

Potential Binding Sites of Pharmacological Chaperone NCGC00241607 on Mutant β-Glucocerebrosidase and Its Efficacy on Patient-Derived Cell Cultures in Gaucher and Parkinson’s Disease
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
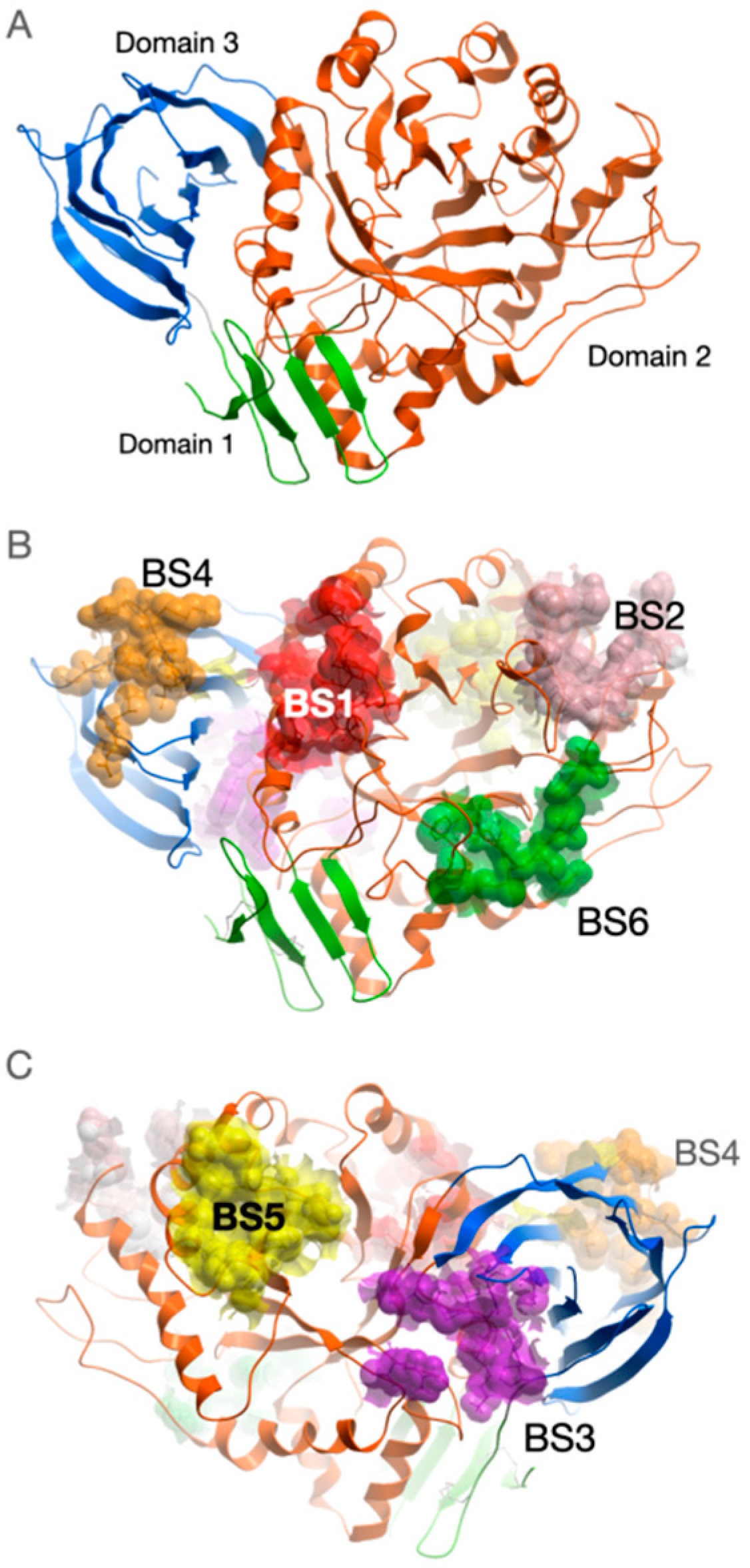
2.1. Binding Sites Description
2.2. NCGC607
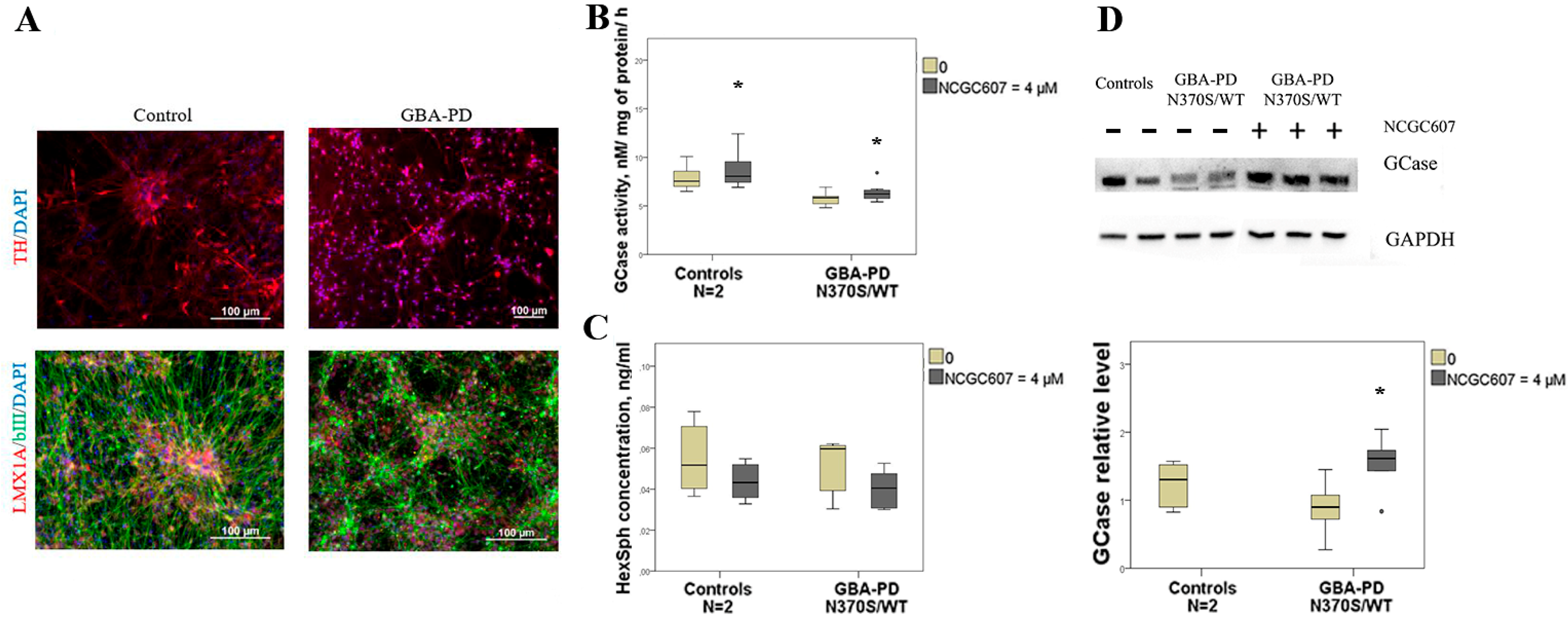
2.3. The Effects of NCGC607 Treatment in Cultured Macrophages and iPSC-Derived DA Neurons from GD and GBA-PD Patients
2.4. The Effects of NCGC607 Treatment in Cultured Macrophages from GD and GBA-PD Patients
2.4.1. GCase Activity and Protein Levels
2.4.2. HexSph Concentration
2.5. The Effects of NCGC607 Treatment in iPSC-Derived DA Neurons from GBA-PD Patient
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Differentiation of PBMCs to Macrophages
4.3. Generation of iPSC and Culture Conditions
4.4. Differentiation of Patient-Specific iPSCs to DA Neurons
4.5. Chemical Synthesis of NCGC607
4.6. Chaperone Treatment of Cultured Macrophages and DA Neurons Derived from Patient-Specific iPSCs
4.7. Quantification of GCase Activity and Hexosylsphingosine (HexSph) Concentration
4.8. Western Blot Analysis
4.9. Molecular Modeling
4.9.1. Construction of N370S GCase and PC NCGC607 Atomistic Models
4.9.2. Molecular Dynamics Simulations
4.9.3. Molecular Docking
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Do, J.; McKinney, C.; Sharma, P.; Sidransky, E. Glucocerebrosidase and Its Relevance to Parkinson Disease. Mol. Neurodegener. 2019, 14, 36. [Google Scholar] [CrossRef] [PubMed]
- Germain, D.P. Gaucher’s Disease: A Paradigm for Interventional Genetics. Clin. Genet. 2004, 65, 77–86. [Google Scholar] [CrossRef] [PubMed]
- The Human Gene Mutation Database. HGMD Professional 2021.4.1. Available online: https://www.hgmd.cf.ac.uk (accessed on 19 May 2023).
- Parlar, S.C.; Grenn, F.P.; Kim, J.J.; Baluwendraat, C.; Gan-Or, Z. Classification of GBA1 Variants in Parkinson’s Disease: The GBA1-PD Browser. Mov. Disord. 2023, 38, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Balwani, M.; Fuerstman, L.; Kornreich, R.; Edelmann, L.; Desnick, R.J. Type 1 Gaucher Disease: Significant Disease Manifestations in “Asymptomatic” Homozygotes Identified by Prenatal Carrier Screening. Arch. Intern. Med. 2010, 170, 1463. [Google Scholar] [CrossRef]
- Goker-Alpan, O.; Schiffmann, R.; Park, J.K.; Stubblefield, B.K.; Tayebi, N.; Sidransky, E. Phenotypic Continuum in Neuronopathic Gaucher Disease: An Intermediate Phenotype between Type 2 and Type 3. J. Pediatr. 2003, 143, 273–276. [Google Scholar] [CrossRef]
- Beutler, E.; Grabowski, G.A. Gaucher Disease. In The Metabolic and Molecular Bases of Inherited Disease; Scriver, C.R., Beaudet, A.L., Sly, W.S., Valle, D., Eds.; McGraw-Hill: New York, NY, USA, 2001. [Google Scholar]
- Charrow, J.; Andersson, H.C.; Kaplan, P.; Kolodny, E.H.; Mistry, P.; Pastores, G.; Rosenbloom, B.E.; Scott, C.R.; Wappner, R.S.; Weinreb, N.J.; et al. The Gaucher Registry: Demographics and Disease Characteristics of 1698 Patients With Gaucher Disease. Arch. Intern. Med. 2000, 160, 2835–2843. [Google Scholar] [CrossRef]
- Gan-Or, Z.; Amshalom, I.; Kilarski, L.L.; Bar-Shira, A.; Gana-Weisz, M.; Mirelman, A.; Marder, K.; Bressman, S.; Giladi, N.; Orr-Urtreger, A. Differential Effects of Severe vs Mild GBA Mutations on Parkinson Disease. Neurology 2015, 84, 880–887. [Google Scholar] [CrossRef]
- Balestrino, R.; Schapira, A.H.V. Parkinson Disease. Eur. J. Neurol. 2020, 27, 27–42. [Google Scholar] [CrossRef]
- Emelyanov, A.K.; Usenko, T.S.; Tesson, C.; Senkevich, K.A.; Nikolaev, M.A.; Miliukhina, I.V.; Kopytova, A.E.; Timofeeva, A.A.; Yakimovsky, A.F.; Lesage, S.; et al. Mutation Analysis of Parkinson’s Disease Genes in a Russian Data Set. Neurobiol. Aging 2018, 71, 267.e7. [Google Scholar] [CrossRef]
- Sidransky, E.; Nalls, M.A.; Aasly, J.O.; Aharon-Peretz, J.; Annesi, G.; Barbosa, E.R.; Bar-Shira, A.; Berg, D.; Bras, J.; Brice, A.; et al. Multi-Center Analysis of Glucocerebrosidase Mutations in Parkinson Disease. N. Engl. J. Med. 2009, 361, 1651. [Google Scholar] [CrossRef]
- Smith, L.; Schapira, A.H.V. GBA Variants and Parkinson Disease: Mechanisms and Treatments. Cells 2022, 11, 1261. [Google Scholar] [CrossRef] [PubMed]
- Schapira, A.H.V. Glucocerebrosidase and Parkinson Disease: Recent Advances. Mol. Cell. Neurosci. 2015, 66, 37. [Google Scholar] [CrossRef] [PubMed]
- Toffoli, M.; Vieira, S.R.L.; Schapira, A.H.V. Genetic Causes of PD: A Pathway to Disease Modification. Neuropharmacology 2020, 170, 108022. [Google Scholar] [CrossRef] [PubMed]
- Senkevich, K.; Rudakou, U.; Gan-Or, Z. New Therapeutic Approaches to Parkinson’s Disease Targeting GBA, LRRK2 and Parkin. Neuropharmacology 2022, 202, 108822. [Google Scholar] [CrossRef] [PubMed]
- Zimran, A.; Elstein, D.; Gonzalez, D.E.; Lukina, E.A.; Qin, Y.; Dinh, Q.; Turkia, H. Ben Treatment-Naïve Gaucher Disease Patients Achieve Therapeutic Goals and Normalization with Velaglucerase Alfa by 4years in Phase 3 Trials. Blood Cells Mol. Dis. 2018, 68, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Han, T.U.; Sam, R.; Sidransky, E. Small Molecule Chaperones for the Treatment of Gaucher Disease and GBA1-Associated Parkinson Disease. Front. Cell Dev. Biol. 2020, 8, 271. [Google Scholar] [CrossRef]
- Tran, M.L.; Génisson, Y.; Ballereau, S.; Dehoux, C. Second-Generation Pharmacological Chaperones: Beyond Inhibitors. Molecules 2020, 25, 3145. [Google Scholar] [CrossRef]
- Maegawa, G.H.B.; Tropak, M.B.; Buttner, J.D.; Rigat, B.A.; Fuller, M.; Pandit, D.; Tang, L.; Kornhaber, G.J.; Hamuro, Y.; Clarke, J.T.R.; et al. Identification and Characterization of Ambroxol as an Enzyme Enhancement Agent for Gaucher Disease. J. Biol. Chem. 2009, 284, 23502–23516. [Google Scholar] [CrossRef]
- Martínez-Bailén, M.; Clemente, F.; Matassini, C.; Cardona, F. GCase Enhancers: A Potential Therapeutic Option for Gaucher Disease and Other Neurological Disorders. Pharmaceuticals 2022, 15, 823. [Google Scholar] [CrossRef]
- Mullin, S.; Smith, L.; Lee, K.; D’Souza, G.; Woodgate, P.; Elflein, J.; Hällqvist, J.; Toffoli, M.; Streeter, A.; Hosking, J.; et al. Ambroxol for the Treatment of Patients with Parkinson Disease with and Without Glucocerebrosidase Gene Mutations: A Nonrandomized, Noncontrolled Trial. JAMA Neurol. 2020, 77, 427–434. [Google Scholar] [CrossRef]
- Zimran, A.; Altarescu, G.; Elstein, D. Pilot Study Using Ambroxol as a Pharmacological Chaperone in Type 1 Gaucher Disease. Blood Cells Mol. Dis. 2013, 50, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Kopytova, A.E.; Rychkov, G.N.; Nikolaev, M.A.; Baydakova, G.V.; Cheblokov, A.A.; Senkevich, K.A.; Bogdanova, D.A.; Bolshakova, O.I.; Miliukhina, I.V.; Bezrukikh, V.A.; et al. Ambroxol Increases Glucocerebrosidase (GCase) Activity and Restores GCase Translocation in Primary Patient-Derived Macrophages in Gaucher Disease and Parkinsonism. Park. Relat. Disord. 2021, 84, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Aflaki, E.; Borger, D.K.; Moaven, N.; Stubblefield, B.K.; Rogers, S.A.; Patnaik, S.; Schoenen, F.J.; Westbroek, W.; Zheng, W.; Sullivan, P.; et al. A New Glucocerebrosidase Chaperone Reduces α-Synuclein and Glycolipid Levels in IPSC-Derived Dopaminergic Neurons from Patients with Gaucher Disease and Parkinsonism. J. Neurosci. 2016, 36, 7441–7452. [Google Scholar] [CrossRef] [PubMed]
- Rogers, S.; Patnaik, S.; Schoenen, F.; Zheng, W.; Choi, J.; Motabar, O.; Southall, N.; Westbroek, W.; Goldin, E.; Sidransky, E.; et al. Discovery, SAR, and Biological Evaluation of Non-Inhibitory Chaperones of Glucocerebrosidase. In Probe Reports from the NIH Molecular Libraries Program; National Center for Biotechnology Information: Bethesda, MD, USA, 2013. [Google Scholar]
- Aflaki, E.; Goldin, E.; Marugan, J.J.; Patnaik, S.; Rogers, S.A.; Schoenen, F.J.; Sidransky, E.; Southall, N.; Westbroek, W.; Zheng, W. Salicylic Acid Derivatives Useful as Glucocerebrosidase Activators. U.S. Patent US2015065469A1, 5 March 2015. [Google Scholar]
- Patnaik, S.; Zheng, W.; Choi, J.H.; Motabar, O.; Southall, N.; Westbroek, W.; Lea, W.A.; Velayati, A.; Goldin, E.; Sidransky, E.; et al. Discovery, SAR, and Biological Evaluation of Non-Inhibitory Small Molecule Chaperones of Glucocerebrosidase. J. Med. Chem. 2012, 55, 5734. [Google Scholar] [CrossRef] [PubMed]
- Aflaki, E.; Moaven, N.; Borger, D.K.; Lopez, G.; Westbroek, W.; Chae, J.J.; Marugan, J.; Patnaik, S.; Maniwang, E.; Gonzalez, A.N.; et al. Lysosomal Storage and Impaired Autophagy Lead to Inflammasome Activation in Gaucher Macrophages. Aging Cell 2016, 15, 77–88. [Google Scholar] [CrossRef]
- Mazzulli, J.R.; Zunke, F.; Tsunemi, T.; Toker, N.J.; Jeon, S.; Burbulla, L.F.; Patnaik, S.; Sidransky, E.; Marugan, J.J.; Sue, C.M.; et al. Activation of β-Glucocerebrosidase Reduces Pathological α-Synuclein and Restores Lysosomal Function in Parkinson’s Patient Midbrain Neurons. J. Neurosci. 2016, 36, 7693. [Google Scholar] [CrossRef]
- Aflaki, E.; Stubblefield, B.K.; Maniwang, E.; Lopez, G.; Moaven, N.; Goldin, E.; Marugan, J.; Patnaik, S.; Dutra, A.; Southall, N.; et al. Macrophage Models of Gaucher Disease for Evaluating Disease Pathogenesis and Candidate Drugs. Sci. Transl. Med. 2014, 6, 240ra73. [Google Scholar] [CrossRef]
- Offman, M.N.; Silman, I.; Sussman, J.L.; Futerman, A.H. Crystal Structure of the Enzyme Acid B-Glucosidase. In Gaucher Disease: Basic and Clinical Perspectives; Future Medicine Ltd: London, UK, 2013; pp. 125–138. [Google Scholar] [CrossRef]
- Toffoli, M.; Chen, X.; Sedlazeck, F.J.; Lee, C.-Y.; Mullin, S.; Higgins, A.; Koletsi, S.; Garcia-Segura, M.E.; Sammler, E.; Scholz, S.W.; et al. Comprehensive Short and Long Read Sequencing Analysis for the Gaucher and Parkinson’s Disease-Associated GBA Gene. Commun. Biol. 2022, 5, 670. [Google Scholar] [CrossRef]
- Nikolaev, M.A.; Kopytova, A.E.; Baidakova, G.V.; Emel’yanov, A.K.; Salogub, G.N.; Senkevich, K.A.; Usenko, T.S.; Gorchakova, M.V.; Koval’chuk, Y.P.; Berkovich, O.A.; et al. Human Peripheral Blood Macrophages As a Model for Studying Glucocerebrosidase Dysfunction. Cell Tissue Biol. 2019, 13, 100–106. [Google Scholar] [CrossRef]
- Muñoz, S.S.; Petersen, D.; Marlet, F.R.; Kücükköse, E.; Galvagnion, C. The Interplay between Glucocerebrosidase, α-Synuclein and Lipids in Human Models of Parkinson’s Disease. Biophys. Chem. 2021, 273, 106534. [Google Scholar] [CrossRef]
- Ivanova, M.M.; Changsila, E.; Turgut, A.; Goker-Alpan, O. Individualized Screening for Chaperone Activity in Gaucher Disease Using Multiple Patient Derived Primary Cell Lines. Am. J. Transl. Res. 2018, 10, 3750–3761. [Google Scholar] [CrossRef] [PubMed]
- Luan, Z.; Li, L.; Higaki, K.; Nanba, E.; Suzuki, Y.; Ohno, K. The Chaperone Activity and Toxicity of Ambroxol on Gaucher Cells and Normal Mice. Brain Dev. 2013, 35, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Schueler, U.H.; Kolter, T.; Kaneski, C.R.; Zirzow, G.C.; Sandhoff, K.; Brady, R.O. Correlation between Enzyme Activity and Substrate Storage in a Cell Culture Model System for Gaucher Disease. J. Inherit. Metab. Dis. 2004, 27, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, R.L. A Guided Tour of the Structural Biology of Gaucher Disease: Acid-β-Glucosidase and Saposin C. Enzym. Res. 2011, 2011, 15. [Google Scholar] [CrossRef]
- Benz, J.; Rufer, A.C.; Huber, S.; Ehler, A.; Hug, M.; Topp, A.; Guba, W.; Hofmann, E.C.; Jagasia, R.; Rodríguez Sarmiento, R.M. Novel β-Glucocerebrosidase Activators That Bind to a New Pocket at a Dimer Interface and Induce Dimerization. Angew. Chem. Int. Ed. 2021, 60, 5436–5442. [Google Scholar] [CrossRef]
- Zheng, J.; Chen, L.; Skinner, O.S.; Ysselstein, D.; Remis, J.; Lansbury, P.; Skerlj, R.; Mrosek, M.; Heunisch, U.; Krapp, S.; et al. β-Glucocerebrosidase Modulators Promote Dimerization of β-Glucocerebrosidase and Reveal an Allosteric Binding Site. J. Am. Chem. Soc. 2018, 140, 5914–5924. [Google Scholar] [CrossRef]
- Rao, G.; Fisch, L.; Srinivasan, S.; D’Amico, F.; Okada, T.; Eaton, C.; Robbins, C. Does This Patient Have Parkinson Disease? JAMA 2003, 289, 347–353. [Google Scholar] [CrossRef]
- Grigor’eva, E.V.; Drozdova, E.S.; Sorogina, D.A.; Malakhova, A.A.; Pavlova, S.V.; Vyatkin, Y.V.; Khabarova, E.A.; Rzaev, J.A.; Medvedev, S.P.; Zakian, S.M. Generation of Induced Pluripotent Stem Cell Line, ICGi034-A, by Reprogramming Peripheral Blood Mononuclear Cells from a Patient with Parkinson’s Disease Associated with GBA Mutation. Stem Cell Res. 2022, 59, 102651. [Google Scholar] [CrossRef]
- Grigor’eva, E.V.; Kopytova, A.E.; Yarkova, E.S.; Pavlova, S.V.; Sorogina, D.A.; Malakhova, A.A.; Malankhanova, T.B.; Baydakova, G.V.; Zakharova, E.Y.; Medvedev, S.P.; et al. Biochemical Characteristics of IPSC-Derived Dopaminergic Neurons from N370S GBA Variant Carriers with and without Parkinson’s Disease. Int. J. Mol. Sci. 2023, 24, 4437. [Google Scholar] [CrossRef]
- de Rus Jacquet, A. Preparation and Co-Culture of IPSC-Derived Dopaminergic Neurons and Astrocytes. Curr. Protoc. Cell Biol. 2019, 85, e98. [Google Scholar] [CrossRef]
- Grigor’eva, E.V.; Malankhanova, T.B.; Surumbayeva, A.; Pavlova, S.V.; Minina, J.M.; Kizilova, E.A.; Suldina, L.A.; Morozova, K.N.; Kiseleva, E.; Sorokoumov, E.D.; et al. Generation of GABAergic Striatal Neurons by a Novel IPSC Differentiation Protocol Enabling Scalability and Cryopreservation of Progenitor Cells. Cytotechnology 2020, 72, 649–663. [Google Scholar] [CrossRef] [PubMed]
- Pchelina, S.; Emelyanov, A.; Baydakova, G.; Andoskin, P.; Senkevich, K.; Nikolaev, M.; Miliukhina, I.; Yakimovskii, A.; Timofeeva, A.; Fedotova, E.; et al. Oligomeric α-Synuclein and Glucocerebrosidase Activity Levels in GBA-Associated Parkinson’s Disease. Neurosci. Lett. 2017, 636, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Pchelina, S.; Baydakova, G.; Nikolaev, M.; Senkevich, K.; Emelyanov, A.; Kopytova, A.; Miliukhina, I.; Yakimovskii, A.; Timofeeva, A.; Berkovich, O.; et al. Blood Lysosphingolipids Accumulation in Patients with Parkinson’s Disease with Glucocerebrosidase 1 Mutations. Mov. Disord. 2018, 33, 1325–1330. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.K.; Elbin, C.S.; Chuang, W.L.; Cooper, S.K.; Marashio, C.A.; Beauregard, C.; Keutzer, J.M. Multiplex Enzyme Assay Screening of Dried Blood Spots for Lysosomal Storage Disorders by Using Tandem Mass Spectrometry. Clin. Chem. 2008, 54, 1725–1728. [Google Scholar] [CrossRef]
- Polo, G.; Burlina, A.P.; Kolamunnage, T.B.; Zampieri, M.; Dionisi-Vici, C.; Strisciuglio, P.; Zaninotto, M.; Plebani, M.; Burlina, A.B. Diagnosis of Sphingolipidoses: A New Simultaneous Measurement of Lysosphingolipids by LC-MS/MS. Clin. Chem. Lab. Med. 2017, 55, 403–414. [Google Scholar] [CrossRef]
- Abagyan, R.; Totrov, M.; Kuznetsov, D. ICM—A New Method for Protein Modeling and Design: Applications to Docking and Structure Prediction from the Distorted Native Conformation. J. Comput. Chem. 1994, 15, 488–506. [Google Scholar] [CrossRef]
- Schmidt, M.W.; Baldridge, K.K.; Boatz, J.A.; Elbert, S.T.; Gordon, M.S.; Jensen, J.H.; Koseki, S.; Matsunaga, N.; Nguyen, K.A.; Su, S.; et al. General Atomic and Molecular Electronic Structure System. J. Comput. Chem. 1993, 14, 1347–1363. [Google Scholar] [CrossRef]
- Bottegoni, G.; Kufareva, I.; Totrov, M.; Abagyan, R. Four-Dimensional Docking: A Fast and Accurate Account of Discrete Receptor Flexibility in Ligand Docking. J. Med. Chem. 2009, 52, 397. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Residues | Binding Free Energy, kcal/mol |
|---|---|---|
| BS1 | S366, T369, S370, Q362, G344, V343, W312, D315, F316, A318, P319, A320 | −53.7 ± 4.0 |
| BS2 | K194, G195, S196, L197, G199, Q200, G239, L249, G250, F251, T252, H255 | −48.7 ± 2.5 |
| BS3 | E429, P428, I427, F426, K425, P29, R170, H451, S455 | −35.3 ± 2.3 |
| BS4 | G64, L65, D467, V468, D443, D444, N442, K441, Q440, S439 | −32.1 ± 2.5 |
| BS5 | H274, V276, R277, L278, H306, K303, Y304, V305, G265, L268, A269 | −34.3 ± 3.2 |
| BS6 | F128, S129, I130, R131, T132, N192, P391 | −29.00 ± 2.9 |
| Patients | GD Type | Type of Therapy | Sex | Age, Years | Mutation in the GBA1 Gene | GCase Residual Activity in Blood, % | HexSph Concentration in Blood, % |
|---|---|---|---|---|---|---|---|
| GD 1 | 1 | ERT | m | 55 | N370S/c84dupG | 0.4 | 6795 |
| GD 2 | 1 | ERT | f | 48 | N370S/L444P | 4.74 | 16,216 |
| GD 3 | 1 | SRT | f | 20 | N370S/L444P | 9.3 | 13,281 |
| GD 4 | 1 | ERT | f | 31 | N370S/L444P | 3.2 | 15,212 |
| GD 5 | 1 | ERT | m | 50 | N370S/- | 4.8 | 1738 |
| GD 6 | 1 | ERT | f | 35 | N370S/R120W | 2.8 | 11,003 |
| GD 7 | 1 | ERT | m | 29 | N370S/L444P | 5.9 | 27,413 |
| GD 8 | 1 | - | m | 26 | N370S/R120W | 10.3 | 45,637 |
| GD 9 | 1 | ERT | f | 42 | N370S/W184R | 5.1 | 21,042 |
| GBA-PD 1 | - | L-DOPA | f | 62 | WT/L444P | 52.6 | 320 |
| GBA-PD 2 | - | L-DOPA | f | 50 | WT/L444P | 50.1 | 192 |
| GBA-PD 3 | - | L-DOPA | m | 42 | N370S/WT | 78.8 | 294 |
| GBA-PD 4 | - | L-DOPA | f | 66 | N370S/WT | 54.7 | 219 |
| GBA-PD 5 | - | L-DOPA | f | 57 | N370S/WT | 55.6 | 357 |
| GBA-PD 6 | - | L-DOPA | f | 56 | N370S/WT | 38.1 | 201 |
| Controls (n = 7) | - | - | 42% men | 41.3 ± 4.2 | WT/WT | 100 | 100 |
| Antibodies | Company | Cat. Ref. | Raised/Type | Dilution |
|---|---|---|---|---|
| Primary antibodies | ||||
| Anti-TH | Millipore, Burlington, VT, USA | AB152 | IgG rabbit polyclonal | 1:400 |
| Anti-LMX1A | Abcam, Cambridge, UK | ab139726 | IgG rabbit polyclonal | 1:100 |
| Anti-TUJ1 | Covance, Princeton, NJ, USA | MMS-435P | IgG2a mouse monoclonal | 1:1000 |
| Anti-GBA | Abcam, Cambridge, UK | Ab125065 | IgG rabbit monoclonal | 1:500 |
| Anti-GAPDH | Sigma-Aldrich, Darmstadt, Germany | SAB2108266 | IgG rabbit polyclonal | 1:18000 |
| Secondary antibodies | ||||
| Alexa Fluor 488 goat anti rabbit IgG (H + L) | Thermo Fisher Scientific, Waltham, MA, USA | A11008 | Goat | 1:400 |
| Alexa Fluor 568 goat anti rabbit IgG (H + L) | Thermo Fisher Scientific, Waltham, MA, USA | A11011 | Goat | 1:400 |
| Alexa Fluor 488 goat anti mouse IgG2a | Thermo Fisher Scientific, Waltham, MA, USA | A21131 | Goat | 1:400 |
| Goat Anti-Rabbit IgG H&L (HRP) | Abcam, Cambridge, UK | Ab6721 | Goat | 1:5000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kopytova, A.E.; Rychkov, G.N.; Cheblokov, A.A.; Grigor’eva, E.V.; Nikolaev, M.A.; Yarkova, E.S.; Sorogina, D.A.; Ibatullin, F.M.; Baydakova, G.V.; Izyumchenko, A.D.; et al. Potential Binding Sites of Pharmacological Chaperone NCGC00241607 on Mutant β-Glucocerebrosidase and Its Efficacy on Patient-Derived Cell Cultures in Gaucher and Parkinson’s Disease. Int. J. Mol. Sci. 2023, 24, 9105. https://doi.org/10.3390/ijms24109105
Kopytova AE, Rychkov GN, Cheblokov AA, Grigor’eva EV, Nikolaev MA, Yarkova ES, Sorogina DA, Ibatullin FM, Baydakova GV, Izyumchenko AD, et al. Potential Binding Sites of Pharmacological Chaperone NCGC00241607 on Mutant β-Glucocerebrosidase and Its Efficacy on Patient-Derived Cell Cultures in Gaucher and Parkinson’s Disease. International Journal of Molecular Sciences. 2023; 24(10):9105. https://doi.org/10.3390/ijms24109105
Chicago/Turabian StyleKopytova, Alena E., George N. Rychkov, Alexander A. Cheblokov, Elena V. Grigor’eva, Mikhail A. Nikolaev, Elena S. Yarkova, Diana A. Sorogina, Farid M. Ibatullin, Galina V. Baydakova, Artem D. Izyumchenko, and et al. 2023. "Potential Binding Sites of Pharmacological Chaperone NCGC00241607 on Mutant β-Glucocerebrosidase and Its Efficacy on Patient-Derived Cell Cultures in Gaucher and Parkinson’s Disease" International Journal of Molecular Sciences 24, no. 10: 9105. https://doi.org/10.3390/ijms24109105