
Flavone and Hydroxyflavones Are Ligands That Bind the Orphan Nuclear Receptor 4A1 (NR4A1)

 ,
,  , ,
, , 
Abstract
:1. Introduction
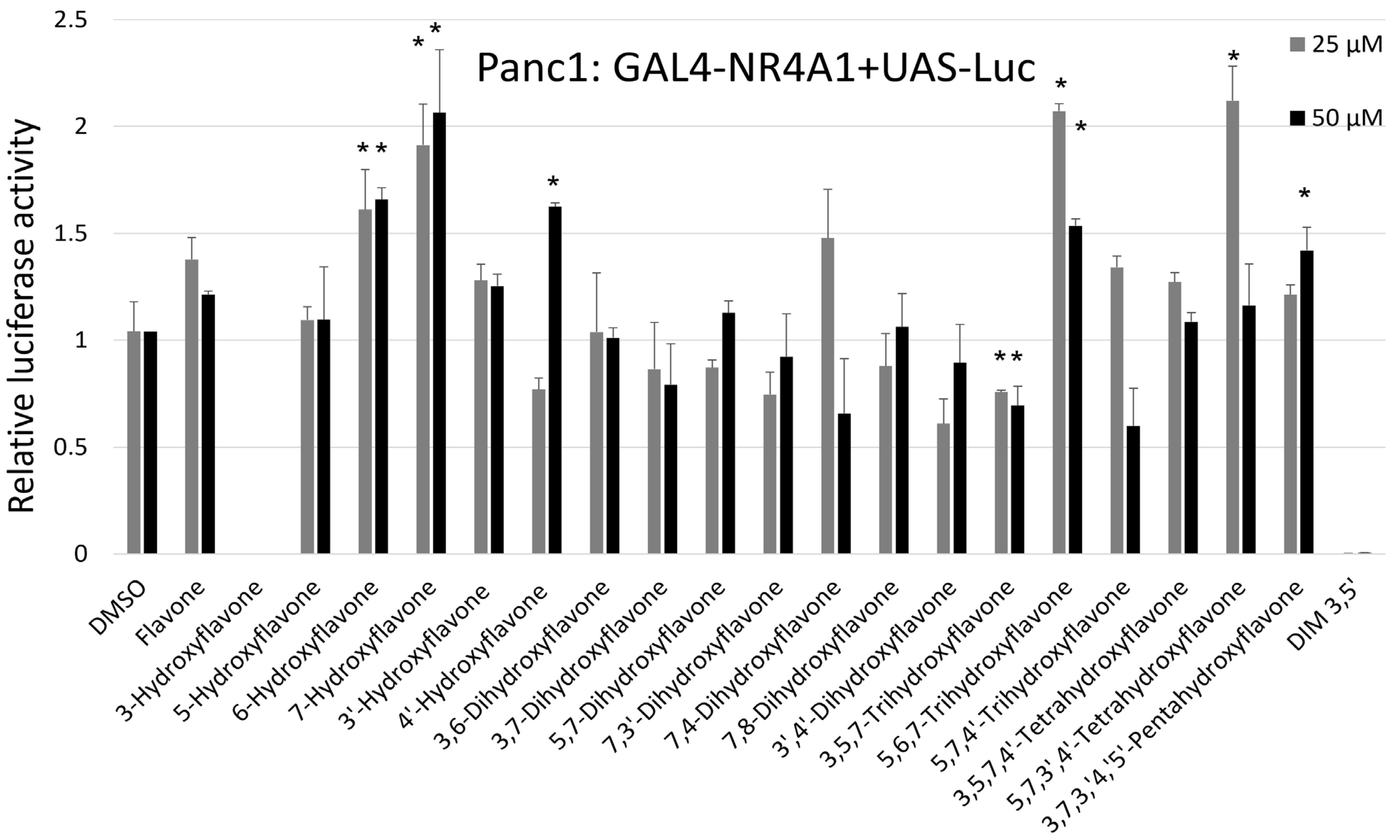
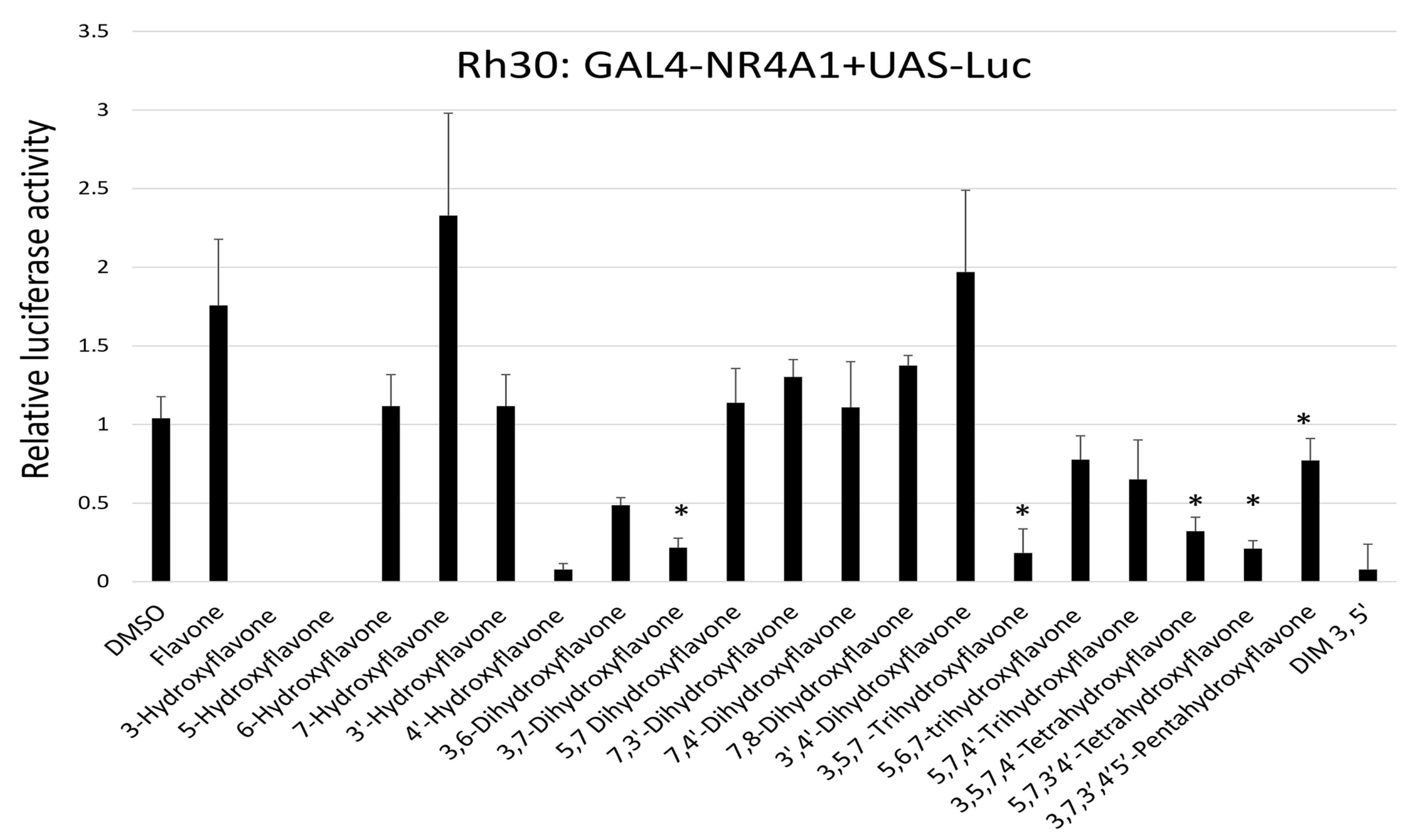
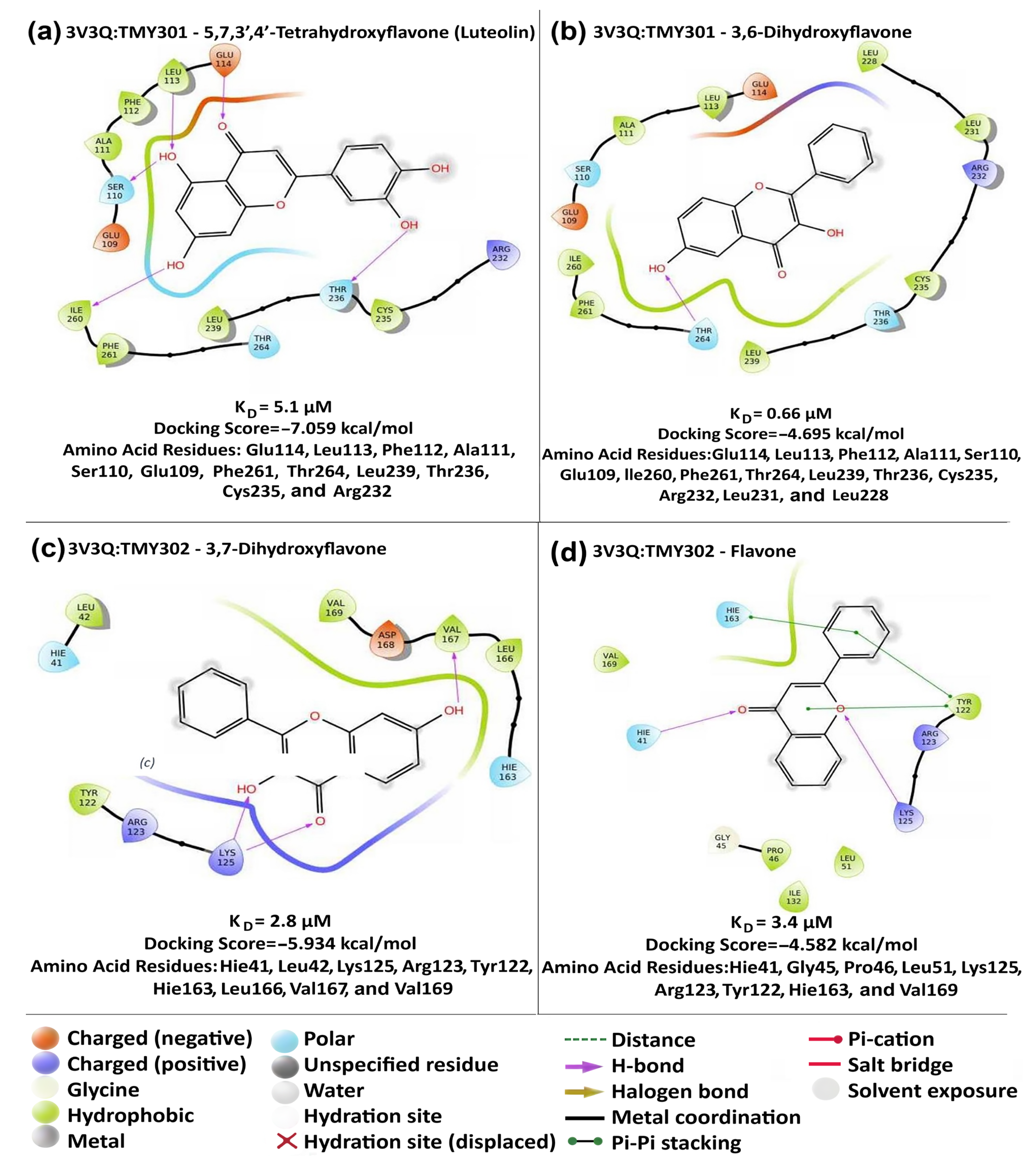
2. Results
3. Discussion
4. Materials and Methods
4.1. Chemicals and Cell Culture
4.2. Plasmids and Luciferase Assay
4.3. Quenching of NR4A1 Tryptophan Fluorescence by Direct Ligand Binding
4.4. Isothermal Titration Calorimetry
4.5. Computation-Based Molecular Modeling Studies
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef]
- Alseekh, S.; de Souza, L.P.; Benina, M.; Fernie, A.R. The style and substance of plant flavonoid decoration; towards defining both structure and function. Phytochemistry 2020, 174, 112347. [Google Scholar] [CrossRef] [PubMed]
- Hung, H.C.; Joshipura, K.J.; Jiang, R.; Hu, F.B.; Hunter, D.; Smith-Warner, S.A.; Colditz, G.A.; Rosner, B.; Spiegelman, D.; Willett, W.C. Fruit and vegetable intake and risk of major chronic disease. J. Natl. Cancer Inst. 2004, 96, 1577–1584. [Google Scholar] [CrossRef] [PubMed]
- Murillo, G.; Mehta, R.G. Cruciferous vegetables and cancer prevention. Nutr. Cancer 2001, 41, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Key, T.J. Fruit and vegetables and cancer risk. Br. J. Cancer 2010, 104, 6–11. [Google Scholar] [CrossRef]
- Rodríguez-García, C.; Sánchez-Quesada, C.; Gaforio, J.J. Dietary Flavonoids as Cancer Chemopreventive Agents: An Updated Review of Human Studies. Antioxidants 2019, 8, 137. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Weng, W.; Gao, R.; Liu, Y. New Insights for Cellular and Molecular Mechanisms of Aging and Aging-Related Diseases: Herbal Medicine as Potential Therapeutic Approach. Oxidative Med. Cell. Longev. 2019, 2019, 4598167. [Google Scholar] [CrossRef] [PubMed]
- Bondonno, N.P.; Dalgaard, F.; Kyrø, C.; Murray, K.; Bondonno, C.P.; Lewis, J.R.; Croft, K.D.; Gislason, G.; Scalbert, A.; Cassidy, A.; et al. Flavonoid intake is associated with lower mortality in the Danish Diet Cancer and Health Cohort. Nat. Commun. 2019, 10, 3651. [Google Scholar] [CrossRef] [PubMed]
- Shishtar, E.; Rogers, G.T.; Blumberg, J.B.; Au, R.; Jacques, P.F. Long-term dietary flavonoid intake and risk of Alzheimer disease and related dementias in the Framingham Offspring Cohort. Am. J. Clin. Nutr. 2020, 112, 343–353. [Google Scholar] [CrossRef]
- Marranzano, M.; Ray, S.; Godos, J.; Galvano, F. Association between dietary flavonoids intake and obesity in a cohort of adults living in the Mediterranean area. Int. J. Food Sci. Nutr. 2018, 69, 1020–1029. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Je, Y. Flavonoid intake and mortality from cardiovascular disease and all causes: A meta-analysis of prospective cohort studies. Clin. Nutr. ESPEN 2017, 20, 68–77. [Google Scholar] [CrossRef]
- Pounis, G.; Costanzo, S.; Bonaccio, M.; Di Castelnuovo, A.; de Curtis, A.; Ruggiero, E.; Persichillo, M.; Cerletti, C.; Donati, M.B.; de Gaetano, G.; et al. Reduced mortality risk by a polyphenol-rich diet: An analysis from the Moli-sani study. Nutrition 2018, 48, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Bi, X.; Yu, B.; Chen, D. Isoflavones: Anti-Inflammatory Benefit and Possible Caveats. Nutrients 2016, 8, 361. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, H.; Yuan, B.; Hu, X.; Okazaki, M. Chemopreventive and anticancer activity of flavonoids and its possibility for clinical use by combining with conventional chemotherapeutic agents. Am. J. Cancer Res. 2019, 9, 1517–1535. [Google Scholar]
- Sharma, S.; Naura, A.S. Potential of phytochemicals as immune-regulatory compounds in atopic diseases: A review. Biochem. Pharmacol. 2020, 173, 113790. [Google Scholar] [CrossRef]
- Oteiza, P.; Fraga, C.; Mills, D.; Taft, D. Flavonoids and the gastrointestinal tract: Local and systemic effects. Mol. Asp. Med. 2018, 61, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Salaritabar, A.; Darvishi, B.; Hadjiakhoondi, F.; Manayi, A.; Sureda, A.; Nabavi, S.F.; Fitzpatrick, L.R.; Bishayee, A. Therapeutic potential of flavonoids in inflammatory bowel disease: A comprehensive review. World J. Gastroenterol. 2017, 23, 5097–5114. [Google Scholar] [CrossRef]
- Algieri, F.; Rodriguez-Nogales, A.; Rodriguez-Cabezas, M.E.; Risco, S.; Ocete, M.A.; Galvez, J. Botanical Drugs as an Emerging Strategy in Inflammatory Bowel Disease: A Review. Mediat. Inflamm. 2015, 2015, 179616. [Google Scholar] [CrossRef]
- Park, S.; Song, G.; Lim, W. Myricetin inhibits endometriosis growth through cyclin E1 down-regulation in vitro and in vivo. J. Nutr. Biochem. 2020, 78, 108328. [Google Scholar] [CrossRef]
- Park, S.; Lim, W.; Bazer, F.W.; Whang, K.-Y.; Song, G. Quercetin inhibits proliferation of endometriosis regulating cyclin D1 and its target microRNAs in vitro and in vivo. J. Nutr. Biochem. 2018, 63, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.; Xu, H.; Man, G.C.; Zhang, T.; Chu, K.O.; Chu, C.Y.; Cheng, J.T.; Li, G.; He, Y.X.; Qin, L.; et al. Prodrug of green tea epigallocatechin-3-gallate (Pro-EGCG) as a potent anti-angiogenesis agent for endometriosis in mice. Angiogenesis 2013, 16, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed]
- Safe, S.; Jayaraman, A.; Chapkin, R.S.; Howard, M.; Mohankumar, K.; Shrestha, R. Flavonoids: Structure–function and mechanisms of action and opportunities for drug development. Toxicol. Res. 2021, 37, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Goya-Jorge, E.; Rodríguez, M.E.J.; Veitía, M.S.-I.; Giner, R.M. Plant Occurring Flavonoids as Modulators of the Aryl Hydrocarbon Receptor. Molecules 2021, 26, 2315. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Lv, Q.; Miao, Y.M.; Qiao, S.M.; Dai, Y.; Wei, Z.F. Cardamonin, a natural flavone, alleviates inflammatory bowel disease by the inhibition of NLRP3 inflammasome activation via an AhR/Nrf2/NQO1 pathway. Biochem. Pharmacol. 2018, 155, 494–509. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.-J.; Shin, H.S.; See, H.-J.; Jung, S.Y.; Kwon, D.-A.; Shon, D.-H. Baicalein induces CD4+Foxp3+ T cells and enhances intestinal barrier function in a mouse model of food allergy. Sci. Rep. 2016, 6, 32225. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, J.; Xie, Y. Improvement strategies for the oral bioavailability of poorly water-soluble flavonoids: An overview. Int. J. Pharm. 2019, 570, 118642. [Google Scholar] [CrossRef]
- Lith, S.C.; van Os, B.W.; Seijkens, T.T.P.; de Vries, C.J.M. ‘Nur’turing tumor T cell tolerance and exhaustion: Novel function for Nuclear Receptor Nur77 in immunity. Eur. J. Immunol. 2020, 50, 1643–1652. [Google Scholar] [CrossRef]
- Pearen, M.A.; Muscat, G.E.O. Minireview: Nuclear Hormone Receptor 4A Signaling: Implications for Metabolic Disease. Mol. Endocrinol. 2010, 24, 1891–1903. [Google Scholar] [CrossRef]
- Safe, S.; Karki, K. The Paradoxical Roles of Orphan Nuclear Receptor 4A (NR4A) in Cancer. Mol. Cancer Res. 2021, 19, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Karki, K.; Mohankumar, K.; Schoeller, A.; Martin, G.; Shrestha, R.; Safe, S. NR4A1 Ligands as Potent Inhibitors of Breast Cancer Cell and Tumor Growth. Cancers 2021, 13, 2682. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, R.; Mohankumar, K.; Martin, G.; Hailemariam, A.; Lee, S.-O.; Jin, U.-H.; Burghardt, R.; Safe, S. Flavonoids kaempferol and quercetin are nuclear receptor 4A1 (NR4A1, Nur77) ligands and inhibit rhabdomyosarcoma cell and tumor growth. J. Exp. Clin. Cancer Res. 2021, 40, 392. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Martin, G.; Mohankumar, K.; Hampton, J.T.; Liu, W.R.; Safe, S. Resveratrol Binds Nuclear Receptor 4A1 (NR4A1) and Acts as an NR4A1 Antagonist in Lung Cancer Cells. Mol. Pharmacol. 2022, 102, 80–91. [Google Scholar] [CrossRef]
- Lee, H.-S.; Kim, S.-H.; Kim, B.-M.; Safe, S.; Lee, S.-O. Broussochalcone A Is a Novel Inhibitor of the Orphan Nuclear Receptor NR4A1 and Induces Apoptosis in Pancreatic Cancer Cells. Molecules 2021, 26, 2316. [Google Scholar] [CrossRef]
- Park, H.; Jin, U.-H.; Martin, G.; Chapkin, R.S.; Davidson, L.A.; Lee, K.; Jayaraman, A.; Safe, S. Structure-activity relationships among mono- and dihydroxy flavones as aryl hydrocarbon receptor (AhR) agonists or antagonists in CACO2 cells. Chem. Interactions 2022, 365, 110067. [Google Scholar] [CrossRef]
- Jin, U.-H.; Park, H.; Li, X.; Davidson, L.A.; Allred, C.; Patil, B.; Jayaprakasha, G.; Orr, A.A.; Mao, L.; Chapkin, R.S.; et al. Structure-Dependent Modulation of Aryl Hydrocarbon Receptor-Mediated Activities by Flavonoids. Toxicol. Sci. 2018, 164, 205–217. [Google Scholar] [CrossRef]
- Zhan, Y.-Y.; Chen, Y.; Zhang, Q.; Zhuang, J.-J.; Tian, M.; Chen, H.-Z.; Zhang, L.-R.; Zhang, H.-K.; He, J.-P.; Wang, W.-J.; et al. The orphan nuclear receptor Nur77 regulates LKB1 localization and activates AMPK. Nat. Chem. Biol. 2012, 8, 897–904. [Google Scholar] [CrossRef]
- Jordan, V.C. Antiestrogens and Selective Estrogen Receptor Modulators as Multifunctional Medicines. 1. Receptor Interactions. J. Med. Chem. 2003, 46, 883–908. [Google Scholar] [CrossRef]
- Burris, T.P.; Solt, L.A.; Wang, Y.; Crumbley, C.; Banerjee, S.; Griffett, K.; Lundasen, T.; Hughes, T.; Kojetin, D.J. Nuclear Receptors and Their Selective Pharmacologic Modulators. Pharmacol. Rev. 2013, 65, 710–778. [Google Scholar] [CrossRef]
- Mullican, S.E.; Zhang, S.; Konopleva, M.; Ruvolo, V.; Andreeff, M.; Milbrandt, J.; Conneely, O.M. Abrogation of nuclear receptors Nr4a3 andNr4a1 leads to development of acute myeloid leukemia. Nat. Med. 2007, 13, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Herrick, A.M.; Mullican, S.E.; Sheehan, A.M.; Conneely, O.M. Reduced NR4A gene dosage leads to mixed myelodysplastic/myeloproliferative neoplasms in mice. Blood 2011, 117, 2681–2690. [Google Scholar] [CrossRef]
- Liu, J.-J.; Zeng, H.-N.; Zhang, L.-R.; Zhan, Y.-Y.; Chen, Y.; Wang, Y.; Wang, J.; Xiang, S.-H.; Liu, W.-J.; Wang, W.-J.; et al. A Unique Pharmacophore for Activation of the Nuclear Orphan Receptor Nur77 In vivo and In vitro. Cancer Res. 2010, 70, 3628–3637. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, D.; Yan, S.; Hu, C.; Huang, Z.; Li, Z.; Peng, S.; Li, X.; Zhu, Y.; Yu, H.; et al. SAR study of celastrol analogs targeting Nur77-mediated inflammatory pathway. Eur. J. Med. Chem. 2019, 177, 171–187. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.S.; Lee, H.S.; Cho, H.R.; Kim, K.J.; Kim, J.H.; Safe, S.; Lee, S.O. Dual targeting of Nur77 and AMPKalpha by isoalantolactone inhibits adipogenesis in vitro and decreases body fat mass in vivo. Int. J. Obes. 2019, 43, 952–962. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Safe, S.; Lee, S.-O. Inactivation of the orphan nuclear receptor NR4A1 contributes to apoptosis induction by fangchinoline in pancreatic cancer cells. Toxicol. Appl. Pharmacol. 2017, 332, 32–39. [Google Scholar] [CrossRef]
- Hu, M.; Luo, Q.; Alitongbieke, G.; Chong, S.; Xu, C.; Xie, L.; Chen, X.; Zhang, D.; Zhou, Y.; Wang, Z.; et al. Celastrol-Induced Nur77 Interaction with TRAF2 Alleviates Inflammation by Promoting Mitochondrial Ubiquitination and Autophagy. Mol. Cell 2017, 66, 141–153.e6. [Google Scholar] [CrossRef]
- Lakshmi, S.P.; Reddy, A.T.; Banno, A.; Reddy, R.C. Molecular, chemical, and structural characterization of prostaglandin A2 as a novel agonist for Nur77. Biochem. J. 2019, 476, 2757–2767. [Google Scholar] [CrossRef]
- Zhou, M.; Peng, B.-R.; Tian, W.; Su, J.-H.; Wang, G.; Lin, T.; Zeng, D.; Sheu, J.-H.; Chen, H. 12-Deacetyl-12-epi-Scalaradial, a Scalarane Sesterterpenoid from a Marine Sponge Hippospongia sp., Induces HeLa Cells Apoptosis via MAPK/ERK Pathway and Modulates Nuclear Receptor Nur77. Mar. Drugs 2020, 18, 375. [Google Scholar] [CrossRef]
- Li, W.; Hang, S.; Fang, Y.; Bae, S.; Zhang, Y.; Zhang, M.; Wang, G.; McCurry, M.D.; Bae, M.; Paik, D.; et al. A bacterial bile acid metabolite modulates Treg activity through the nuclear hormone receptor NR4A1. Cell Host. Microbe 2021, 29, 1366–1377.e9. [Google Scholar] [CrossRef]
- Vinayavekhin, N.; Saghatelian, A. Discovery of a Protein–Metabolite Interaction between Unsaturated Fatty Acids and the Nuclear Receptor Nur77 Using a Metabolomics Approach. J. Am. Chem. Soc. 2011, 133, 17168–17171. [Google Scholar] [CrossRef]
- Rajan, S.; Jang, Y.; Kim, C.-H.; Kim, W.; Toh, H.T.; Jeon, J.; Song, B.; Serra, A.; Lescar, J.; Yoo, J.Y.; et al. PGE1 and PGA1 bind to Nurr1 and activate its transcriptional function. Nat. Chem. Biol. 2020, 16, 876–886. [Google Scholar] [CrossRef]
- Bruning, J.M.; Wang, Y.; Oltrabella, F.; Tian, B.; Kholodar, S.A.; Liu, H.; Bhattacharya, P.; Guo, S.; Holton, J.M.; Fletterick, R.J.; et al. Covalent Modification and Regulation of the Nuclear Receptor Nurr1 by a Dopamine Metabolite. Cell Chem. Biol. 2019, 26, 674–685.e6. [Google Scholar] [CrossRef] [PubMed]
- de Vera, I.M.; Giri, P.K.; Munoz-Tello, P.; Brust, R.; Fuhrmann, J.; Matta-Camacho, E.; Shang, J.; Campbell, S.; Wilson, H.D.; Granados, J.; et al. Identification of a Binding Site for Unsaturated Fatty Acids in the Orphan Nuclear Receptor Nurr1. ACS Chem. Biol. 2016, 11, 1795–1799. [Google Scholar] [CrossRef] [PubMed]
- Kholodar, S.A.; Lang, G.; Cortopassi, W.A.; Iizuka, Y.; Brah, H.S.; Jacobson, M.P.; England, P.M. Analogs of the Dopamine Metabolite 5,6-Dihydroxyindole Bind Directly to and Activate the Nuclear Receptor Nurr1. ACS Chem. Biol. 2021, 16, 1159–1163. [Google Scholar] [CrossRef]
- Ponte, L.G.S.; Pavan, I.C.B.; Mancini, M.C.S.; Da Silva, L.G.S.; Morelli, A.P.; Severino, M.B.; Bezerra, R.M.N.; Simabuco, F.M. The Hallmarks of Flavonoids in Cancer. Molecules 2021, 26, 2029. [Google Scholar] [CrossRef]
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2018, 11, 28. [Google Scholar] [CrossRef]
- Xie, Y.; Tian, Y.; Zhang, Y.; Zhang, Z.; Chen, R.; Li, M.; Tang, J.; Bian, J.; Li, Z.; Xu, X. Overview of the development of selective androgen receptor modulators (SARMs) as pharmacological treatment for osteoporosis (1998–2021). Eur. J. Med. Chem. 2022, 230, 114119. [Google Scholar] [CrossRef] [PubMed]
- D’arrigo, G.; Gianquinto, E.; Rossetti, G.; Cruciani, G.; Lorenzetti, S.; Spyrakis, F. Binding of Androgen- and Estrogen-Like Flavonoids to Their Cognate (Non)Nuclear Receptors: A Comparison by Computational Prediction. Molecules 2021, 26, 1613. [Google Scholar] [CrossRef]
- Choi, S.Y.; Ha, T.Y.; Ahn, J.Y.; Kim, S.R.; Kang, K.S.; Hwang, I.K.; Kim, S. Estrogenic activities of isoflavones and flavones and their structure-activity relationships. Planta Med. 2008, 74, 25–32. [Google Scholar] [CrossRef]
- Halgren, T.A.; Murphy, R.B.; Friesner, R.A.; Beard, H.S.; Frye, L.L.; Pollard, W.T.; Banks, J.L. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 2. Enrichment Factors in Database Screening. J. Med. Chem. 2004, 47, 1750–1759. [Google Scholar] [CrossRef] [PubMed]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra precision glide: Docking and scoring incorporating a model of hydrophobic enclosure for protein-ligand complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [PubMed]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
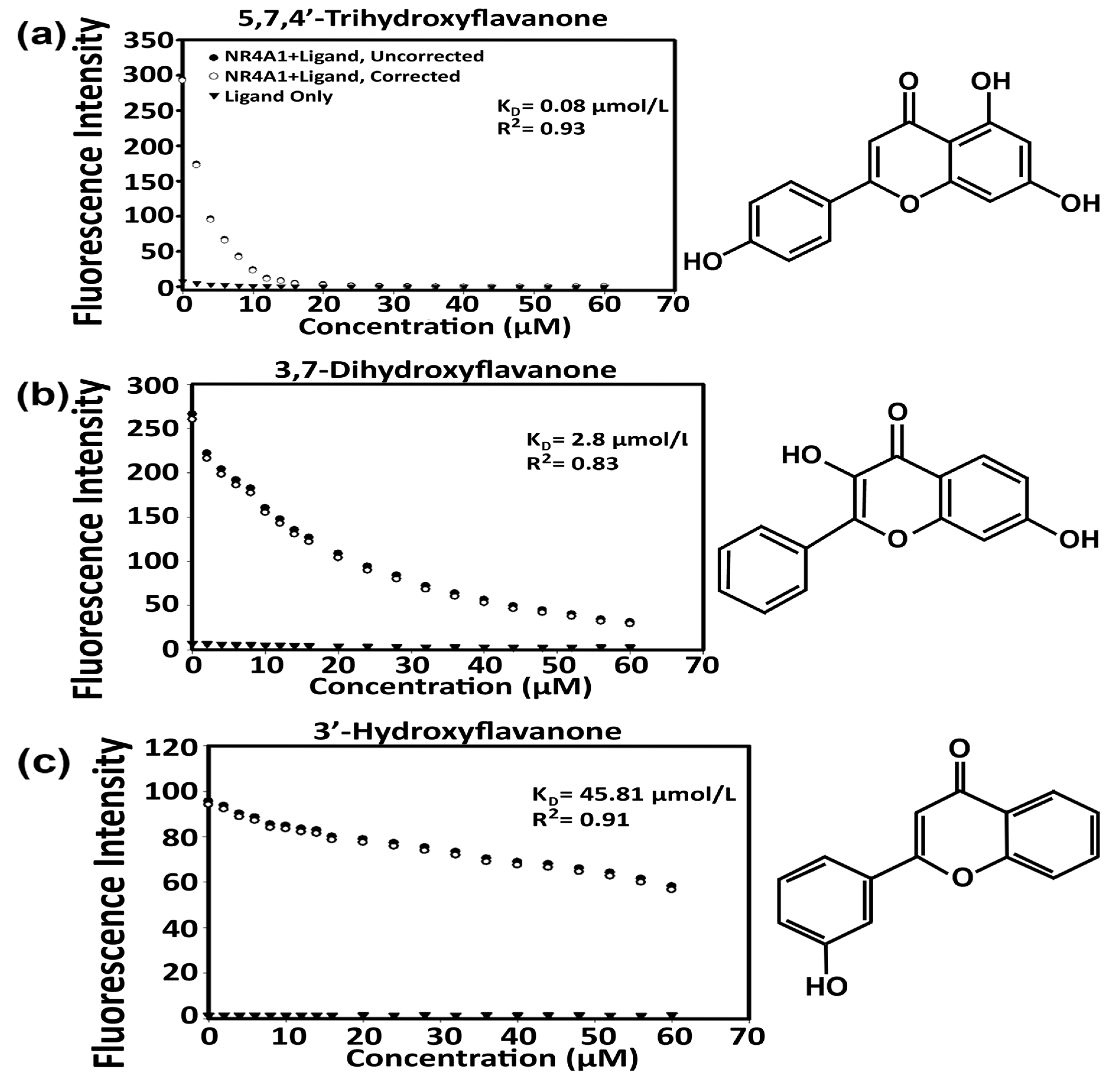
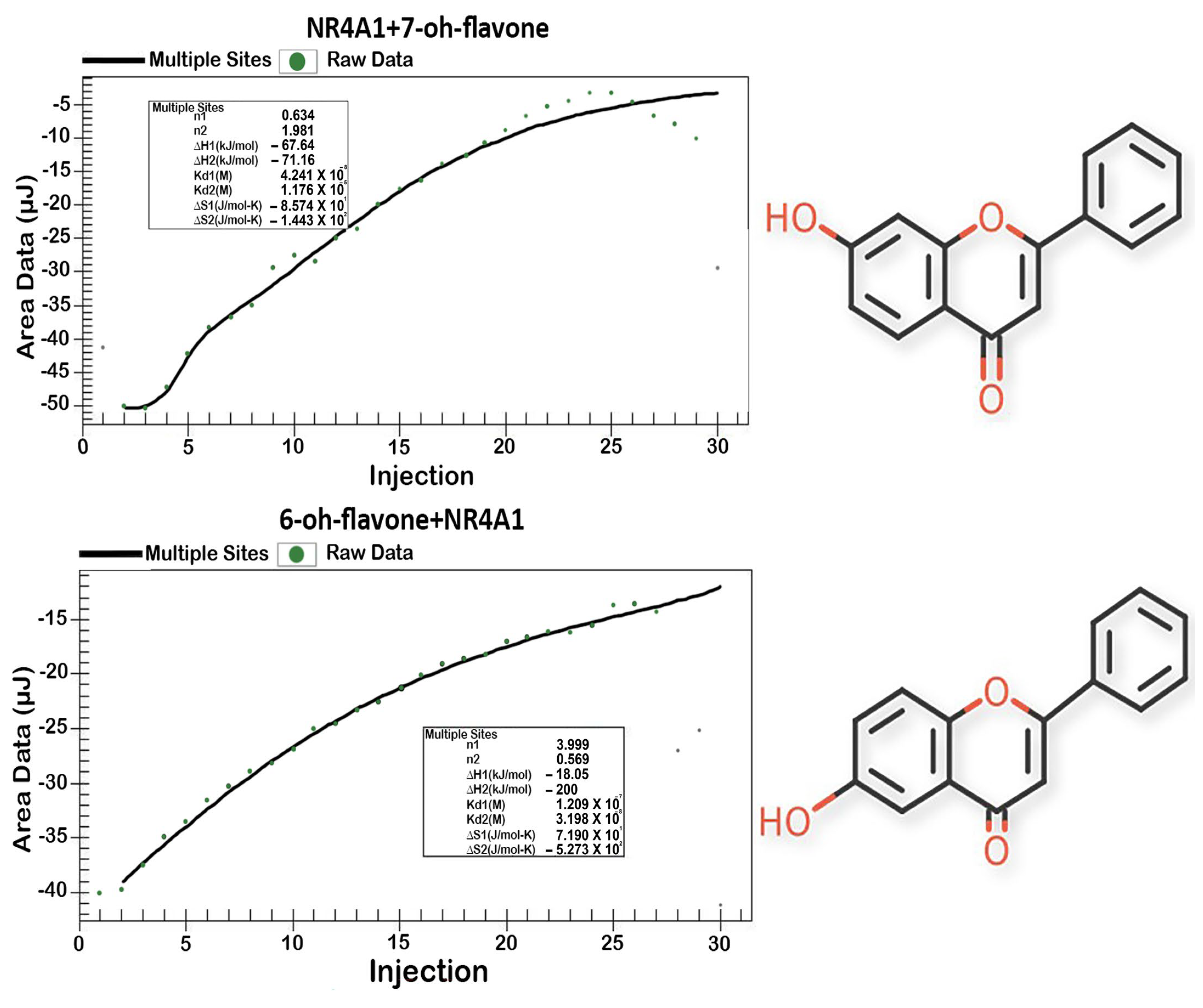
| Compound | Direct Binding Assay | ITC Assay | ||
|---|---|---|---|---|
| KD | R2 | Kd1, mmol/L | ΔG, kJ/mol | |
| Flavone | 3.4 | 0.9 | 0.075 | −40.7 |
| 3-Hydroxyflavone | N/A * | N/A | 0.0225 | −43.6 |
| 5-Hydroxyflavone | 1.4 | 0.87 | 18 | −27 |
| 6-Hydroxyflavone | 12.5 | 0.86 | 0.032 | −42.9 |
| 7-Hydroxyflavone | 7.4 | 0.93 | 0.042 | −42.1 |
| 3′-Hydroxyflavone | 45.8 | 0.91 | 0.33 | −37 |
| 4′-Hydroxyflavone | 13.02 | 0.9 | 0.15 | −39.4 |
| 3,6-Dihydroxyflavone | 0.66 | 0.98 | 444 | −19.1 |
| 3,7-Dihydroxyflavone | 2.8 | 0.83 | 0.18 | −38.5 |
| 5,7 Dihydroxyflavone | 11.3 | 0.89 | 0.001 | −51.3 |
| 7,3′-Dihydroxyflavone | 21.4 | 0.89 | 0.17 | −38.7 |
| 7,4’-Dihydroxyflavone | 9.1 | 0.68 | 0.49 | −36.1 |
| 7,8-Dihydroxyflavone | 17.64 | 0.99 | 0.042 | −42.1 |
| 3′,4′-Dihydroxyflavone | 0.96 | 0.59 | 9.5 | −28.7 |
| 3,5,7 -Trihydroxyflavone (Galangin) | 0.36 | 0.64 | 0.001 | −51.2 |
| 5,6,7 -Trihydroxyflavone (Baicalein) | 1.85 | 0.91 | 0.13 | −39.4 |
| 5,7,4′-Trihydroxyflavone (Apigenin) | 1.77 | 0.87 | 0.57 | −35.7 |
| 3,5,7,4′-Tetrahydroxyflavone (Kaempferol) | 3.1 | 0.95 | 97.1 | −22.9 |
| 5,7,3′4′-Tetrahydroxyflavone (Luteolin) | 5.1 | 0.98 | 0.019 | −44.1 |
| 3,7,3′,4′5′-Pentahydroxyflavone (Quercetin) | 0.81 | 0.97 | 0.35 | −36.8 |
| Compound | Site TMY301: Docking Score kcal/mol | Site TMY302: Docking Score kcal/mol |
|---|---|---|
| Flavone | −5.741 | −4.582 |
| 3-Hydroxyflavone | −5.507 | −4.716 |
| 5-Hydroxyflavone | −6.302 | −4.732 |
| 6-Hydroxyflavone | −5.619 | −5.4 |
| 7-Hydroxyflavone | −5.878 | −5.477 |
| 3′-Hydroxyflavone | −6.756 | −5.176 |
| 4′-Hydroxyflavone | −5.861 | −4.717 |
| 3,6-Dihydroxyflavone | −4.695 | −5.51 |
| 3,7-Dihydroxyflavone | −5.934 | −5.851 |
| 5,7 Dihydroxyflavone | −6.399 | −5.543 |
| 7,3′-Dihydroxyflavone | −6.719 | −5.559 |
| 7,4′-Dihydroxyflavone | −5.575 | −5.42 |
| 7,8-Dihydroxyflavone | −5.941 | −5.049 |
| 3′,4′-Dihydroxyflavone | −5.736 | −5.507 |
| 3,5,7-Trihydroxyflavone (Galangin) | −6.791 | −4.83 |
| 5,6,7-Trihydroxyflavone (Baicalein) | −5.711 | −5.711 |
| 5,7,4′-Trihydroxyflavone (Apigenin) | −6.441 | −4.782 |
| 3,5,7,4′-Tetrahydroxyflavone (Kaempferol) | −6.555 | −5.433 |
| 5,7,3′4′-Tetrahydroxyflavone (Luteolin) | −7.059 | −5.832 |
| 3,7,3′,4′5′-Pentahydroxyflavone (Quercetin) | −6.437 | −5.397 |
| 4′,5,7-Trihydroxyflavone (Naringenin) | −6.618 | −5.656 |
| Binding Pocket Residues | GLU114, LEU113, PHE112, ALA111, SER110, GLU109, LEU108, ILE260, PHE261, THR264, PRO266, LEU239, THR236, CYS235, ARG232, ARG184, VAL179 | HIE41, LEU42, ASP43, SER44, GLY45, PRO46, SER47, THR48, LEU51, ILE132, LYS125, ARG123, TYR122, ARG119, LEU162, HIE163, LEU165, LEU166, VAL167, VAL169, PHE172 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.; Upadhyay, S.; Mariyam, F.; Martin, G.; Hailemariam, A.; Lee, K.; Jayaraman, A.; Chapkin, R.S.; Lee, S.-O.; Safe, S. Flavone and Hydroxyflavones Are Ligands That Bind the Orphan Nuclear Receptor 4A1 (NR4A1). Int. J. Mol. Sci. 2023, 24, 8152. https://doi.org/10.3390/ijms24098152
Lee M, Upadhyay S, Mariyam F, Martin G, Hailemariam A, Lee K, Jayaraman A, Chapkin RS, Lee S-O, Safe S. Flavone and Hydroxyflavones Are Ligands That Bind the Orphan Nuclear Receptor 4A1 (NR4A1). International Journal of Molecular Sciences. 2023; 24(9):8152. https://doi.org/10.3390/ijms24098152
Chicago/Turabian StyleLee, Miok, Srijana Upadhyay, Fuada Mariyam, Greg Martin, Amanuel Hailemariam, Kyongbum Lee, Arul Jayaraman, Robert S. Chapkin, Syng-Ook Lee, and Stephen Safe. 2023. "Flavone and Hydroxyflavones Are Ligands That Bind the Orphan Nuclear Receptor 4A1 (NR4A1)" International Journal of Molecular Sciences 24, no. 9: 8152. https://doi.org/10.3390/ijms24098152