
Genome-Wide Investigation and Functional Analysis Reveal That CsGeBP4 Is Required for Tea Plant Trichome Formation
Abstract
:1. Introduction
2. Results
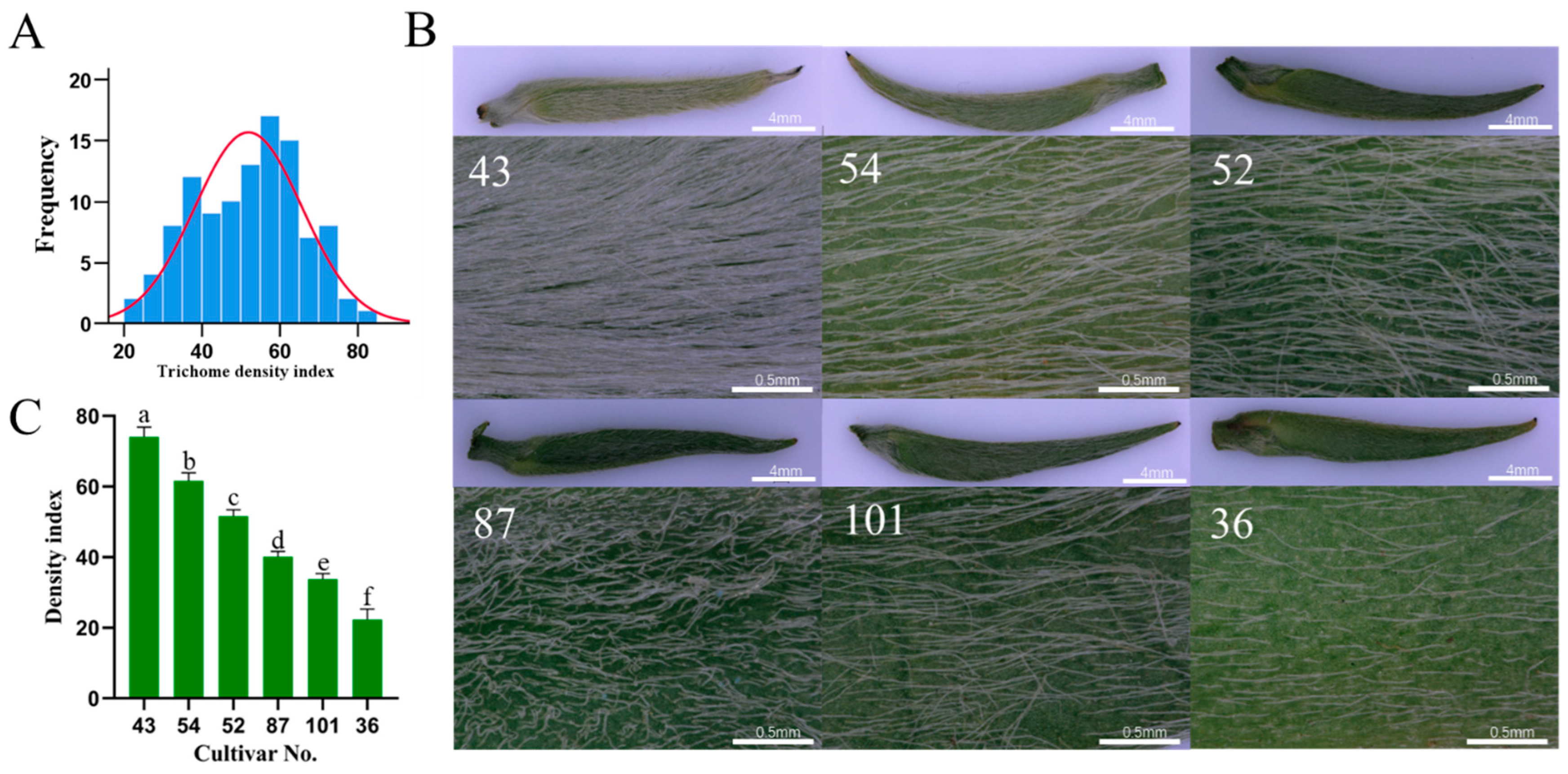
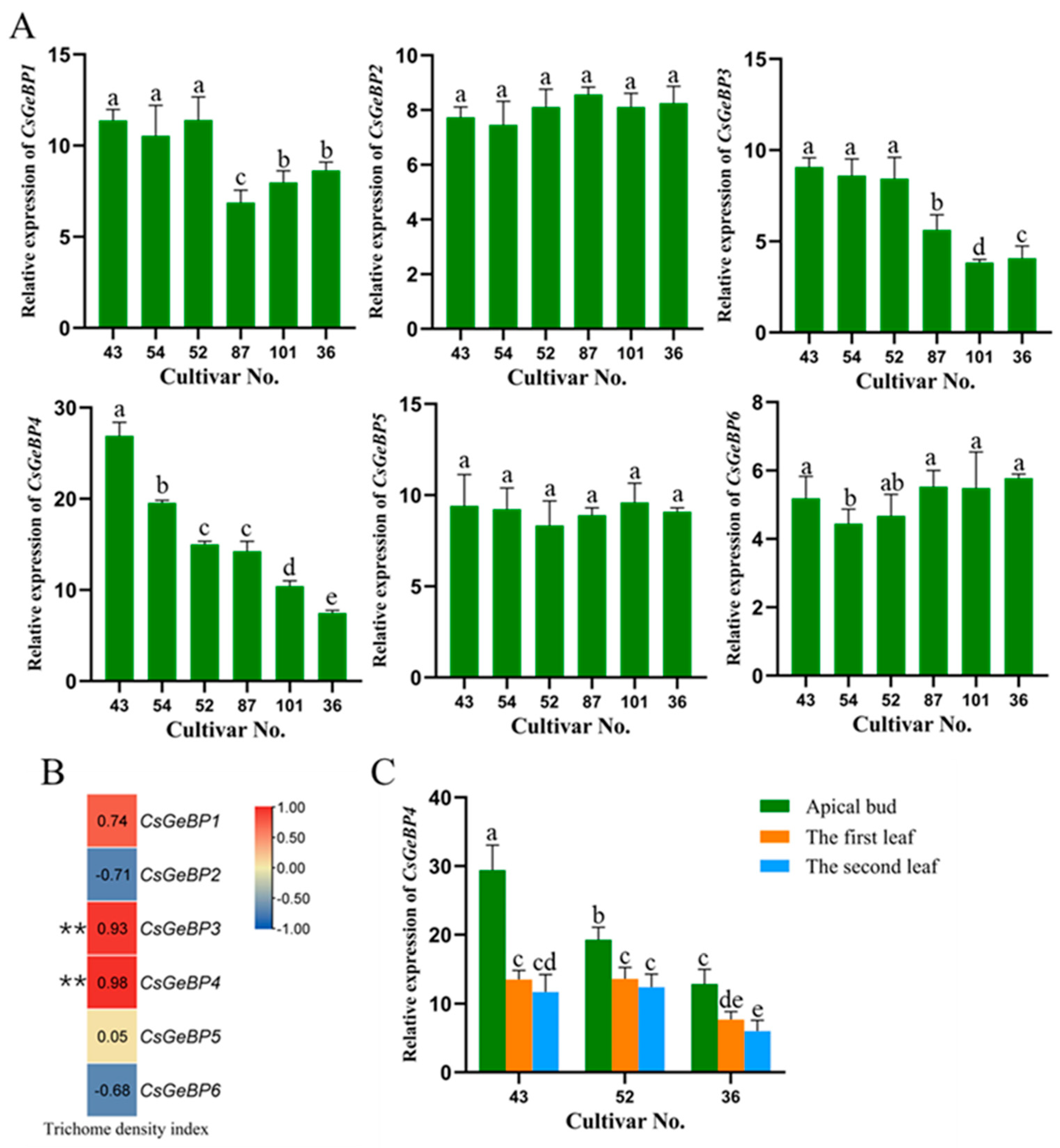
2.1. Association of Trichome Phenotype with Transcription Factor CsGeBPs
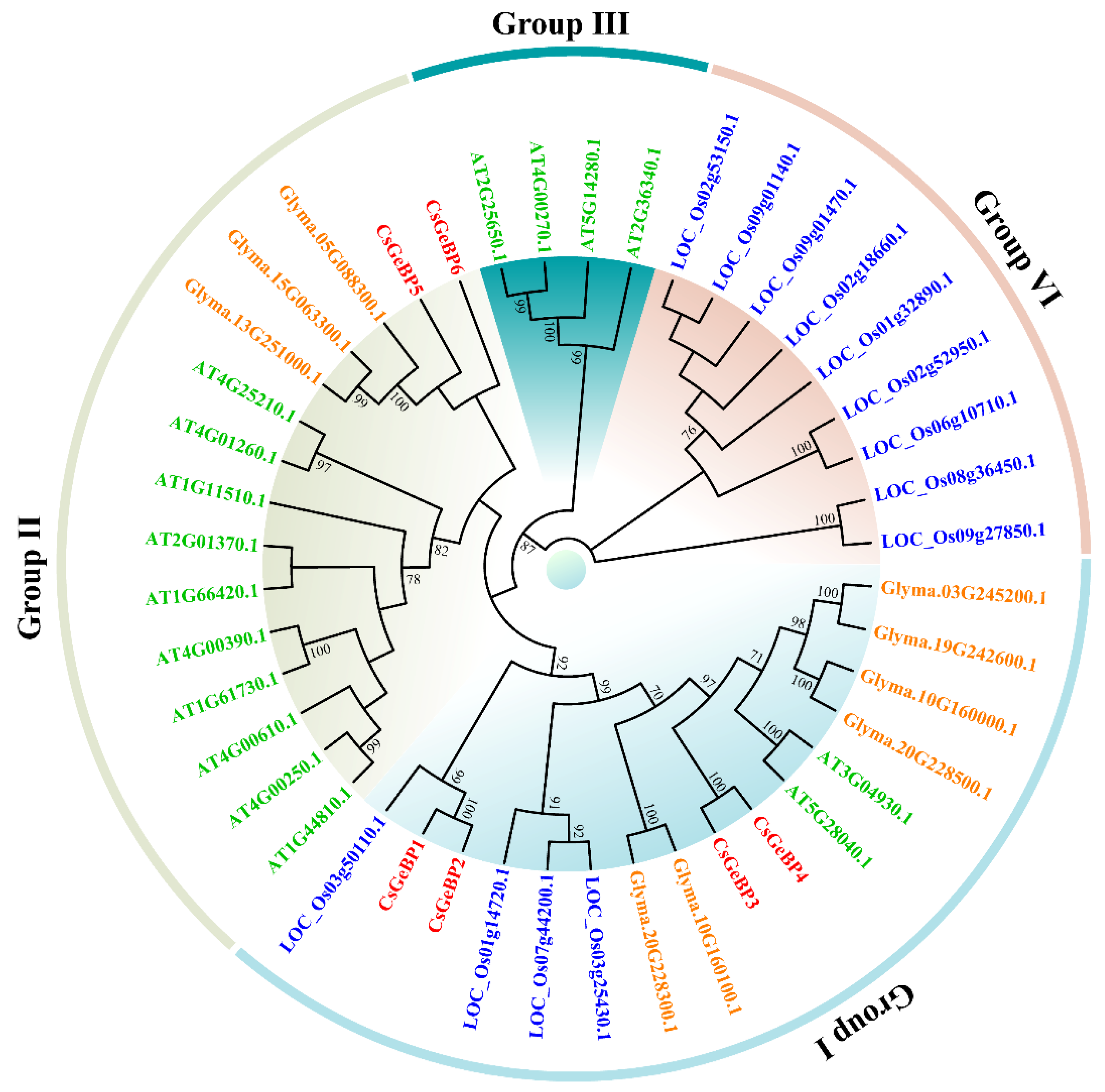
2.2. Identification and Phylogenetic Analysis of GeBP Family in Tea Plants
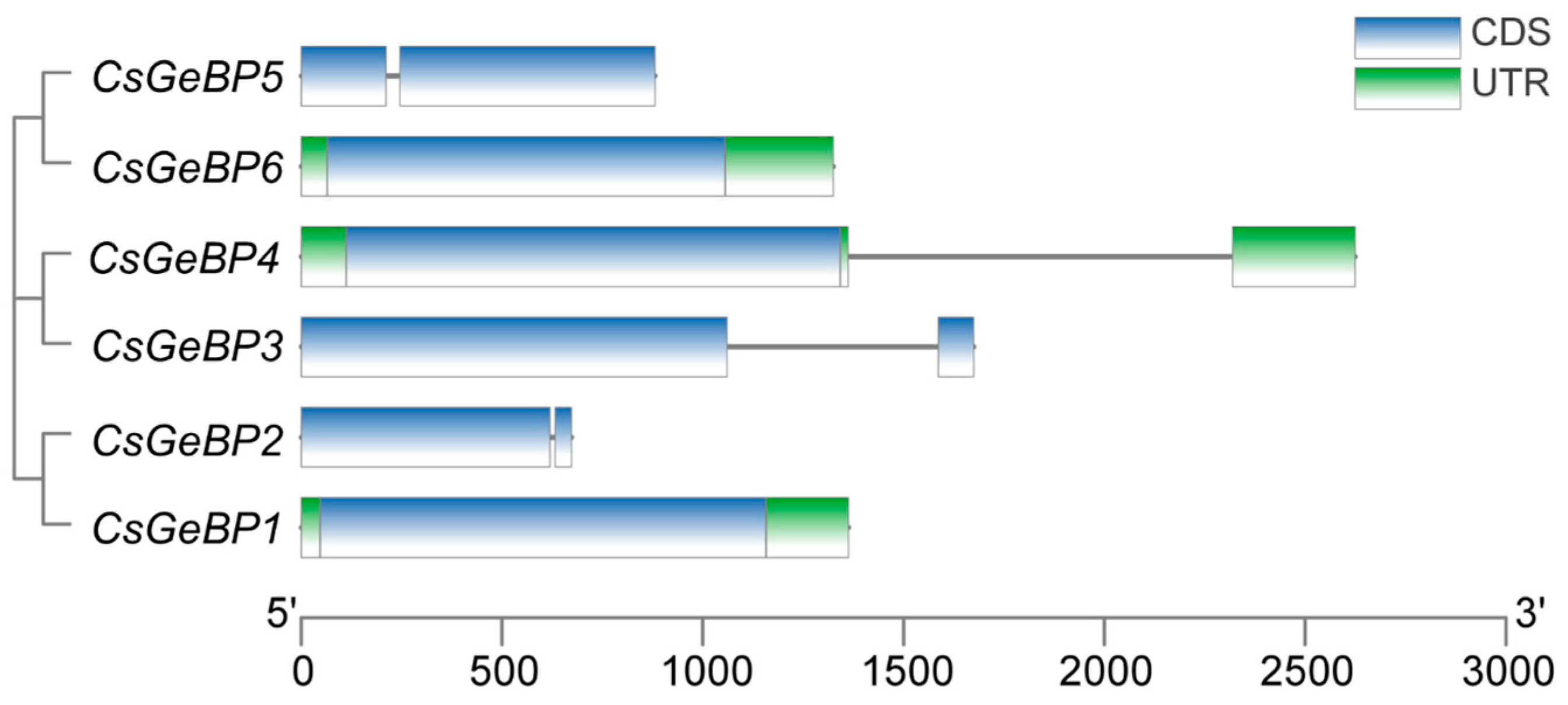
2.3. Chromosomal Distribution and Gene Structure of CsGeBPs
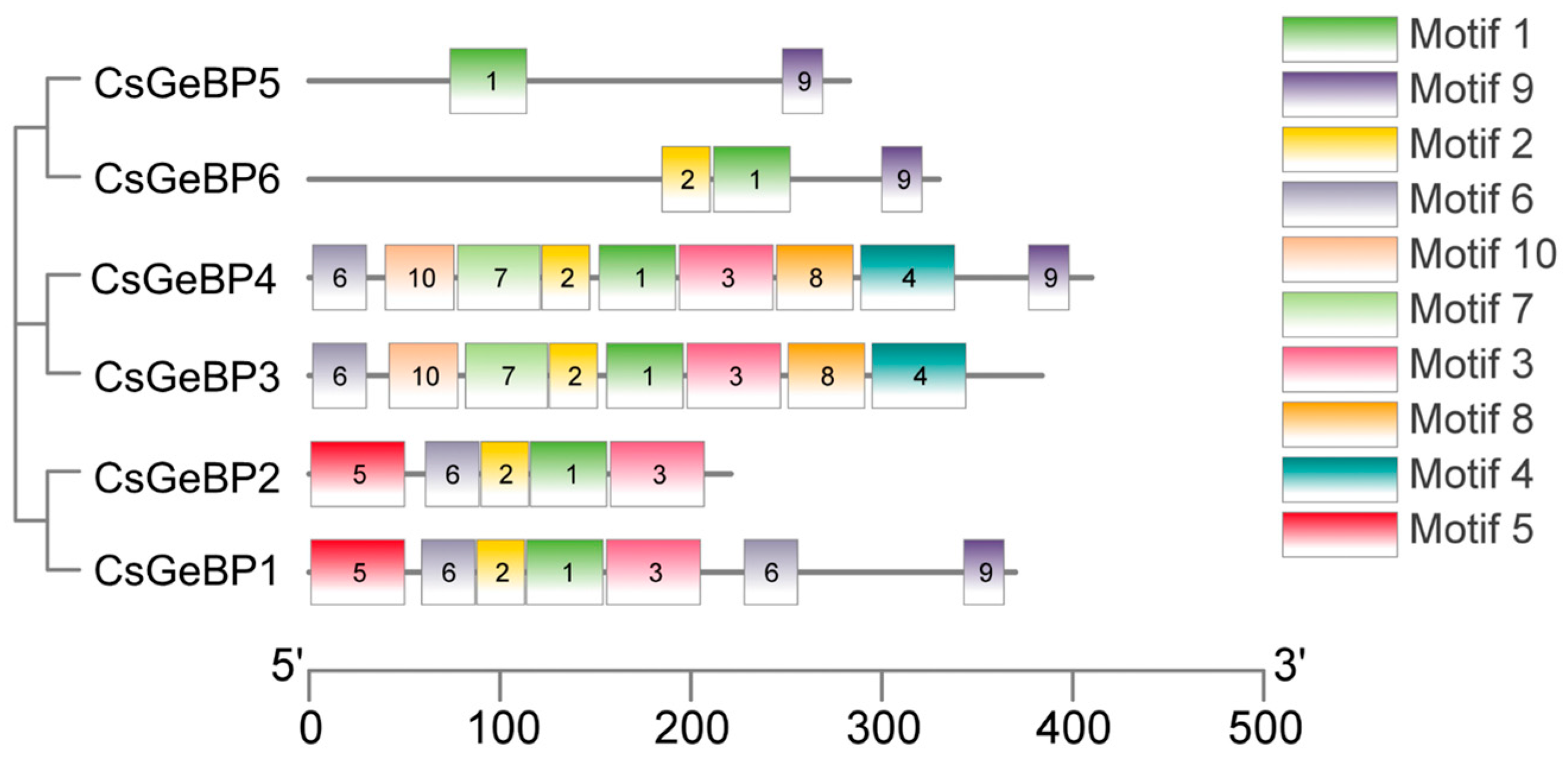
2.4. Conserved Motif Analysis of the CsGeBP Proteins
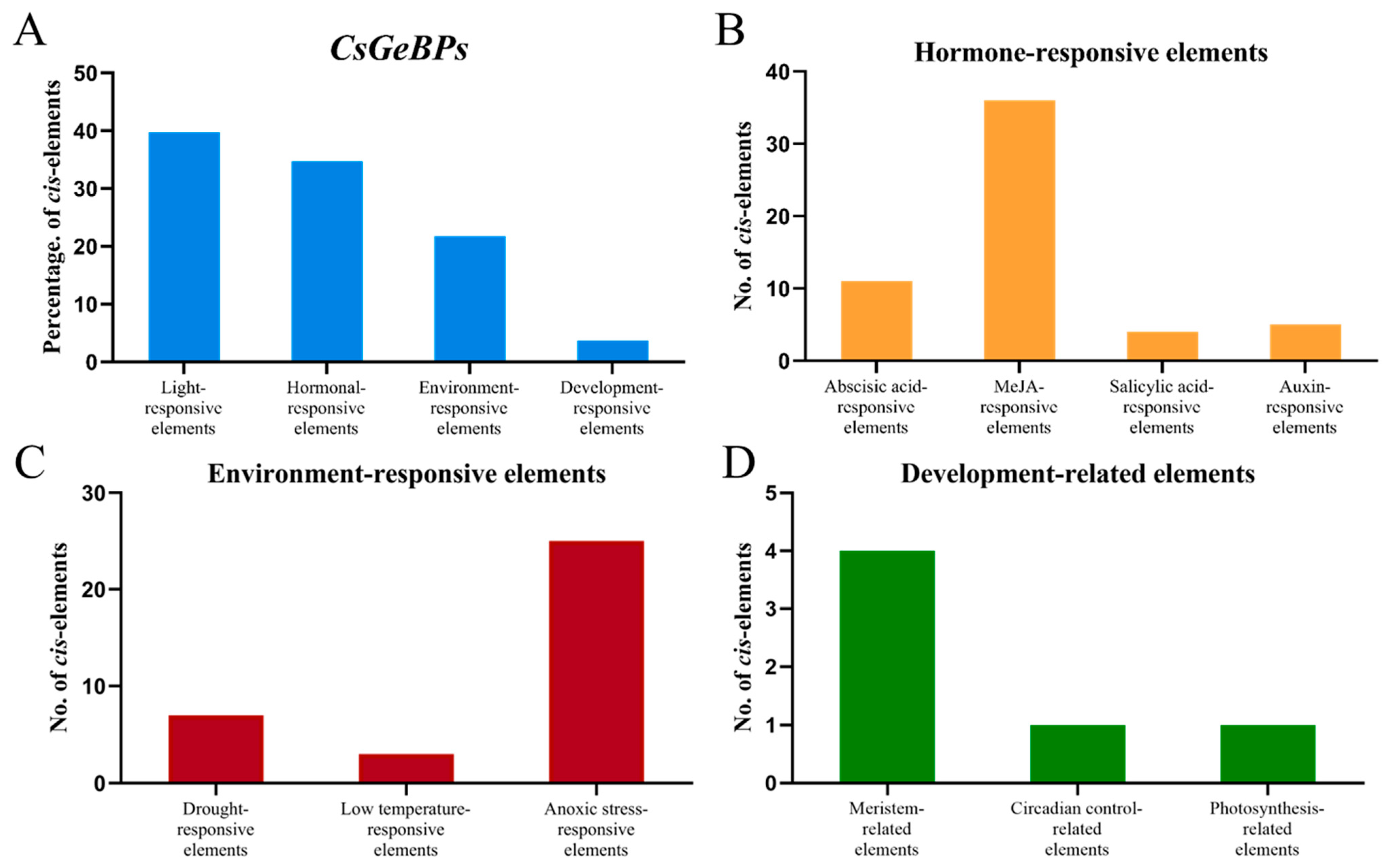
2.5. Cis-Regulatory Elements: Analysis of CsGeBP Gene Promoters
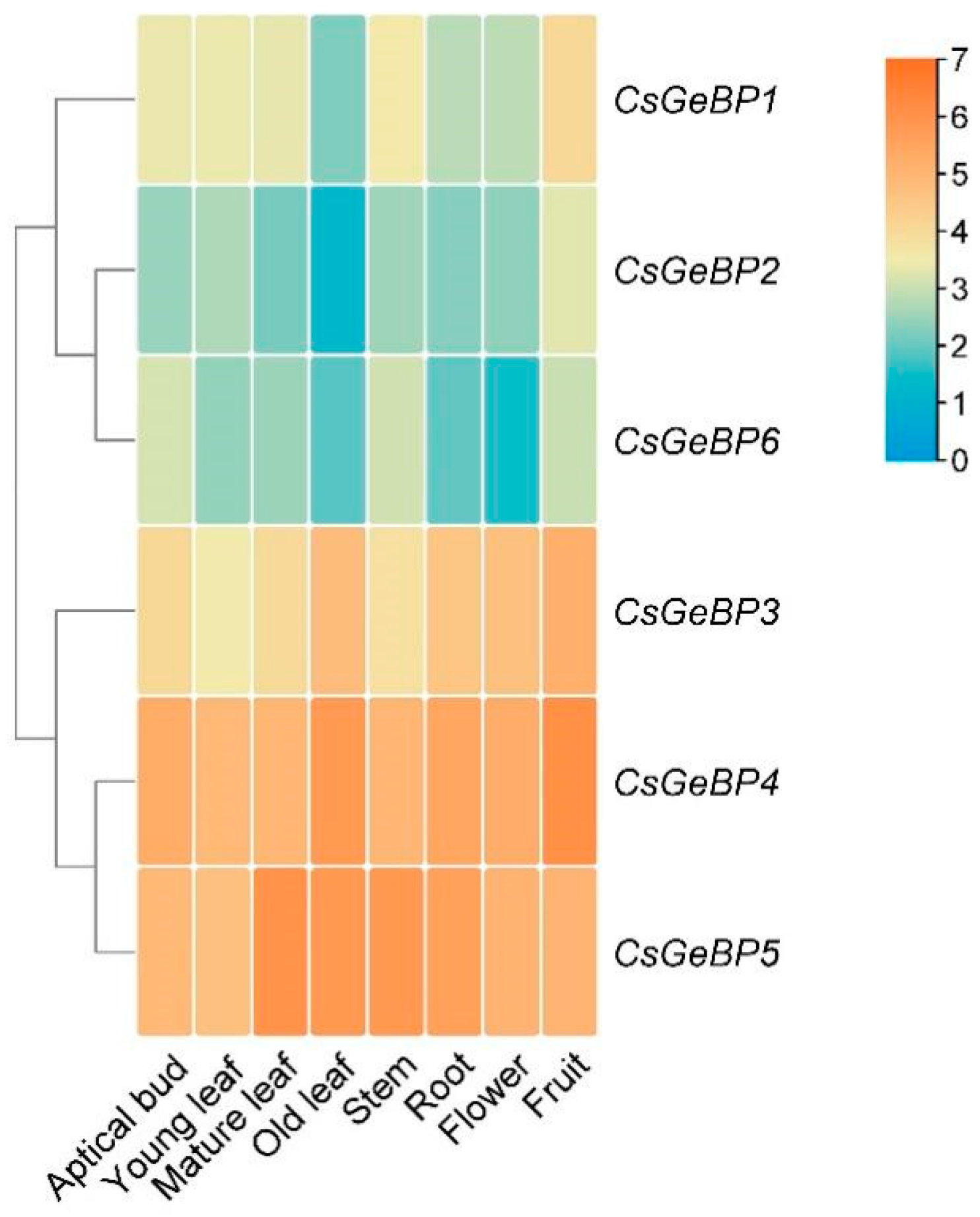
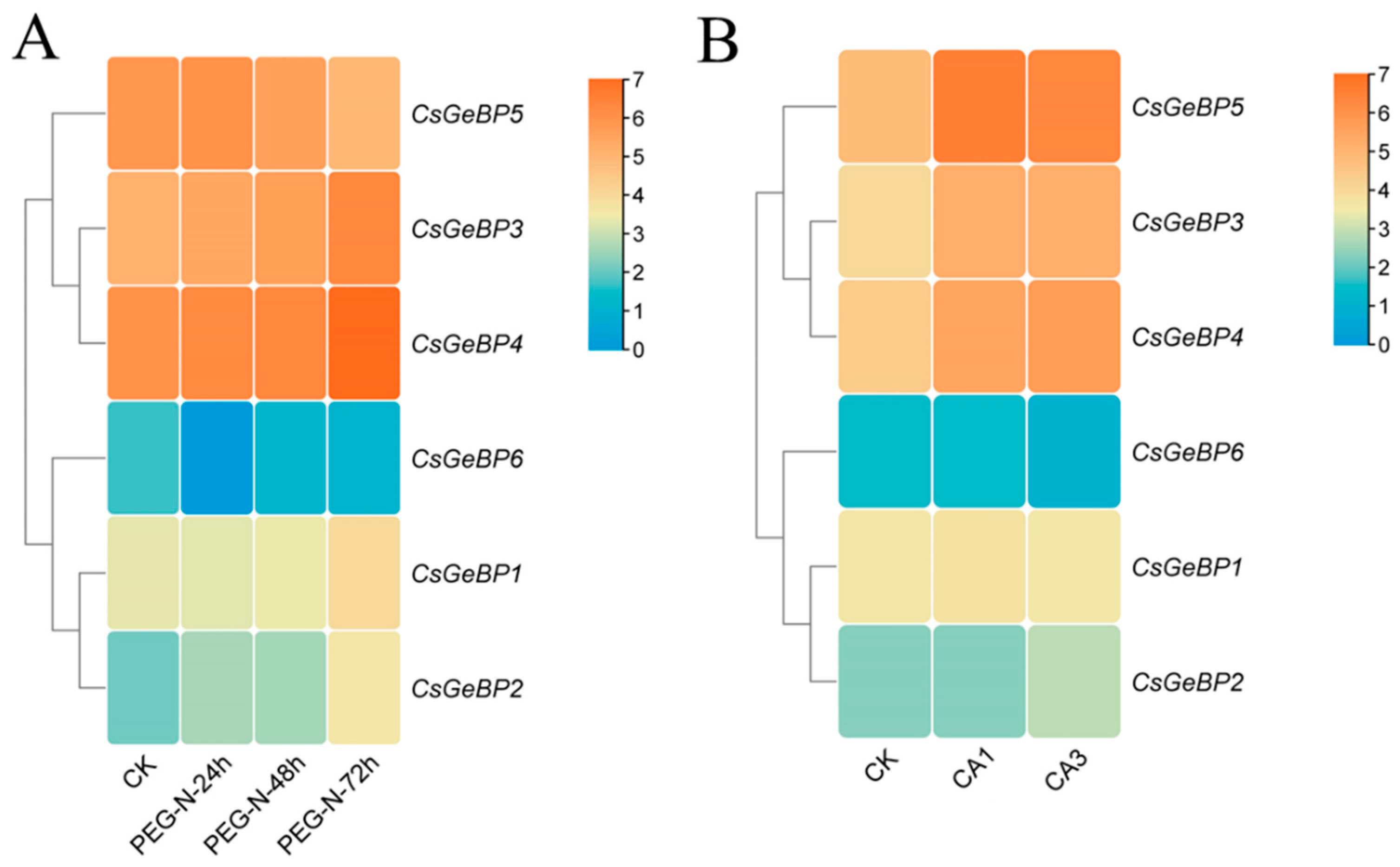
2.6. Expression Pattern Analysis of CsGeBP Gene in Tea Plants
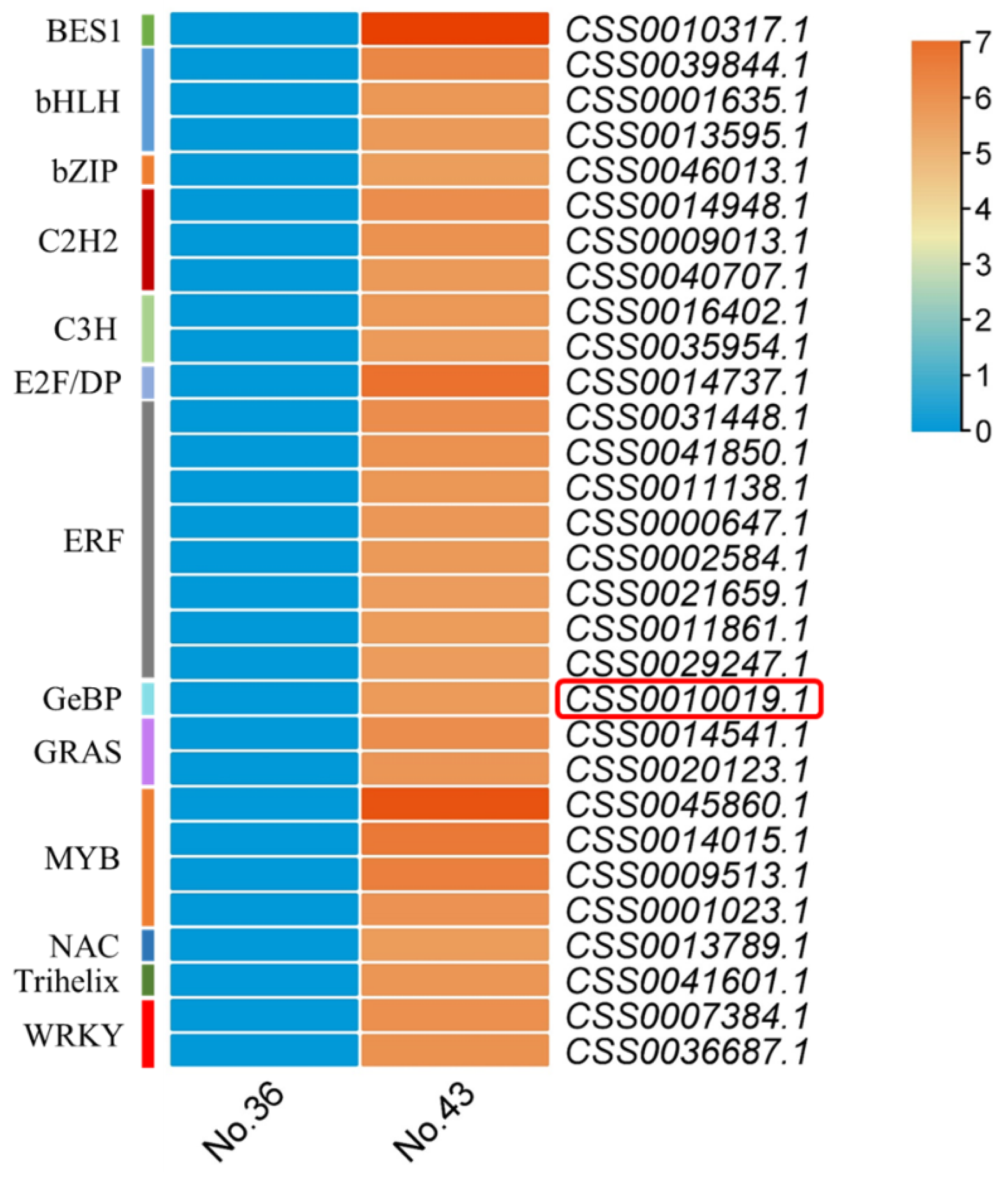
2.7. CsGeBP4 was Highly Related to the High-Dense Trichome Phenotype of Tea Plants
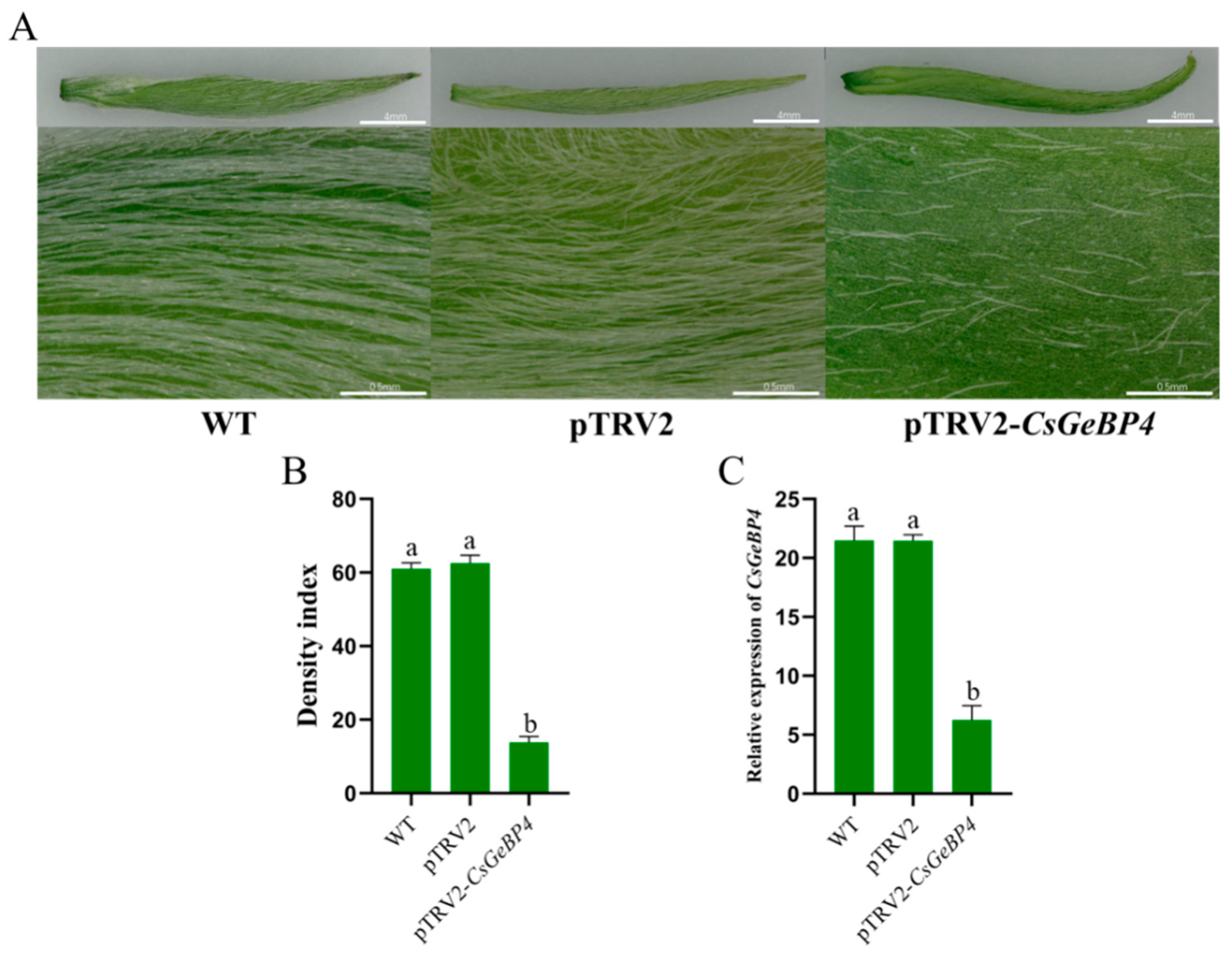
2.8. Silencing of CsGeBP4 Inhibited Trichome Development in Tea Plants
3. Discussion
3.1. Association of CsGeBP Gene Family with High-Dense Trichome Phenotypes in Tea Plants
3.2. Genome-Wide Analysis of GeBP Gene Family and Their Involvement in Tea Plant Growth and Environmental Responses
3.3. CsGeBP4 Is Required for Tea Plant Trichome Formation
4. Materials and Methods
4.1. Plant Materials and Morphological Observations
4.2. Identification of GeBP Family Genes in the Tea Plant Genome
4.3. Phylogenetic Analysis of GeBPs among Different Plant Species
4.4. Chromosomal Distribution, Gene Structure, and Conserved Motif Analysis
4.5. Identification of Putative Cis-Regulatory Elements in the Promoters of CsGeBPs
4.6. Expression Analysis of CsGeBPs in Different Tissues and in Response to Environmental Stresses
4.7. RNA Extraction and qRT-PCR Analysis
4.8. Differentially Expressed Gene Analysis by Transcriptome Sequencing
4.9. VIGS-Based Gene Silencing in Tea Plants
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, P.; Xu, Y.; Zhang, Y.; Fu, J.; Yu, S.; Guo, H.; Chen, Z.; Chen, C.; Yang, X.; Wang, S.; et al. Metabolite profiling and transcriptome analysis revealed the chemical contributions of tea trichomes to tea flavors and tea plant defenses. J. Agric. Food Chem. 2020, 68, 11389–11401. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Cao, H.-L.; Chen, D.; Lin, H.-Z.; Wang, Z.; Hu, J.; Yang, G.-Y.; Guo, Y.-Q.; Ye, N.-X.; Hao, X.-Y. Comparative transcriptome study of hairy and hairless tea plant (Camellia sinensis) shoots. J. Plant Physiol. 2018, 229, 41–52. [Google Scholar] [CrossRef]
- Sun, B.; Zhu, Z.; Liu, R.; Wang, L.; Dai, F.; Cao, F.; Liu, S. TRANSPARENT TESTA GLABRA1 (TTG1) regulates leaf trichome density in tea Camellia sinensis. Nord. J. Bot. 2020, 38, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Fu, J.; Xu, Y.; Shen, Y.; Zhang, Y.; Ye, Z.; Tong, W.; Zeng, X.; Yang, J.; Tang, D.; et al. CsMYB1 integrates the regulation of trichome development and catechins biosynthesis in tea plant domestication. New Phytol. 2022, 234, 902–917. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tian, N.; Hu, M.; Sandhu, D.; Jin, Q.; Gu, M.; Zhang, X.; Peng, Y.; Zhang, J.; Chen, Z.; et al. Comparative transcriptome analysis reveals key pathways and genes involved in trichome development in tea plant (Camellia sinensis). Front. Plant Sci. 2022, 13, 997778. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zeng, L.; Liao, Y.; Tang, J.; Yang, Z. Evaluation of the contribution of trichomes to metabolite compositions of tea (Camellia sinensis) leaves and their products. LWT 2020, 122, 109023. [Google Scholar] [CrossRef]
- Balkunde, R.; Pesch, M.; Hülskamp, M. Trichome Patterning in Arabidopsis thaliana from genetic to molecular models. Curr. Top. Dev. Biol. 2010, 91, 299–321. [Google Scholar] [CrossRef]
- Bleeker, P.M.; Mirabella, R.; Diergaarde, P.J.; VanDoorn, A.; Tissier, A.; Kant, M.R.; Prins, M.; de Vos, M.; Haring, M.A.; Schuurink, R.C. Improved herbivore resistance in cultivated tomato with the sesquiterpene biosynthetic pathway from a wild relative. Proc. Natl. Acad. Sci. USA 2012, 109, 20124–20129. [Google Scholar] [CrossRef] [Green Version]
- Gonzales-Vigil, E.; Hufnagel, D.E.; Kim, J.; Last, R.L.; Barry, C.S. Evolution of TPS20-related terpene synthases influences chemical diversity in the glandular trichomes of the wild tomato relative Solanum habrochaites. Plant J. 2012, 71, 921–935. [Google Scholar] [CrossRef] [Green Version]
- Luu, V.T.; Weinhold, A.; Ullah, C.; Dressel, S.; Schoettner, M.; Gase, K.; Gaquerel, E.; Xu, S.; Baldwin, I.T. O-Acyl sugars protect a wild tobacco from both native fungal pathogens and a specialist herbivore. Plant Physiol. 2017, 174, 370–386. [Google Scholar] [CrossRef] [Green Version]
- Schilmiller, A.L.; Last, R.L.; Pichersky, E. Harnessing plant trichome biochemistry for the production of useful compounds. Plant J. 2008, 54, 702–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, P.; Wang, J.; Zhou, Y.; Gao, L.; Wang, J.; Gong, C. Adaptional evolution of trichome in Caragana korshinskii to natural drought stress on the Loess Plateau, China. Ecol. Evol. 2016, 6, 3786–3795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Shen, C.; Meng, P.; Tan, G.; Lv, L. Analysis and review of trichomes in plants. BMC Plant Biol. 2021, 21, 70. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, A.M.; Schena, M.; Walbot, V.; Davis, R.W. Epidermal cell fate determination in Arabidopsis: Patterns defined by a steroid-inducible regulator. Science 1994, 266, 436–439. [Google Scholar] [CrossRef]
- Chien, J.C.; Sussex, I.M. Differential regulation of trichome formation on the adaxial and abaxial leaf surfaces by gibberellins and photoperiod in Arabidopsis thaliana (L.) Heynh. Plant Physiol. 1996, 111, 1321–1328. [Google Scholar] [CrossRef] [Green Version]
- Larkin, J.; Young, N.; Prigge, M.; Marks, M.D. The control of trichome spacing and number in Arabidopsis. Development 1996, 122, 997–1005. [Google Scholar] [CrossRef]
- Telfer, A.; Bollman, K.; Poethig, R. Phase change and the regulation of trichome distribution in Arabidopsis thaliana. Development 1997, 124, 645–654. [Google Scholar] [CrossRef]
- Perazza, D.; Vachon, G.; Herzog, M. Gibberellins promote trichome formation by up-regulating GLABROUS1 in Arabidopsis. Plant Physiol. 1998, 117, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.-Y.; Miao, H.; Ding, L.-H.; Wehner, T.C.; Liu, P.-N.; Wang, Y.; Zhang, S.-P.; Gu, X.-F. A new glabrous gene (CsGl3) identified in trichome development in cucumber (Cucumis sativus L.). PLoS ONE 2016, 11, e0148422. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Li, H.; Zhang, J.; Luo, Z.; Gong, P.; Zhang, C.; Li, J.; Wang, T.; Zhang, Y.; Lu, Y.; et al. A regulatory gene induces trichome formation and embryo lethality in tomato. Proc. Natl. Acad. Sci. USA 2011, 108, 11836–11841. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Lu, Y.; Yuan, Y.; Zhang, X.; Geng, J.; Chen, Y.; Cloutier, S.; McVetty, P.B.E.; Li, G. Map-based cloning and characterization of a gene controlling hairiness and seed coat color traits in Brassica rapa. Plant Mol. Biol. 2009, 69, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Gruber, M.; Alahakoon, U.; Taheri, A.; Nagubushana, N.; Zhou, R.; Aung, B.; Sharpe, A.; Hannoufa, A.; Bonham-Smith, P.; Hegedus, D.D.D. The biochemical composition and transcriptome of cotyledons from Brassica napus lines expressing the AtGL3 transcription factor and exhibiting reduced flea beetle feeding. BMC Plant Biol. 2018, 18, 64. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, J.-G. Arabidopsis transient expression analysis reveals that activation of GLABRA2 may require concurrent binding of GLABRA1 and GLABRA3 to the promoter of GLABRA2. Plant Cell Physiol. 2008, 49, 1792–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, C.S.; Kolevski, B.; Smyth, D.R. TRANSPARENT TESTA GLABRA2, a trichome and seed coat development gene of Arabidopsis, encodes a WRKY transcription factor. Plant Cell 2002, 14, 1359–1375. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Morohashi, K.; Hatlestad, G.; Grotewold, E.; Lloyd, A. The TTG1-bHLH-MYB complex controls trichome cell fate and patterning through direct targeting of regulatory loci. Development 2008, 135, 1991–1999. [Google Scholar] [CrossRef] [Green Version]
- Tominaga-Wada, R.; Nukumizu, Y.; Sato, S.; Kato, T.; Tabata, S.; Wada, T. Functional divergence of MYB-Related genes, WEREWOLF and AtMYB23 in Arabidopsis. Biosci. Biotechnol. Biochem. 2012, 76, 883–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Wang, X.; Zhu, D.; Cui, S.; Li, X.; Cao, Y.; Ma, L. A single amino acid substitution in IIIf subfamily of basic helix-loop-helix transcription factor AtMYC1 leads to trichome and root hair patterning defects by abolishing its interaction with partner proteins in arabidopsis. J. Biol. Chem. 2012, 287, 14109–14121. [Google Scholar] [CrossRef] [Green Version]
- Li, S.F.; Milliken, O.N.; Pham, H.; Seyit, R.; Napoli, R.; Preston, J.; Koltunow, A.M.; Parish, R.W. The Arabidopsis MYB5 transcription factor regulates mucilage synthesis, seed coat development, and trichome morphogenesis. Plant Cell 2009, 21, 72–89. [Google Scholar] [CrossRef] [Green Version]
- Kirik, V.; Lee, M.M.; Wester, K.; Herrmann, U.; Zheng, Z.; Oppenheimer, D.; Schiefelbein, J.; Hulskamp, M. Functional diversification of MYB23 and GL1 genes in trichome morphogenesis and initiation. Development 2005, 132, 1477–1485. [Google Scholar] [CrossRef] [Green Version]
- Payne, C.T.; Zhang, F.; Lloyd, A.M. GL3 encodes a bHLH protein that regulates trichome development in arabidopsis through interaction with GL1 and TTG1. Genetics 2000, 156, 1349–1362. [Google Scholar] [CrossRef]
- Zhang, F.; Gonzalez, A.; Zhao, M.; Payne, C.T.; Lloyd, A. A network of redundant bHLH proteins functions in all TTG1-dependent pathways of Arabidopsis. Development 2003, 130, 4859–4869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhardt, C.; Lee, M.M.; Gonzalez, A.; Zhang, F.; Lloyd, A.; Schiefelbein, J. The bHLH genes GLABRA3 (GL3) and ENHANCER OF GLABRA3 (EGL3) specify epidermal cell fate in the Arabidopsis root. Development 2003, 130, 6431–6439. [Google Scholar] [CrossRef] [Green Version]
- Chopra, D.; Mapar, M.; Stephan, L.; Albani, M.C.; Deneer, A.; Coupland, G.; Willing, E.-M.; Schellmann, S.; Schneeberger, K.; Fleck, C.; et al. Genetic and molecular analysis of trichome development in Arabis alpina. Proc. Natl. Acad. Sci. USA 2019, 116, 12078–12083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Chopra, D.; Schrader, A.; Hülskamp, M. Evolutionary comparison of competitive protein-complex formation of MYB, bHLH, and WDR proteins in plants. J. Exp. Bot. 2019, 70, 3197–3209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Gong, X.; Cao, W.; Zhao, J.; Fu, L.; Wang, X.; Schumaker, K.S.; Guo, Y. SAD2 in Arabidopsis functions in trichome initiation through mediating GL3 function and regulating GL1, TTG1 and GL2 Expression. J. Integr. Plant Biol. 2008, 50, 906–917. [Google Scholar] [CrossRef] [PubMed]
- Szymanski, D.B.; Lloyd, A.M.; Marks, M. Progress in the molecular genetic analysis of trichome initiation and morphogenesis in Arabidopsis. Trends Plant Sci. 2000, 5, 214–219. [Google Scholar] [CrossRef]
- Das, P.; Chettri, V.; Ghosh, S.; Ghosh, C. Micromorphological studies of the leaf and stem of Camellia sinensis (L.) Kuntze with reference to their taxonomic significance. Microsc. Res. Tech. 2022. [Google Scholar] [CrossRef]
- Ruiqing, S.; Nan, Y.; Zhixin, L.; Yuqi, X.; Yu, S.; Qingyuan, G.; Jinji, P.; Zhiguo, D.; He, Z. Identification and expression analysis of mango GeBP transcription factor family genes. Non-wood For. Res. 2021, 39, 137–147. [Google Scholar] [CrossRef]
- Wakamatsu, J.; Wada, T.; Tanaka, W.; Fujii, S.; Fujikawa, Y.; Sambongi, Y.; Tominaga, R. Identification of six CPC-like genes and their differential expression in leaves of tea plant, Camellia sinensis. J. Plant Physiol. 2021, 263, 153465. [Google Scholar] [CrossRef]
- Li, G.; Li, Y.; Yao, X.; Lu, L. Establishment of a virus-induced gene-silencing (VIGS) system in tea plant and its use in the functional analysis of CsTCS1. Int. J. Mol. Sci. 2022, 24, 392. [Google Scholar] [CrossRef]
- Perazza, D.; Laporte, F.; Balagué, C.; Chevalier, F.; Remo, S.; Bourge, M.; Larkin, J.; Herzog, M.; Vachon, G. GeBP/GPL transcription factors regulate a subset of CPR5-dependent processes. Plant Physiol. 2011, 157, 1232–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curaba, J.; Herzog, M.; Vachon, G. GeBP, the first member of a new gene family in Arabidopsis, encodes a nuclear protein with DNA-binding activity and is regulated by KNAT1. Plant J. 2003, 33, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Chuck, G.; Lincoln, C.; Hake, S. KNAT1 induces lobed leaves with ectopic meristems when overexpressed in Arabidopsis. Plant Cell 1996, 8, 1277–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dill, A.; Jung, H.-S.; Sun, T.-P. The DELLA motif is essential for gibberellin-induced degradation of RGA. Proc. Natl. Acad. Sci. USA 2001, 98, 14162–14167. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhang, Q.; He, Y.; Liu, W.; Xu, Y.; Liu, K.; Xian, F.; Li, J.; Hu, J. Genome-wide identification, expansion mechanism and expression profiling analysis of GLABROUS1 enhancer-binding protein (GeBP) gene family in Gramineae crops. Int. J. Mol. Sci. 2021, 22, 8758. [Google Scholar] [CrossRef]
- Jasinski, S.; Piazza, P.; Craft, J.; Hay, A.; Woolley, L.; Rieu, I.; Phillips, A.; Hedden, P.; Tsiantis, M. KNOX action in Arabidopsis is mediated by coordinate regulation of cytokinin and gibberellin activities. Curr. Biol. 2005, 15, 1560–1565. [Google Scholar] [CrossRef] [Green Version]
- Yanai, O.; Shani, E.; Dolezal, K.; Tarkowski, P.; Sablowski, R.; Sandberg, G.; Samach, A.; Ori, N. Arabidopsis KNOXI proteins activate cytokinin biosynthesis. Curr. Biol. 2005, 15, 1566–1571. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, F.; Perazza, D.; Laporte, F.; Le Hénanff, G.; Hornitschek, P.; Bonneville, J.-M.; Herzog, M.; Vachon, G. GeBP and GeBP-Like proteins are noncanonical leucine-zipper transcription factors that regulate cytokinin response in Arabidopsis. Plant Physiol. 2008, 146, 1142–1154. [Google Scholar] [CrossRef] [Green Version]
- Khare, D.; Mitsuda, N.; Lee, S.; Song, W.; Hwang, D.; Ohme-Takagi, M.; Martinoia, E.; Lee, Y.; Hwang, J. Root avoidance of toxic metals requires the GeBP-LIKE 4 transcription factor in Arabidopsis thaliana. New Phytol. 2017, 213, 1257–1273. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Liu, Y.; Liu, C.; Zhang, F.; Wei, J.; Li, B. Genome-wide characterization and expression analysis of GeBP family genes in Soybean. Plants 2022, 11, 1848. [Google Scholar] [CrossRef]
- Xuemeng, S.; Kebin, Y.; Jingjing, S. Genome wide identification and expression analysis of GeBP transcription factor gene family in moso bamboo. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2020, 44, 41–48. [Google Scholar] [CrossRef]
- Kai, C.; Jinqiu, L.; Haihui, S.; Zhenzhu, Z.; Xiuling, C.; Wang, A. Identification, evolution and expression analysis of GeBP transcription factors family in Tomato. Mol. Plant Breed. 2017, 15, 3438–3445. [Google Scholar] [CrossRef]
- Wang, Y.; Wan, C.; Li, L.; Xiang, Z.; Wang, J.; Li, Y.; Zhao, D. Transcriptomic analysis of Yunwu Tribute Tea leaves under cold stress. Curr. Issues Mol. Biol. 2023, 45, 699–720. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.; Li, F.; Tong, W.; Li, P.; Wu, Q.; Zhao, H.; Ge, R.; Li, R.; Li, Y.; Zhang, Z.; et al. Tea plant information archive: A comprehensive genomics and bioinformatics platform for tea plant. Plant Biotechnol. J. 2019, 17, 1938–1953. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Yang, H.; Wang, S.; Zhao, J.; Liu, C.; Gao, L.; Xia, E.; Lu, Y.; Tai, Y.; She, G.; et al. Draft genome sequence of Camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality. Proc. Natl. Acad. Sci. USA 2018, 115, E4151–E4158. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Cai, M.; Yu, X.; Wang, L.; Guo, C.; Ming, R.; Zhang, J. Transcriptome dynamics of Camellia sinensis in response to continuous salinity and drought stress. Tree Genet. Genomes 2017, 13, 78. [Google Scholar] [CrossRef]
- Wang, X.-C.; Zhao, Q.-Y.; Ma, C.-L.; Zhang, Z.-H.; Cao, H.-L.; Kong, Y.-M.; Yue, C.; Hao, X.-Y.; Chen, L.; Ma, J.-Q.; et al. Global transcriptome profiles of Camellia sinensis during cold acclimation. BMC Genom. 2013, 14, 415. [Google Scholar] [CrossRef] [Green Version]
- Xia, E.; Tong, W.; Hou, Y.; An, Y.; Chen, L.; Wu, Q.; Liu, Y.; Yu, J.; Li, F.; Li, R.; et al. The reference genome of tea plant and resequencing of 81 diverse accessions provide insights into its genome evolution and adaptation. Mol. Plant 2020, 13, 1013–1026. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, G.; Li, X.; Shen, Q.; Wu, Q.; Zhuang, J.; Zhang, X.; Xia, E.; Zhang, Z.; Qian, Y.; et al. Comparative analysis of phenolic compound metabolism among tea plants in the section Thea of the genus Camellia. Food Res. Int. 2020, 135, 109276. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Phylogenies and the comparative method. Am. Nat. 1985, 125, 1–15. [Google Scholar] [CrossRef]
- Wang, L.; Stegemann, J.P. Extraction of high quality RNA from polysaccharide matrices using cetlytrimethylammonium bromide. Biomaterials 2010, 31, 1612–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Chromosome No. | Length (bp) | Intron | Exon | Amino Acid (aa) | Molecular Weight (Da) | Isoelectric Point | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|
| CSS0047893.1 | CsGeBP1 | Chr1 | 1363 | 0 | 1 | 370 | 40,951.80 | 8.28 | Nucleus |
| CSS0008596.1 | CsGeBP2 | Chr1 | 673 | 1 | 2 | 221 | 24,638.69 | 9.85 | Nucleus |
| CSS0012610.1 | CsGeBP3 | Chr6 | 1675 | 1 | 2 | 384 | 42,104.87 | 4.63 | Nucleus |
| CSS0010019.1 | CsGeBP4 | Chr6 | 2624 | 1 | 2 | 410 | 45,031.43 | 4.65 | Nucleus |
| CSS0022087.1 | CsGeBP5 | Chr7 | 881 | 1 | 2 | 283 | 31,931.65 | 9.40 | Nucleus |
| CSS0046322.1 | CsGeBP6 | Chr14 | 1325 | 0 | 1 | 330 | 36,379.28 | 5.41 | Nucleus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Zhou, W.; Yao, X.; Zhao, Q.; Lu, L. Genome-Wide Investigation and Functional Analysis Reveal That CsGeBP4 Is Required for Tea Plant Trichome Formation. Int. J. Mol. Sci. 2023, 24, 5207. https://doi.org/10.3390/ijms24065207
Zhou H, Zhou W, Yao X, Zhao Q, Lu L. Genome-Wide Investigation and Functional Analysis Reveal That CsGeBP4 Is Required for Tea Plant Trichome Formation. International Journal of Molecular Sciences. 2023; 24(6):5207. https://doi.org/10.3390/ijms24065207
Chicago/Turabian StyleZhou, Hao, Wei Zhou, Xinzhuan Yao, Qi Zhao, and Litang Lu. 2023. "Genome-Wide Investigation and Functional Analysis Reveal That CsGeBP4 Is Required for Tea Plant Trichome Formation" International Journal of Molecular Sciences 24, no. 6: 5207. https://doi.org/10.3390/ijms24065207