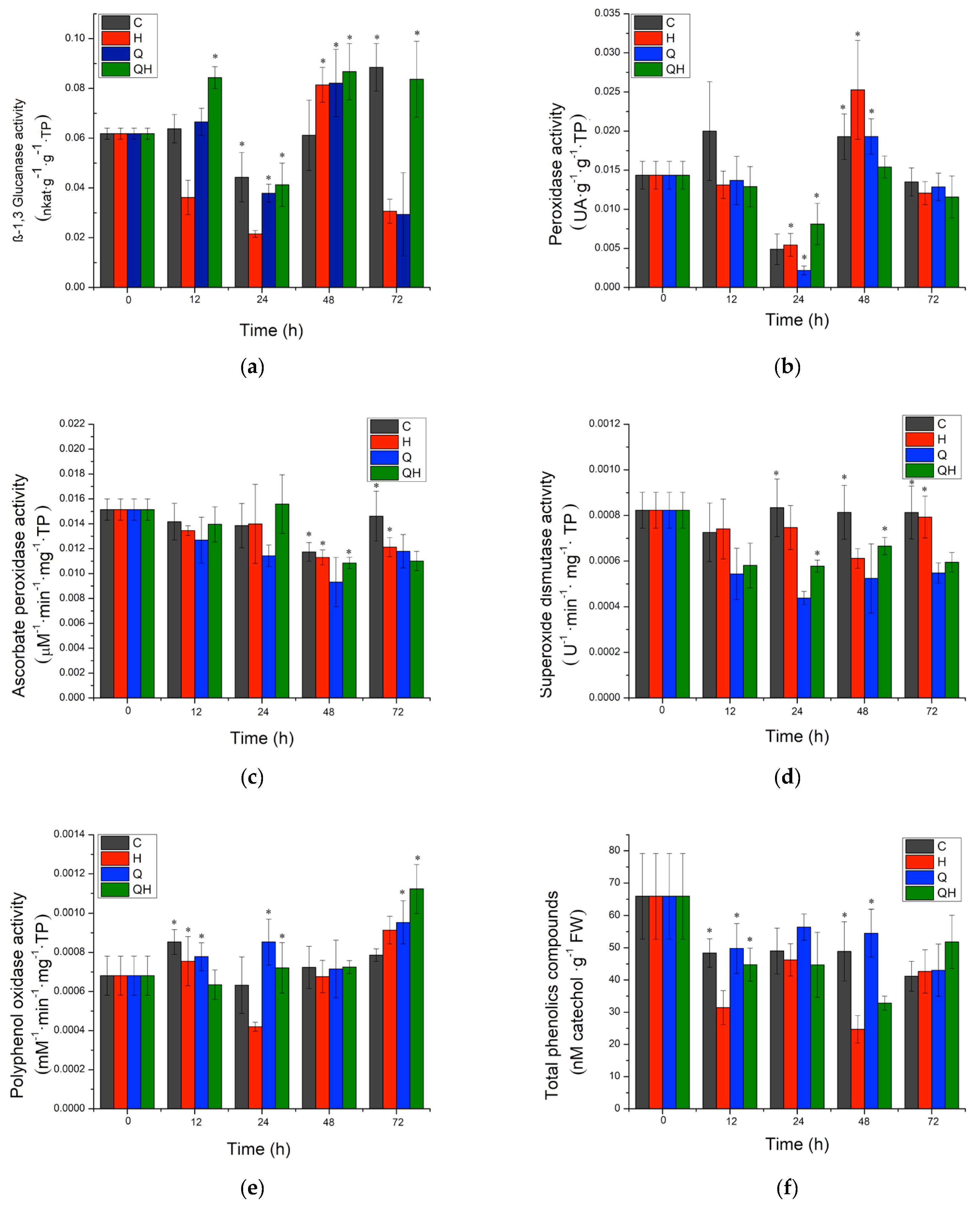
2.2. PR-Protein Activity and Phytoalexin Accumulation
2.3. Quantitative Analysis of Defense-Related Gene Expression by Real-Time PCR
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Biological Material
4.3. High-Density Chitosan Solution
4.4. Biological Effectiveness Test with Food-Grade Chitosan
4.4.1. Disease Incidence
4.4.2. Area under the Disease Progress Curve (AUDPC)
4.4.3. Observation of the Presence of Pathogen in the Plant Tissue
4.5. Enzymatic Activity Evaluation Related to the Pathogenesis and Accumulation of Phytoalexins
4.5.1. β-1,3 Glucanase Activity
4.5.2. Peroxidase Activity
4.5.3. Superoxide Dismutase Activity
4.5.4. Ascorbate Peroxidase Activity
4.5.5. Polyphenol Oxidase Activity
4.5.6. Protein Quantification
4.5.7. Phytoalexin Accumulation
4.6. Quantitative Analysis of Defense-Related Gene Expression by Real-Time PCR
4.6.1. Primers Design
4.6.2. RNA Extraction and cDNA Synthesis
4.6.3. RT qPCR
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- King, H. Trading the Soft Commodity Markets, Investopedia. Available online: https://www.investopedia.com/articles/optioninvestor/07/soft_market.asp (accessed on 19 May 2023).
- ASERCA. Panorama Agropecuario Semanal de Café. Available online: https://info.aserca.gob.mx/panorama/semanal.asp?de=cafe (accessed on 19 May 2023).
- Salcedo-Sarmiento, S.; Aucique-Pérez, C.E.; Silveira, P.R.; Colmán, A.A.; Silva, A.L.; Corrêa-Mansur, P.S.; Rodrigues, F.A.; Evans, H.C.; Barreto, R.W. Elucidating the interactions between the rust Hemileia vastatrix and a Calonectria mycoparasite and the coffee plant. iScience 2021, 24, 102352. [Google Scholar] [CrossRef] [PubMed]
- Gichuru, E.; Alwora, G.; Gimase, J.; Kathurima, C. Coffee Leaf Rust (Hemileia vastatrix) in Kenya—A Review. Agronomy 2021, 11, 2590. [Google Scholar] [CrossRef]
- Barka, G.D.; Caixeta, E.T.; Ferreira, S.S.; Zambolim, L. In silico guided structural and functional analysis of genes with potential involvement in resistance to coffee leaf rust: A functional marker based approach. PLoS ONE 2020, 15, e0222747. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Chen, L.; Zhang, P.; Zhou, J.; Lu, X.; Tian, W. Carbohydrate polymers exhibit great potential as effective elicitors in organic agriculture: A review. Carbohydr. Polym. 2020, 230, 115637. [Google Scholar] [CrossRef] [PubMed]
- Gorni, P.H.; Pacheco, A.C.; Lima-Moro, A.; Silva, J.F.A.; Moreli, R.R.; de Miranda, G.R.; Pelegrini, J.M.; Zaniboni, C.; Spera, K.D.; Junior, J.L.B.; et al. Elicitation Improves the Leaf Area, Enzymatic Activities, Antioxidant Activity and Content of Secondary Metabolites in Achillea millefolium L. Grown in the Field. J. Plant Growth Regul. 2021, 40, 1652–1666. [Google Scholar] [CrossRef]
- Andrade-Monteiro, A.C.; Vilela-de Resende, M.L.; Teixeira-Valente, T.C.; Ribeiro-Junior, P.M.; Foresti-Pereira, V.; Rodrigues-da Costa, J.; Gourlat-da Silva, J.A. Manganese Phosphite in Coffee Defence against Hemileia vastatrix, the Coffee Rust Fungus: Biochemical and Molecular Analyses. J. Phytopathol. 2016, 164, 1043–1053. [Google Scholar] [CrossRef]
- Budzinki, I.G.F.; Camargo, P.O.; Lemos, S.M.C.; Guyot, R.; Calzado, N.F.; Ivamoto-Suzuki, S.T.; Domingues, D.S. Transcriptomic alterations in roots of two contrasting Coffea arabica cultivars after hexanoic acid priming. Front. Genet. 2022, 13, 925811. [Google Scholar] [CrossRef]
- Pereira-Silva, P.D.F.; Vilela-De Resende, M.L.; Reichel, T.; De Lima-Santos, M.; Dos Santos-Botelho, D.M.; Batista-Ferreira, E.; Chagas-Freitas, N. Potassium Phosphite Activates Components Associated with Constitutive Defense Responses in Coffea arabica Cultivars. Mol. Biotechnol. 2023, 65, 1777–1795. [Google Scholar] [CrossRef]
- López-Velázquez, J.C.; Haro-González, J.N.; García-Morales, S.; Espinosa-Andrews, H.; Navarro-López, D.E.; Montero-Cortés, M.I.; Qui-Zapata, J.A. Evaluation of the Physicochemical Properties of Chitosans in Inducing the Defense Response of Coffea arabica against the Fungus Hemileia vastatrix. Polymers 2021, 13, 1940. [Google Scholar] [CrossRef]
- Zheng, K.; Lu, J.; Li, J.; Yu, Y.; Zhang, J.; He, Z.; Ismail, O.M.; Wu, J.; Xie, X.; Li, X.; et al. Efficiency of chitosan application against Phytophthora infestans and the activation of defence mechanisms in potato. Int. J. Biol. Macromol. 2021, 182, 1670–1680. [Google Scholar] [CrossRef]
- Sofi, K.G.; Metzger, C.; Riemann, M.; Nick, P. Chitosan triggers actin remodelling and activation of defence genes that is repressed by calcium influx in grapevine cells. Plant Sci. 2023, 326, 111527. [Google Scholar] [CrossRef] [PubMed]
- López-Velázquez, J.C.; García-Morales, S.; Espinosa-Andrews, H.; Montero-Cortés, M.I.; Qui-Zapata, J.A. Control de la roya del café utilizando moléculas de quitosanos de diferentes orígenes. In Proceedings of the XLIII Encuentro Nacional de la AMIDIQ, Puerto Vallarta, México, 23–26 August 2022. [Google Scholar]
- Jogaiah, S.; Satapute, P.; De Britto, S.; Konappa, N.; Udayashankar, A.C. Exogenous priming of chitosan induces upregulation of phytohormones and resistance against cucumber powdery mildew disease is correlated with localized biosynthesis of defense enzymes. Int. J. Biol. Macromol. 2020, 162, 1825–1838. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Fernandez, M.; Marhuenda-Egea, F.C.; Lopez-Moya, F.; Arnao, M.B.; Cabrera-Escribano, F.; Nueda, M.J.; Gunsé, B.; Lopez-Llorca, L.V. Chitosan Induces Plant Hormones and Defenses in Tomato Root Exudates. Front. Plant Sci. 2020, 11, 572087. [Google Scholar] [CrossRef] [PubMed]
- Soares-Leal, F.D.; Santos-Neto, H.; Lopes-Pinheiro, I.C.; Marques-Oliveira, J.; Alexandre-Pozza, A.A.; Ampélio-Pozza, E. Copper and silver nanoparticles control coffee rust: Decrease the quantity of sprayed active ingredients and is an alternative for sustainable coffee production. Eur. J. Plant Pathol. 2023, 1, 1–13. [Google Scholar] [CrossRef]
- Fajardo-Franco, M.L.; Aguilar-Tlatelpa, M.; Guzmán-Plazola, R.A. Biofungicides evaluation in two coffee cultivars for Hemileia vastatrix control. RMF 2020, 38, 293–306. [Google Scholar] [CrossRef]
- Plaza-Pérez, C.D.; Ampélio-Pozza, E.; Alexandre-Pozza, A.A.; Elmer, W.H.; Bastos-Pereira, A.; Gomes-Guimarães, D.D.S.; Andrade-Monteiro, A.C.; Vilela-De Resende, M.L. Boron, zinc and manganese suppress rust on coffee plants grown in a nutrient solution. Eur. J. Plant Pathol. 2020, 156, 727–738. [Google Scholar] [CrossRef]
- Tugizimana, F.; Mhlongo, M.I.; Piater, L.A.; Dubery, I.A. Metabolomics in Plant Priming Research: The Way Forward? Int. J. Mol. Sci. 2018, 19, 1759. [Google Scholar] [CrossRef]
- Gangireddygari, V.S.R.; Chung, B.N.; Cho, I.S.; Yoon, J.Y. Inhibitory Effect of Chitosan and Phosphate Cross-linked Chitosan against Cucumber Mosaic Virus and Pepper Mild Mottle Virus. Plant Pathol. J. 2021, 37, 632–640. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, X.; Kennedy, J.F.; Jiang, M.; Cai, Q.; Wu, X. Chitosan induces resistance to tuber rot in stored potato caused by Alternaria tenuissima. Int. J. Biol. Macromol. 2019, 140, 851–857. [Google Scholar] [CrossRef]
- Liu, Y.; Wisniewski, M.; Kennedy, J.F.; Jiang, Y.; Tang, J.; Liu, J. Chitosan and oligochitosan enhance ginger (Zingiber officinale Roscoe) resistance to rhizome rot caused by Fusarium oxysporum in storage. Carbohydr. Polym. 2016, 151, 474–479. [Google Scholar] [CrossRef]
- Samarah, N.H.; AL-Quraan, N.A.; Massad, R.S.; Welbaum, G.E. Treatment of bell pepper (Capsicum annuum L.) seeds with chitosan increases chitinase and glucanase activities and enhances emergence in a standard cold test. Sci. Hortic. 2020, 269, 109393. [Google Scholar] [CrossRef]
- Khan, M.; Ali, S.; Al Azzawi, T.N.I.; Saqib, S.; Ullah, F.; Ayaz, A.; Zaman, W. The Key Roles of ROS and RNS as a Signaling Molecule in Plant–Microbe Interactions. Antioxidants 2023, 12, 268. [Google Scholar] [CrossRef]
- Al-Mokadem, A.Z.; Alnaggar, A.E.A.M.; Mancy, A.G.; Sofy, A.R.; Sofy, M.R.; Mohamed, A.K.S.H.; Abou Ghazala, M.M.A.; El-Zabalawy, K.M.; Salem, N.F.G.; Elnosary, M.E.; et al. Foliar Application of Chitosan and Phosphorus Alleviate the Potato virus Y-Induced Resistance by Modulation of the Reactive Oxygen Species, Antioxidant Defense System Activity and Gene Expression in Potato. Agronomy 2022, 12, 3064. [Google Scholar] [CrossRef]
- Kidwai, M.; Ahmad, I.Z.; Chakrabarty, D. Class III peroxidase: An indispensable enzyme for biotic/abiotic stress tolerance and a potent candidate for crop improvement. Plant Cell Rep. 2020, 39, 1381–1393. [Google Scholar] [CrossRef]
- Fischer, I.H.; Morales, J.V.P.; da Silva, L.M.; de Almeida Bertani, R.M.; Deus, A.C.F.; Pascholati, S.F. Activity of Phosphites and Chitosan on Biochemical Responses and Target Spot Control in Cucumber Plants. Gesunde Pflanz. 2023, 75, 825–835. [Google Scholar] [CrossRef]
- Luján-Hidalgo, M.C.; Jiménez-Aguilar, L.A.; Ruiz-Lau, N.; Reyes-Zambrano, S.J.; Gutiérrez-Miceli, F.A. Cambios bioquímicos en respuesta al ataque de roya en plantaciones de café. Polibotánica 2020, 49, 149–160. [Google Scholar] [CrossRef]
- Varghese, L.; Thomas, G. Chitosan triggers tolerance to Pythium myriotylum infection in ginger (Zingiber officinale) by modulating multiple defense signaling pathways. Physiol. Mol. Plant Pathol. 2023, 125, 101983. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Zulfiqar, F.; Raza, A.; Mohsin, S.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhao, P.; Zhang, P.; Su, L.; Jia, H.; Wei, X.; Fang, J.; Jia, H. Integrative transcriptomics and metabolomics data exploring the effect of chitosan on postharvest grape resistance to Botrytis cinerea. Postharvest Biol. Technol. 2020, 167, 111248. [Google Scholar] [CrossRef]
- Mapuranga, J.; Zhang, N.; Zhang, L.; Chang, J.; Yang, W. Infection Strategies and Pathogenicity of Biotrophic Plant Fungal Pathogens. Front. Microbiol. 2022, 13, 799396. [Google Scholar] [CrossRef]
- Guo, Y.; Zhou, J.; Zhang, J.; Zhang, S. Chitosan combined with sodium silicate treatment induces resistance against rot caused by Alternaria alternata in postharvest jujube fruit. J. Phytopathol. 2019, 167, 451–460. [Google Scholar] [CrossRef]
- Zhang, P.; Jia, H.; Gong, P.; Ehsan, S.; Pang, Q.; Dong, T.; Li, T.; Jin, H.; Fang, J. Chitosan induces jasmonic acid production leading to resistance of ripened fruit against Botrytis cinerea infection. Food Chem. 2021, 337, 127772. [Google Scholar] [CrossRef]
- Godana, E.A.; Yang, Q.; Zhao, L.; Zhang, X.; Liu, J.; Zhang, H. Pichia anomala Induced With Chitosan Triggers Defense Response of Table Grapes Against Post-harvest Blue Mold Disease. Front. Microbiol. 2021, 12, 704519. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Suleiman, S.; Ahmad, M. Induction of Biochemical Changes in Santa Teriza Lime Leaves by Chitosan Application Influence Citrus Leaf Miner Damage. Am. J. Plant Sci. 2022, 13, 403–415. [Google Scholar] [CrossRef]
- Mesén, F.; Jiménez, L.D. Producción de Clones de Café Por Miniestacas. Available online: https://repositorio.catie.ac.cr/bitstream/handle/11554/8176/Produccion_de_clones_de_cafe.pdf?sequence=1&isAllowed=y (accessed on 22 May 2023).
- Ferreira-Possa, K.; Gourlat-Silva, J.A.; Vilela-Resende, M.L.; Tenente, R.; Pinheiro, C.; Chaves, I.; Planchon, S.; Andrade-Monteiro, A.C.; Renaut, J.; Figueiredo-Carvalho, M.A.; et al. Primary Metabolism Is Distinctly Modulated by Plant Resistance Inducers in Coffea arabica Leaves Infected by Hemileia vastatrix. Front. Plant Sci. 2020, 11, 309. [Google Scholar] [CrossRef] [PubMed]
- SENASICA. Roya del Cafeto. Hemileia vastatrix Berkeley & Broome Ficha Técnica No. 40. Available online: http://www.cesavep.org/descargas/RDC/Ficha_Tecnica_Roya_del_cafeto.pdf (accessed on 22 May 2023).
- Edy, N.; Anshary, A.; Basir-Cyio, M.M.; Lakani, I.; Kadir, S.R.A.; Mahmud, S. Incidence and Severity of Ganoderma Rot Disease in Tropical Land-use Systems and Their Virulence to Palm Oil. Plant Pathol. J. 2020, 19, 98–105. [Google Scholar] [CrossRef]
- Bocianowski, J.; Tratwal, A.; Nowosad, K. Genotype by environment interaction for area under the disease-progress curve (AUDPC) value in spring barley using additive main effects and multiplicative interaction model. Australas. Plant Pathol. 2020, 49, 525–529. [Google Scholar] [CrossRef]
- López-Velázquez, J.C.; Rodríguez-Rodríguez, R.; Espinosa-Andrews, H.; Qui-Zapata, J.A.; García-Morales, S.; Navarro-López, D.E.; Luna-Bárcenas, G.; Vassallo-Brigneti, E.C.; García-Carvajal, Z.Y. Gelatin–chitosan–PVA hydrogels and their application in agriculture. J. Chem. Technol. Biotechnol. 2019, 94, 3495–3504. [Google Scholar] [CrossRef]
- Kruger, N.J. The Bradford Method for Protein Quantitation. In The Protein Protocols Handbook, 3rd ed.; Walker, J.M., Ed.; Springer Protocols Handbooks; Humana Press: Totowa, NJ, USA, 2009; Volume 1, pp. 17–24. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef]
- Meng, L.; Feldman, L. A rapid TRIzol-based two-step method for DNA-free RNA extraction from Arabidopsis siliques and dry seeds. Biotechnol. J. 2010, 5, 183–186. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Disease Incidence (%) | AUDPC (au) |
|---|---|---|
| C | 0 | 0.0 ± 0.0 c |
| H | 100 | 102 ± 14.3 a |
| Q1 | 100 | 40.0 ± 20.6 b |
| Q5 | 100 | 25.0 ± 11.66 b |
| Genes | Oligonucleotide Sequences (5′-3′) | Size (pb) | Accession Number |
|---|---|---|---|
| Peroxidase (POX) | FW: GTGGATGCGGAGTACCTGAA RV: AACCGTTTGGACCTCCAGTT | 153 | evm.TU.Scaffold_618.242 |
| Catalase (CAT) | FW: GATGCACCCAATTCCTTCTGC RV: CCCAGCGACAGATAAAGCG | 140 | evm.TU.Scaffold_2256.195 |
| ß-1,3 glucanase | FW: GGGTGACCCTACAAAAGCCA RV: GGCCTGGAGGAAAGGTTCAT | 121 | evm.TU.Scaffold_517.1 |
| Phenylalanine ammonia lyase (PAL) | FW: CTTGTGAGGGGAGAGTTGGG RV: GGTAGGTGGCTCTTGTCAGC | 112 | evm.TU.Scaffold_618.1114 |
| NPR1 | FW: TTGGTTATGAGGCTGCTGCT RV: GGCTTTAGATGCTGCAAGGC | 138 | evm.model.Scaffold_523.152 |
| GAPDH | FW: CCCTTGGGGTGAAACTGGAG RV: AACATGGGTGCATCCTTGCT | 138 | evm.model.Scaffold_608.230 |
| Actin 8 | FW: ATTAGCTCGACAAGACGCCC RV: CTCACGTTCCATGTGTTGCG | 142 | evm.model.Scaffold_315.601 |
| 14-3-3 | FW: GCTGAGTTCAAAACTGGGGCTG RV: ATTGGGTGTGTTGGAGCCAG | 110 | evm.model.Scaffold_2421.275 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Velázquez, J.C.; García-Morales, S.; López-Sánchez, G.P.; Montero-Cortés, M.I.; Uc-Várguez, A.; Qui-Zapata, J.A. High-Density Chitosan Induces a Biochemical and Molecular Response in Coffea arabica during Infection with Hemileia vastatrix. Int. J. Mol. Sci. 2023, 24, 16165. https://doi.org/10.3390/ijms242216165
López-Velázquez JC, García-Morales S, López-Sánchez GP, Montero-Cortés MI, Uc-Várguez A, Qui-Zapata JA. High-Density Chitosan Induces a Biochemical and Molecular Response in Coffea arabica during Infection with Hemileia vastatrix. International Journal of Molecular Sciences. 2023; 24(22):16165. https://doi.org/10.3390/ijms242216165
Chicago/Turabian StyleLópez-Velázquez, Julio César, Soledad García-Morales, Gloria Paola López-Sánchez, Mayra Itzcalotzin Montero-Cortés, Alberto Uc-Várguez, and Joaquín Alejandro Qui-Zapata. 2023. "High-Density Chitosan Induces a Biochemical and Molecular Response in Coffea arabica during Infection with Hemileia vastatrix" International Journal of Molecular Sciences 24, no. 22: 16165. https://doi.org/10.3390/ijms242216165