
MicroRNA-146b-5p Suppresses Pro-Inflammatory Mediator Synthesis via Targeting TRAF6, IRAK1, and RELA in Lipopolysaccharide-Stimulated Human Dental Pulp Cells

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
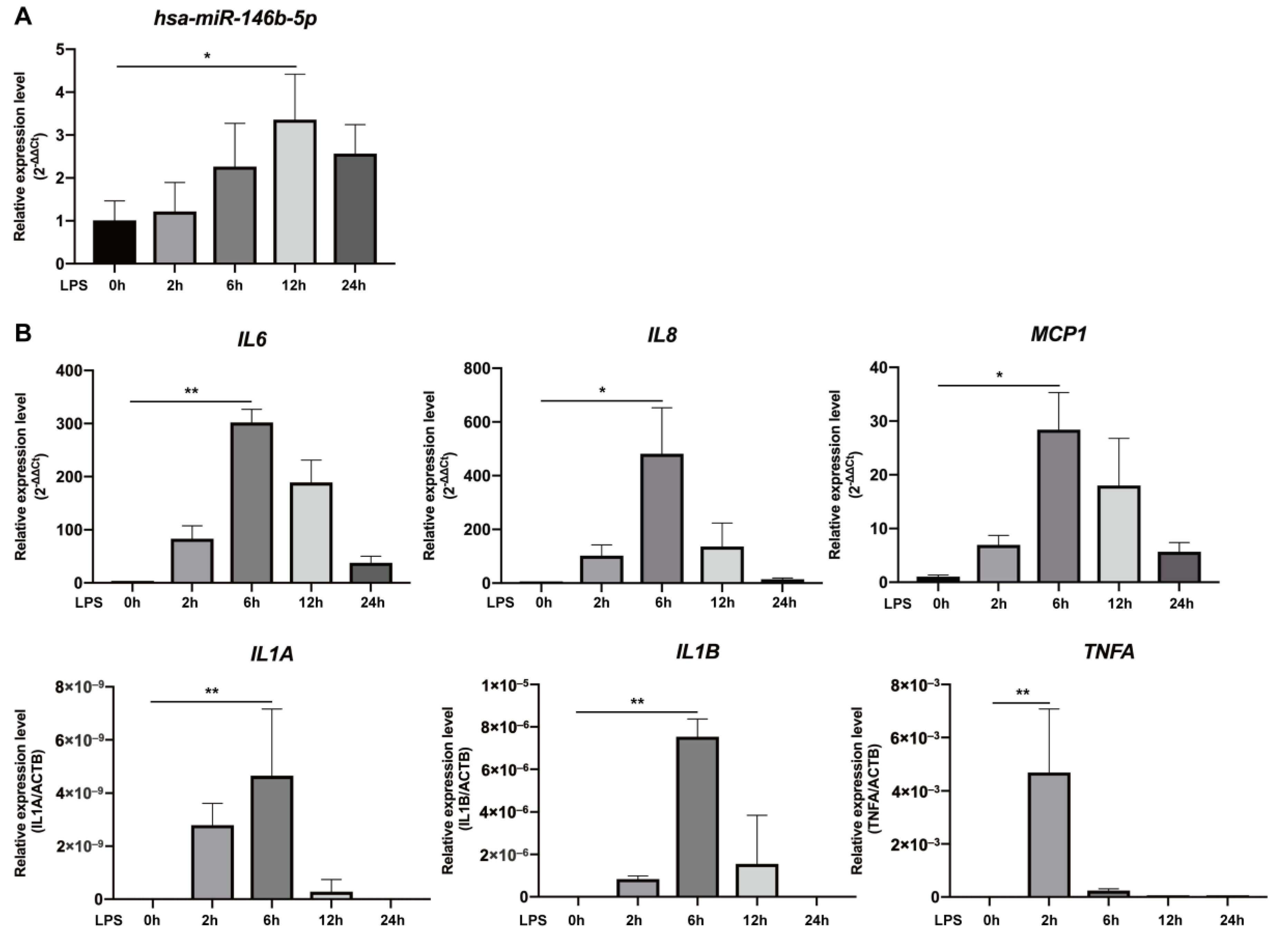
2.1. Expression of miR-146b-5p and mRNA of Pro-Inflammatory Mediators Was Up-Regulated in LPS-Stimulated hDPCs
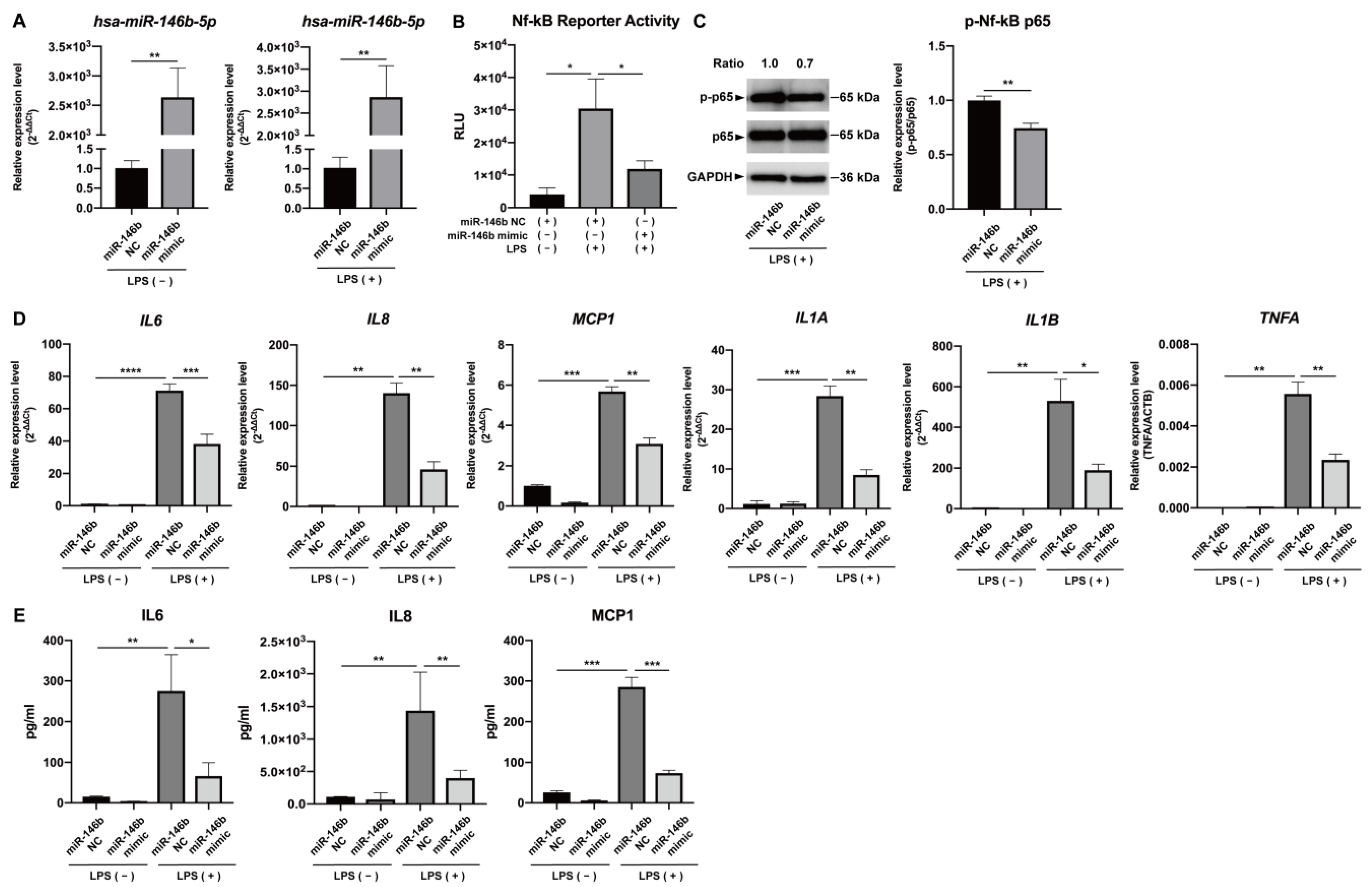
2.2. NF-κB and JAK/STAT3 Signaling Were Involved in the Expression of Pro-Inflammatory Cytokines and miR-146b-5p in LPS-Stimulated hDPCs, Respectively
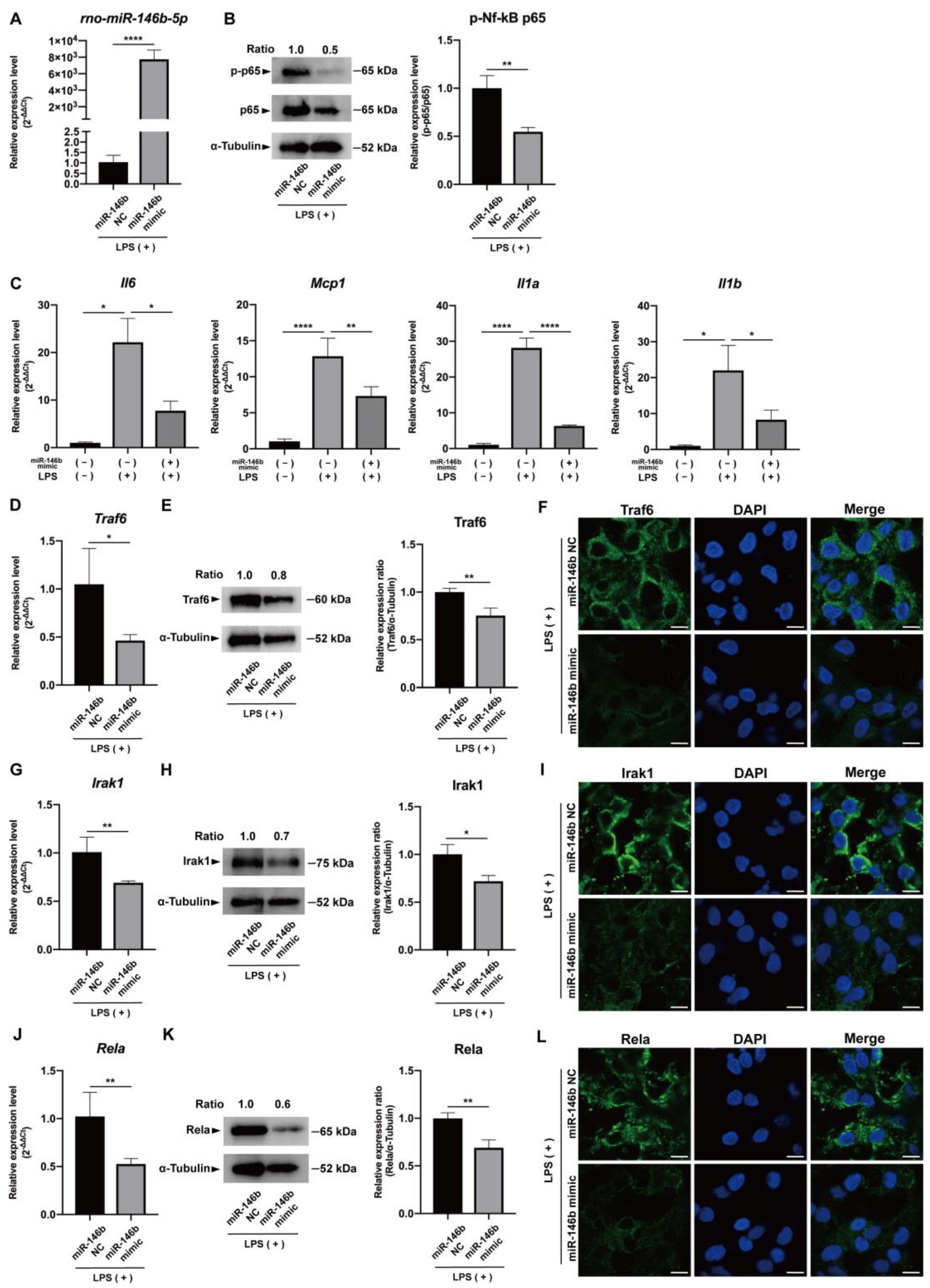
2.3. miR-146b-5p Down-Regulated NF-κB Signaling and Synthesis of Pro-Inflammatory Cytokines in LPS-Stimulated hDPCs
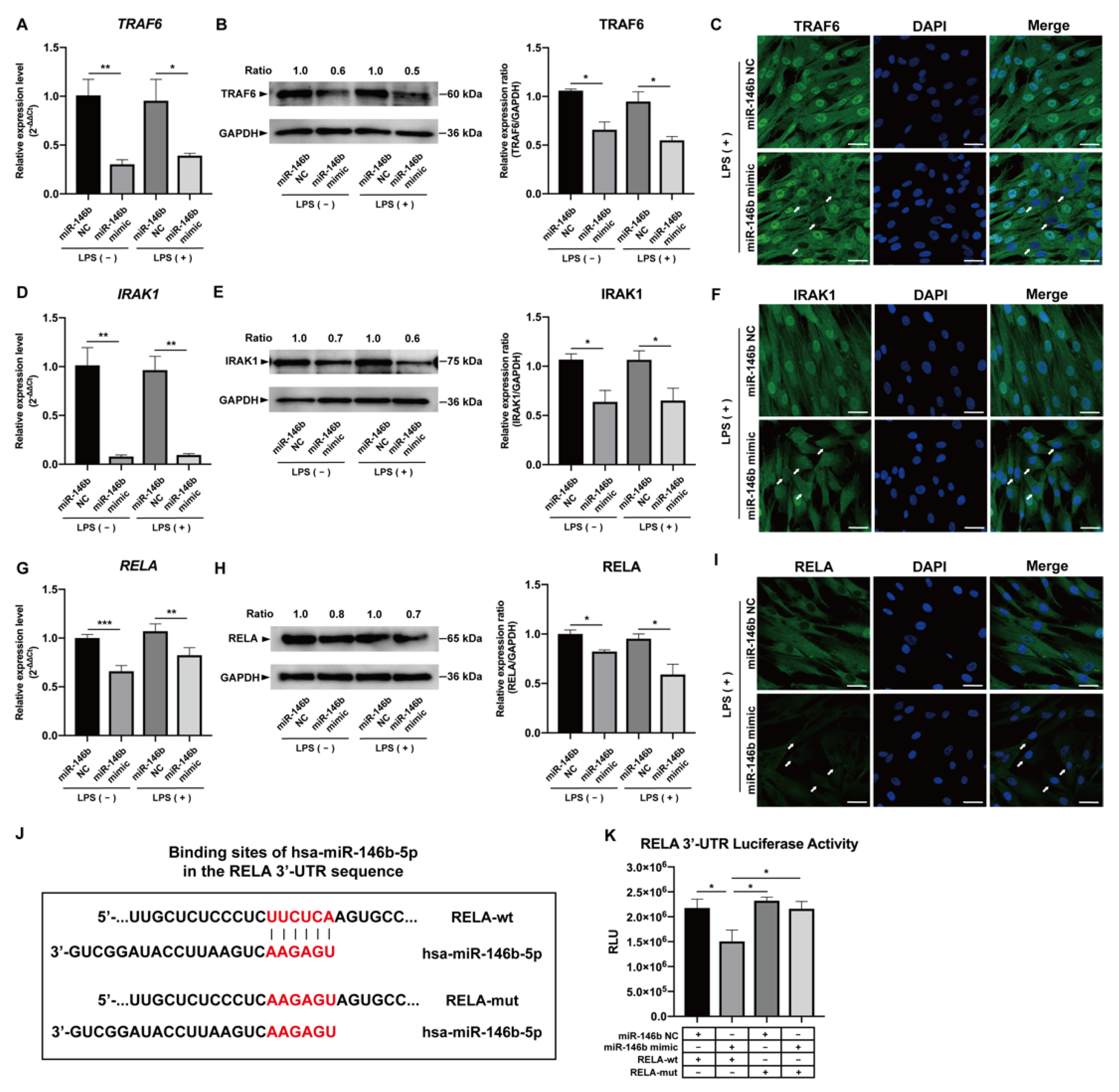
2.4. miR-146b-5p Targeted TRAF6, IRAK1, and RELA
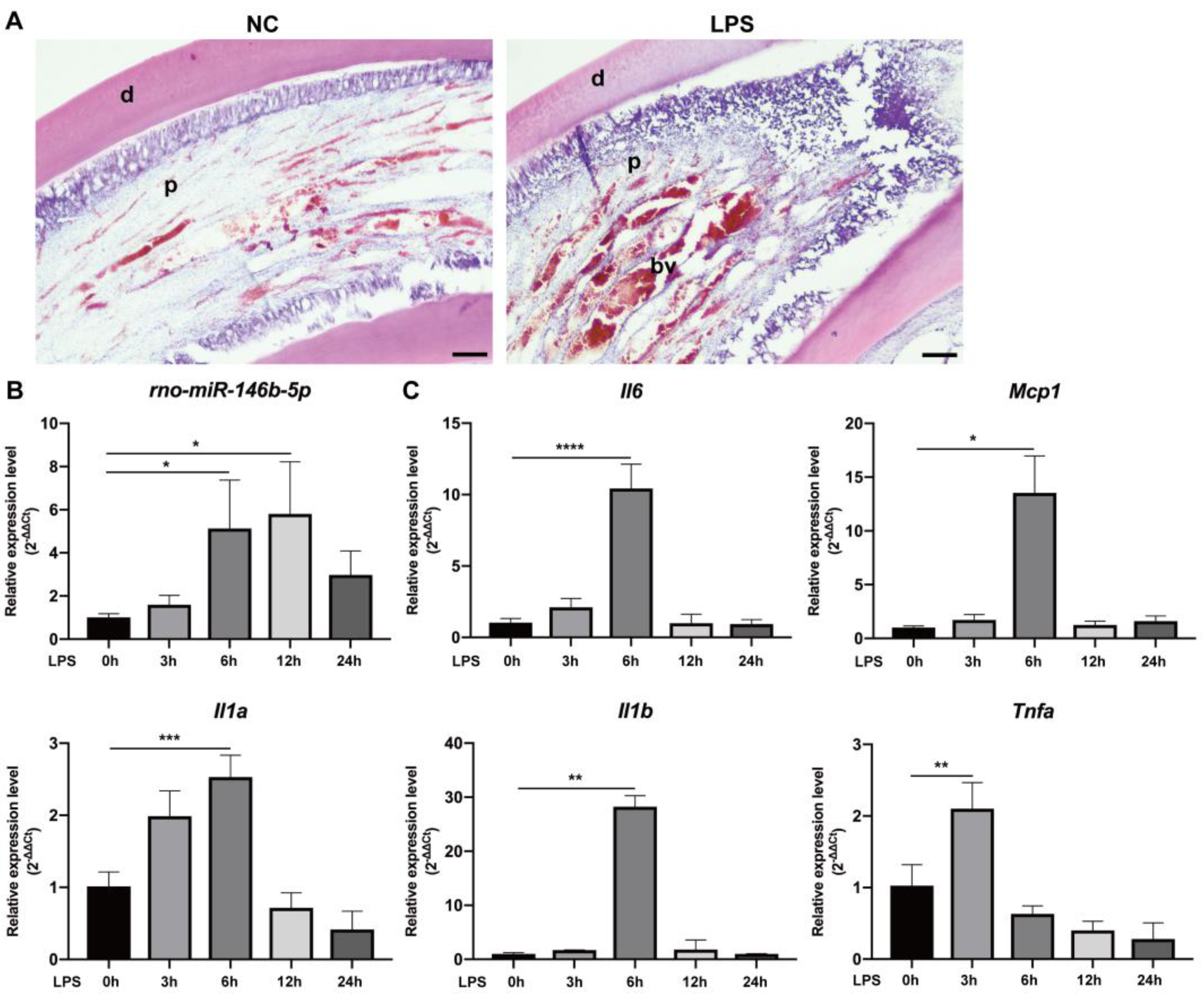
2.5. miR-146b-5p Down-Regulated Pro-Inflammatory Cytokines in Rat Dental Pulp Tissue via Targeting TRAF6, IRAK1, and RELA Ex Vivo
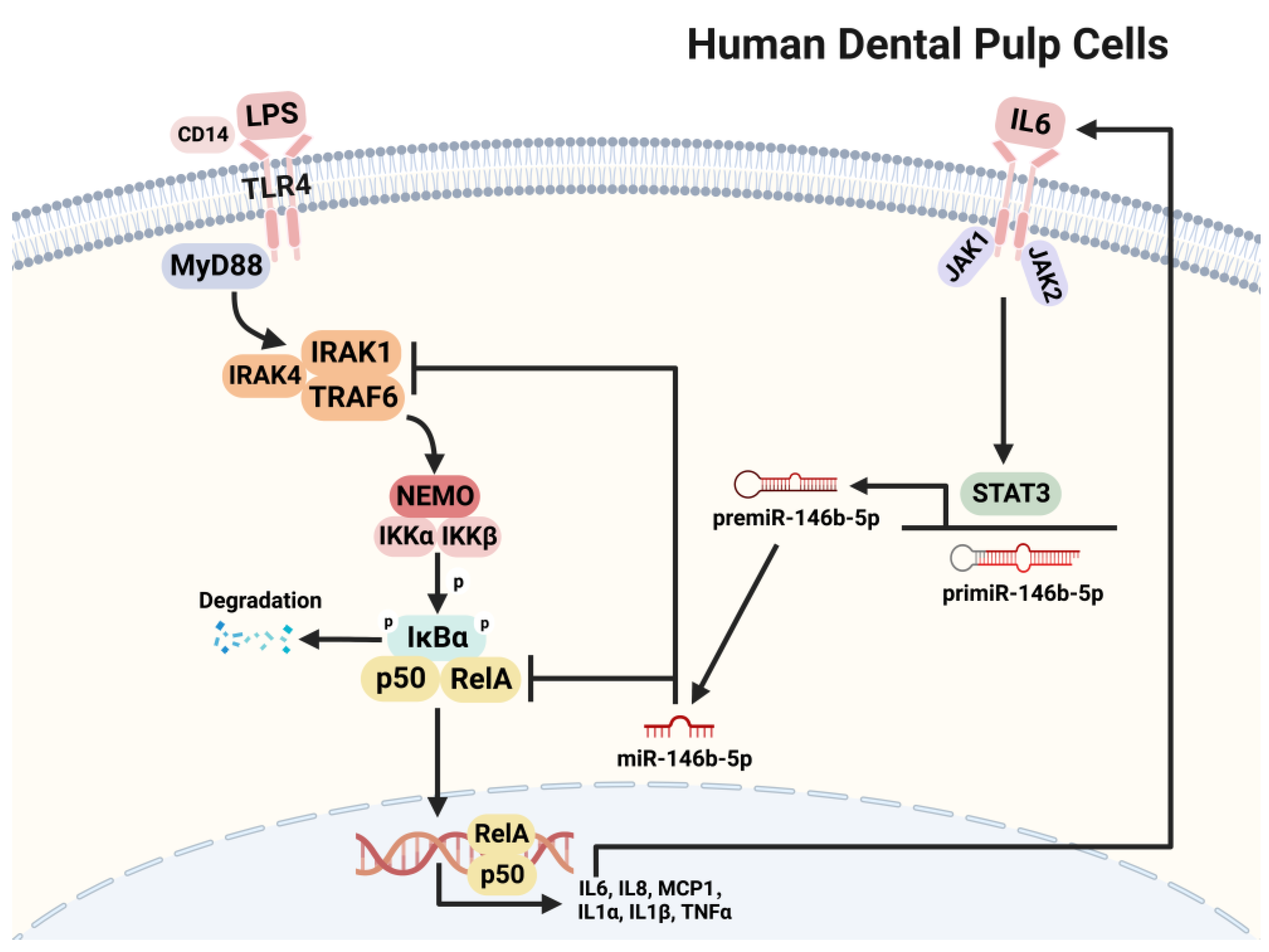
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. miR-146b-5p Mimic Transfection
4.3. LPS-Induced Rat Pulp Inflammation In Vivo
4.4. Ex Vivo miR-146b-5p Mimic Transfection
4.5. RT-qPCR
4.6. Western Blotting
4.7. Cytometric Bead Array (CBA)
4.8. Luciferase Assay with NF-κB and RELA 3′-UTR Reporter Vectors
4.9. Immunofluorescence
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, C.; Abbott, P.V. An overview of the dental pulp: Its functions and responses to injury. Aust. Dent. J. 2007, 52, S4–S16. [Google Scholar] [CrossRef] [PubMed]
- Jontell, M.; Okiji, T.; Dahlgren, U.; Bergenholtz, G. Immune defense mechanisms of the dental pulp. Crit. Rev. Oral Biol. Med. 1998, 9, 179–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, P.R.; Holder, M.J.; Smith, A.J. Inflammation and regeneration in the dentin-pulp complex: A double-edged sword. J. Endod. 2014, 40, S46–S51. [Google Scholar] [CrossRef] [PubMed]
- Farges, J.C.; Alliot-Licht, B.; Renard, E.; Ducret, M.; Gaudin, A.; Smith, A.J.; Cooper, P.R. Dental pulp defence and repair mechanisms in dental caries. Mediat. Inflamm. 2015, 2015, 230251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.; Zhang, C.; Tani-Ishii, N.; Shi, S.; Wang, C.Y. NF-κB activation in human dental pulp stem cells by TNF and LPS. J. Dent. Res. 2005, 84, 994–998. [Google Scholar] [CrossRef] [PubMed]
- Zanini, M.; Meyer, E.; Simon, S. Pulp inflammation diagnosis from clinical to inflammatory mediators: A systematic review. J. Endod. 2017, 43, 1033–1051. [Google Scholar] [CrossRef] [PubMed]
- Cooper, P.R.; Takahashi, Y.; Graham, L.W.; Simon, S.; Imazato, S.; Smith, A.J. Inflammation-regeneration interplay in the dentine-pulp complex. J. Dent. 2010, 38, 687–697. [Google Scholar] [CrossRef]
- Rechenberg, D.K.; Galicia, J.C.; Peters, O.A. Biological markers for pulpal inflammation: A systematic review. PLoS ONE 2016, 11, e0167289. [Google Scholar] [CrossRef] [Green Version]
- Hobert, O. Gene regulation by transcription factors and microRNAs. Science 2008, 319, 1785–1786. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Sonkoly, E.; Ståhle, M.; Pivarcsi, A. MicroRNAs and immunity: Novel players in the regulation of normal immune function and inflammation. Semin. Cancer Biol. 2008, 18, 131–140. [Google Scholar] [CrossRef]
- Sugatani, T.; Hruska, K.A. Impaired micro-RNA pathways diminish osteoclast differentiation and function. J. Biol. Chem. 2009, 284, 4667–4678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anokye-Danso, F.; Trivedi, C.M.; Juhr, D.; Gupta, M.; Cui, Z.; Tian, Y.; Zhang, Y.; Yang, W.; Gruber, P.J.; Epstein, J.A.; et al. Highly efficient miRNA-mediated reprogramming of mouse and human somatic cells to pluripotency. Cell Stem Cell 2011, 8, 376–388. [Google Scholar] [CrossRef] [Green Version]
- Spellicy, S.E.; Hess, D.C. The immunomodulatory capacity of induced pluripotent stem cells in the post-stroke environment. Front. Cell Dev. Biol. 2021, 9, 647415. [Google Scholar] [CrossRef] [PubMed]
- Marrazzo, P.; Pizzuti, V.; Zia, S.; Sargenti, A.; Gazzola, D.; Roda, B.; Bonsi, L.; Alviano, F. Microfluidic tools for enhanced characterization of therapeutic stem cells and prediction of their potential antimicrobial secretome. Antibiotics 2021, 10, 750. [Google Scholar] [CrossRef]
- Nara, K.; Kawashima, N.; Noda, S.; Fujii, M.; Hashimoto, K.; Tazawa, K.; Okiji, T. Anti-inflammatory roles of microRNA 21 in lipopolysaccharide-stimulated human dental pulp cells. J. Cell. Physiol. 2019, 234, 21331–21341. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.M.; Rao, D.S.; Baltimore, D. microRNA regulation of inflammatory responses. Annu. Rev. Immunol. 2012, 30, 295–312. [Google Scholar] [CrossRef]
- Taganov, K.D.; Boldin, M.P.; Chang, K.J.; Baltimore, D. NF-κB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef] [Green Version]
- Comer, B.S.; Camoretti-Mercado, B.; Kogut, P.C.; Halayko, A.J.; Solway, J.; Gerthoffer, W.T. MicroRNA-146a and microRNA-146b expression and anti-inflammatory function in human airway smooth muscle. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 307, L727–L734. [Google Scholar] [CrossRef]
- Cheng, H.S.; Sivachandran, N.; Lau, A.; Boudreau, E.; Zhao, J.L.; Baltimore, D.; Delgado-Olguin, P.; Cybulsky, M.I.; Fish, J.E. MicroRNA-146 represses endothelial activation by inhibiting pro-inflammatory pathways. EMBO Mol. Med. 2013, 5, 1017–1034. [Google Scholar] [CrossRef]
- Xie, Y.F.; Shu, R.; Jiang, S.Y.; Liu, D.L.; Ni, J.; Zhang, X.L. MicroRNA-146 inhibits pro-inflammatory cytokine secretion through IL-1 receptor-associated kinase 1 in human gingival fibroblasts. J. Inflamm. 2013, 10, 20. [Google Scholar] [CrossRef] [Green Version]
- Zhong, S.; Zhang, S.; Bair, E.; Nares, S.; Khan, A.A. Differential expression of microRNAs in normal and inflamed human pulps. J. Endod. 2012, 38, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, Z.; Yang, Y. In vitro screening of LPS-induced miRNAs in leukocytes derived from cord blood and their possible roles in regulating TLR signals. Pediatr. Res. 2014, 75, 595–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtale, G.; Mirolo, M.; Renzi, T.A.; Rossato, M.; Bazzoni, F.; Locati, M. Negative regulation of Toll-like receptor 4 signaling by IL-10-dependent microRNA-146b. Proc. Natl. Acad. Sci. USA 2013, 110, 11499–11504. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Signaling to NF-κB by Toll-like receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef]
- Hirao, K.; Yumoto, H.; Takahashi, K.; Mukai, K.; Nakanishi, T.; Matsuo, T. Roles of TLR2, TLR4, NOD2, and NOD1 in pulp fibroblasts. J. Dent. Res. 2009, 88, 762–767. [Google Scholar] [CrossRef]
- Marrazzo, P.; Mandrone, M.; Chiocchio, I.; Zambonin, L.; Barbalace, M.C.; Zalambani, C.; Angeloni, C.; Malaguti, M.; Prata, C.; Poli, F.; et al. By-product extracts from Castanea sativa counteract hallmarks of neuroinflammation in a microglial model. Antioxidants 2023, 12, 808. [Google Scholar] [CrossRef]
- Yoon, S.; Woo, S.U.; Kang, J.H.; Kim, K.; Shin, H.J.; Gwak, H.S.; Park, S.; Chwae, Y.J. NF-κB and STAT3 cooperatively induce IL6 in starved cancer cells. Oncogene 2012, 31, 3467–3481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, M.; Kawashima, N.; Tazawa, K.; Hashimoto, K.; Nara, K.; Noda, S.; Kuramoto, M.; Orikasa, S.; Nagai, S.; Okiji, T. HIF1α inhibits LPS-mediated induction of IL-6 synthesis via SOCS3-dependent CEBPβ suppression in human dental pulp cells. Biochem. Biophys. Res. Commun. 2020, 522, 308–314. [Google Scholar] [CrossRef]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jin, H.; Ao, X.; Dong, M.; Liu, S.; Lu, Y.; Niu, W. JAK2-STAT3 signaling pathway is involved in rat periapical lesions induced by Enterococcus faecalis. Oral Dis. 2019, 25, 1769–1779. [Google Scholar] [CrossRef]
- Murray, P.J. The JAK-STAT signaling pathway: Input and output integration. J. Immunol. 2007, 178, 2623–2629. [Google Scholar] [CrossRef] [Green Version]
- Xiang, M.; Birkbak, N.J.; Vafaizadeh, V.; Walker, S.R.; Yeh, J.E.; Liu, S.; Kroll, Y.; Boldin, M.; Taganov, K.; Groner, B.; et al. STAT3 induction of miR-146b forms a feedback loop to inhibit the NF-κB to IL-6 signaling axis and STAT3-driven cancer phenotypes. Sci. Signal. 2014, 7, ra11. [Google Scholar] [CrossRef] [Green Version]
- Christian, F.; Smith, E.L.; Carmody, R.J. The regulation of NF-κB subunits by phosphorylation. Cells 2016, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Milanovic, M.; Kracht, M.; Schmitz, M.L. The cytokine-induced conformational switch of nuclear factor κB p65 is mediated by p65 phosphorylation. Biochem. J. 2014, 457, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Deng, L.; Wang, C.; Spencer, E.; Yang, L.; Braun, A.; You, J.; Slaughter, C.; Pickart, C.; Chen, Z.J. Activation of the IkappaB kinase complex by TRAF6 requires a dimeric ubiquitin-conjugating enzyme complex and a unique polyubiquitin chain. Cell 2000, 103, 351–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flannery, S.; Bowie, A.G. The interleukin-1 receptor-associated kinases: Critical regulators of innate immune signalling. Biochem. Pharmacol. 2010, 80, 1981–1991. [Google Scholar] [CrossRef]
- Rhyasen, G.W.; Starczynowski, D.T. IRAK signalling in cancer. Br. J. Cancer 2015, 112, 232–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhaumik, D.; Scott, G.K.; Schokrpur, S.; Patil, C.K.; Campisi, J.; Benz, C.C. Expression of microRNA-146 suppresses NF-κB activity with reduction of metastatic potential in breast cancer cells. Oncogene 2008, 27, 5643–5647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baek, D.; Villén, J.; Shin, C.; Camargo, F.D.; Gygi, S.P.; Bartel, D.P. The impact of microRNAs on protein output. Nature 2008, 455, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Scaife, C.L.; Kuang, J.; Wills, J.C.; Trowbridge, D.B.; Gray, P.; Manning, B.M.; Eichwald, E.J.; Daynes, R.A.; Kuwada, S.K. Nuclear factor kappaB inhibitors induce adhesion-dependent colon cancer apoptosis: Implications for metastasis. Cancer Res. 2002, 62, 6870–6878. [Google Scholar]
- Pierce, J.W.; Schoenleber, R.; Jesmok, G.; Best, J.; Moore, S.A.; Collins, T.; Gerritsen, M.E. Novel inhibitors of cytokine-induced IκBα phosphorylation and endothelial cell adhesion molecule expression show anti-inflammatory effects in vivo. J. Biol. Chem. 1997, 272, 21096–21103. [Google Scholar] [CrossRef] [Green Version]
- Pardanani, A.; Tefferi, A. Targeting myeloproliferative neoplasms with JAK inhibitors. Curr. Opin. Hematol. 2011, 18, 105–110. [Google Scholar] [CrossRef]
- Quintás-Cardama, A.; Vaddi, K.; Liu, P.; Manshouri, T.; Li, J.; Scherle, P.A.; Caulder, E.; Wen, X.; Li, Y.; Waeltz, P.; et al. Preclinical characterization of the selective JAK1/2 inhibitor INCB018424: Therapeutic implications for the treatment of myeloproliferative neoplasms. Blood 2010, 115, 3109–3117. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse | Accession No. | Size, bp |
|---|---|---|---|---|
| <human> | ||||
| ACTB | 5′-GTAGCACAGCTTCTCCTTAATGTCA-3′ | 5′-CTGACTGACTACCTCATGAAGATCC-3′ | NM_001101.3 | 102 |
| IL6 | 5′-TATACCTCAAACTCCAAAAGACCAG-3′ | 5′-ACAAGAGTAACATGTGTGAAAGCAG-3′ | NM_000600.4 | 157 |
| IL8 | 5′-TCAGTGCATAAAGACATACTCCAAA-3′ | 5′-TCTTCCATCAGAAAGCTTTACAATAA-3′ | NM_000584.4 | 121 |
| IL1A | 5′-GCAGTCACATACAATTGAGTTITTG-3′ | 5′-TTACATAATCTGGATGAAGCAGTGA-3′ | NM_000575.4 | 116 |
| IL1B | 5′-CTCAAATTCCAGCTTGTTATTGATT-3′ | 5′-GTTGAAAGATGATAAGCCCACTCTA-3′ | NM_000576.2 | 124 |
| MCP1 | 5′-CACCTGCTGTTATAACTTCACCAAT-3′ | 5′-GTTGAAAGATGATAAGCCCACTCTA-3′ | NM_002982.4 | 130 |
| TNFA | 5′-CCTGGTATGAGCCCATCTATCTG-3′ | 5′-GCAATGATCCCAAAGTAGACCTG-3′ | NM_000594.3 | 130 |
| TRAF6 | 5′-ACCCTCTAACTGGTGAATAGTTTCC-3′ | 5′-CTACAAGAGAACACCCAGTCACAC-3′ | NM 145803.2 | 129 |
| IRAK1 | 5′-ACAGAAGATGGTCCAGAAGCTG-3′ | 5′-AGCTCTGAAATTCATCACTTTCTTC-3′ | NM_001569.3 | 140 |
| RELA | 5′-TTCCAAGTTCCTATAGAAGAGCAGC-3′ | 5′-TCAAAGATGGGATGAGAAAGGACAG-3′ | NM_021975.4 | 134 |
| <rat> | ||||
| Actb | 5′-GTAAAGACCTCTATGCCAACACAGT-3′ | 5′-GGAGCAATGATCTTGATCTTCATGG -3′ | NM_031144.3 | 127 |
| Il6 | 5′-TAAGGACCAAGACCATCCAACTCAT-3′ | 5′-AGTGAGGAATGTCCACAAACTGATA-3′ | NM_012589.2 | 125 |
| Il1a | 5′-CCAATCTGTACTGTTCACTTCGTTC-3′ | 5′-TTCCCGTCTTTAGATGGTTAGCTTT-3′ | NM_017019.2 | 137 |
| Il1b | 5′-AGAAGAATCTAGTTGTCCGTGTGTA-3′ | 5′-GCTTGTGCTTCATTCATAAACACTC-3′ | NM_031512.2 | 139 |
| Mcp1 | 5′-CTAAGGACTTCAGCACCTTTGAATG-3′ | 5′-GTTCTCTGTCATACTGGTCACTTCT-3′ | NM_031530.1 | 120 |
| Tnfa | 5′-AAACGGAGCTAAACTACCAGCTATC-3′ | 5′-CCTGGTCACCAAATCAGCATTATTA-3′ | NM_012675.3 | 139 |
| Traf6 | 5′-ATTGTTGAAATATGCTCTAGGCAGC-3′ | 5′-TGGAAACCAAGCTATACTGAACAGA-3′ | NM_001107754.2 | 123 |
| Irak1 | 5′-AAGTTCTCATGGTGTACAAACTCCT-3′ | 5′-GCCTTGTCTGTGCTTACATTATGAG-3′ | NM_001127555.1 | 120 |
| Rela | 5′-CTTTCTCAAGTGCCTTAATAGCAGG-3′ | 5′-TTCAGAGCTAGAAAGAGCAAGAGTC-3′ | NM_199267.2 | 121 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, P.; Sunada-Nara, K.; Kawashima, N.; Fujii, M.; Wang, S.; Kieu, T.Q.; Yu, Z.; Okiji, T. MicroRNA-146b-5p Suppresses Pro-Inflammatory Mediator Synthesis via Targeting TRAF6, IRAK1, and RELA in Lipopolysaccharide-Stimulated Human Dental Pulp Cells. Int. J. Mol. Sci. 2023, 24, 7433. https://doi.org/10.3390/ijms24087433
Han P, Sunada-Nara K, Kawashima N, Fujii M, Wang S, Kieu TQ, Yu Z, Okiji T. MicroRNA-146b-5p Suppresses Pro-Inflammatory Mediator Synthesis via Targeting TRAF6, IRAK1, and RELA in Lipopolysaccharide-Stimulated Human Dental Pulp Cells. International Journal of Molecular Sciences. 2023; 24(8):7433. https://doi.org/10.3390/ijms24087433
Chicago/Turabian StyleHan, Peifeng, Keisuke Sunada-Nara, Nobuyuki Kawashima, Mayuko Fujii, Shihan Wang, Thoai Quoc Kieu, Ziniu Yu, and Takashi Okiji. 2023. "MicroRNA-146b-5p Suppresses Pro-Inflammatory Mediator Synthesis via Targeting TRAF6, IRAK1, and RELA in Lipopolysaccharide-Stimulated Human Dental Pulp Cells" International Journal of Molecular Sciences 24, no. 8: 7433. https://doi.org/10.3390/ijms24087433