
Timing Expression of miR203a-3p during OA Disease: Preliminary In Vitro Evidence
 , , ,
, , ,  , and
, and
Abstract
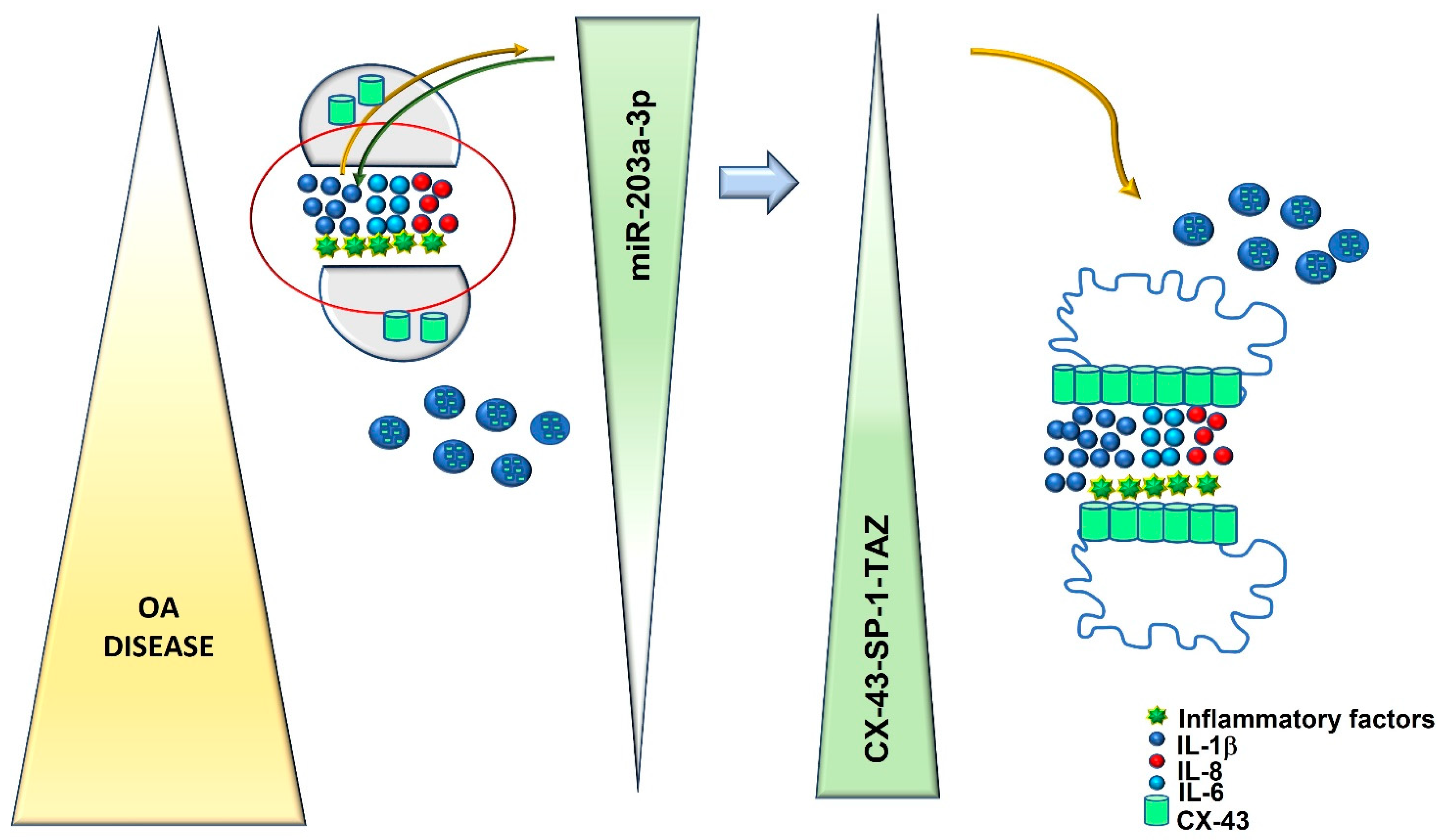
:1. Introduction
2. Results
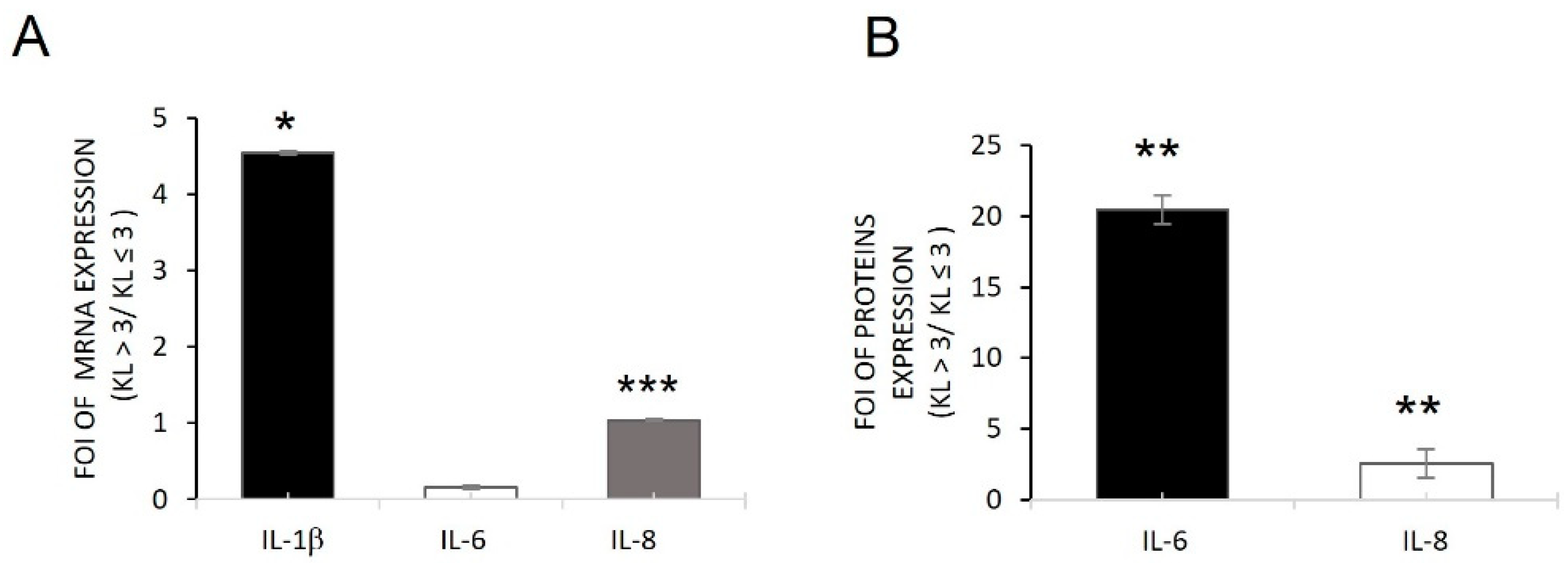
2.1. Inflammatory Conditions of OA Patient-Derived Cells
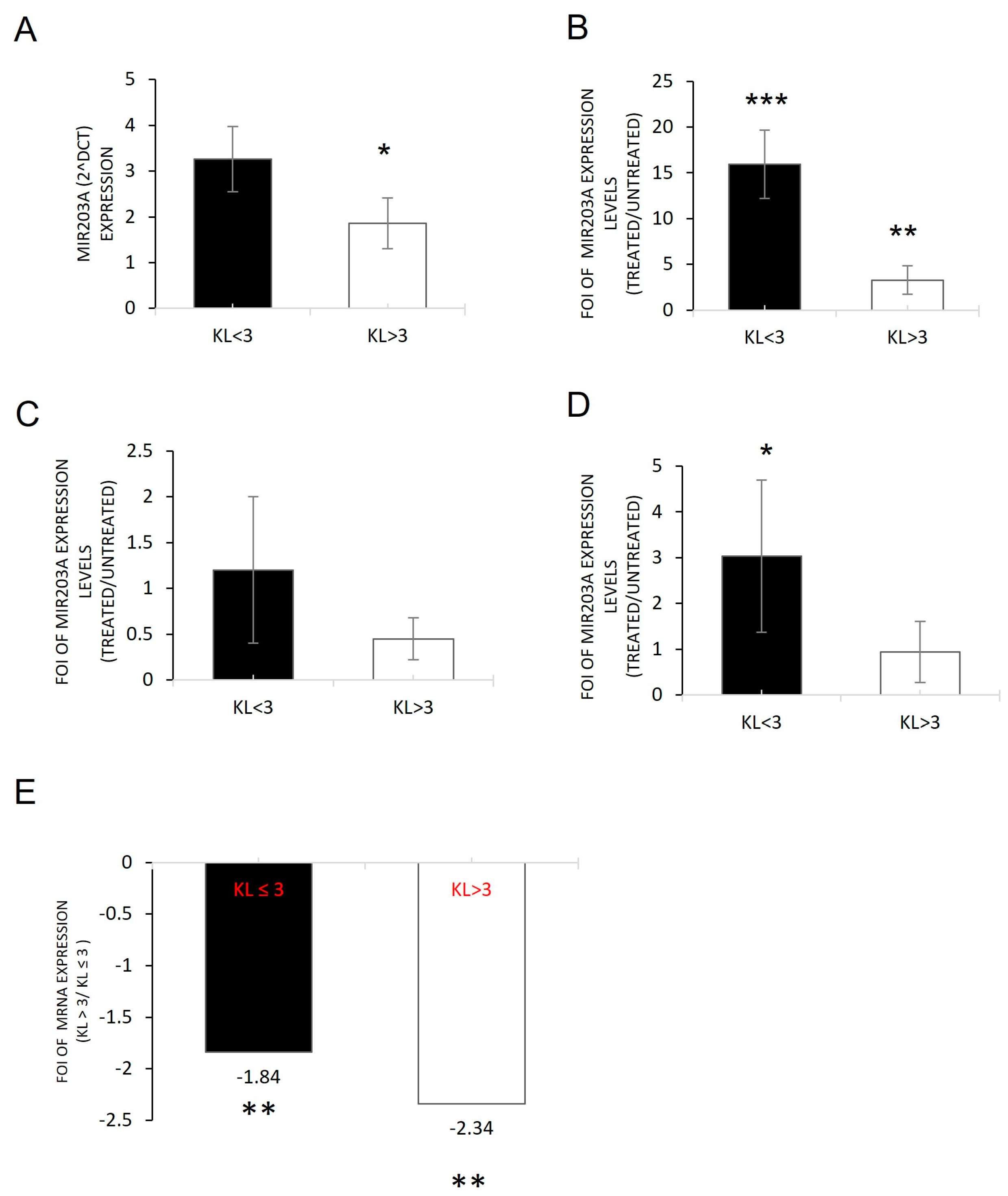
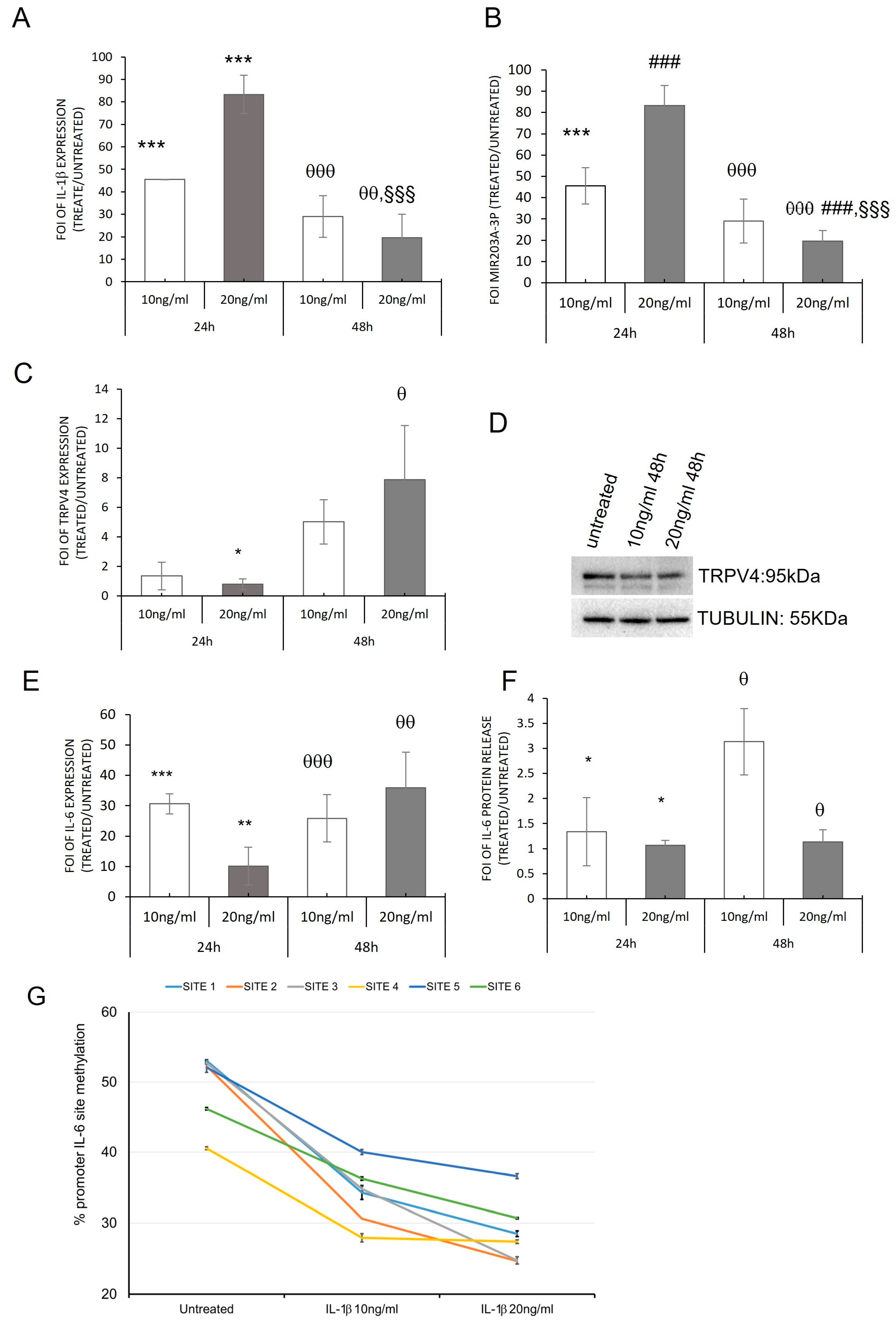
2.2. miRNAs Induced by Inflammatory Microenvironments on OB-OA-Derived Cells
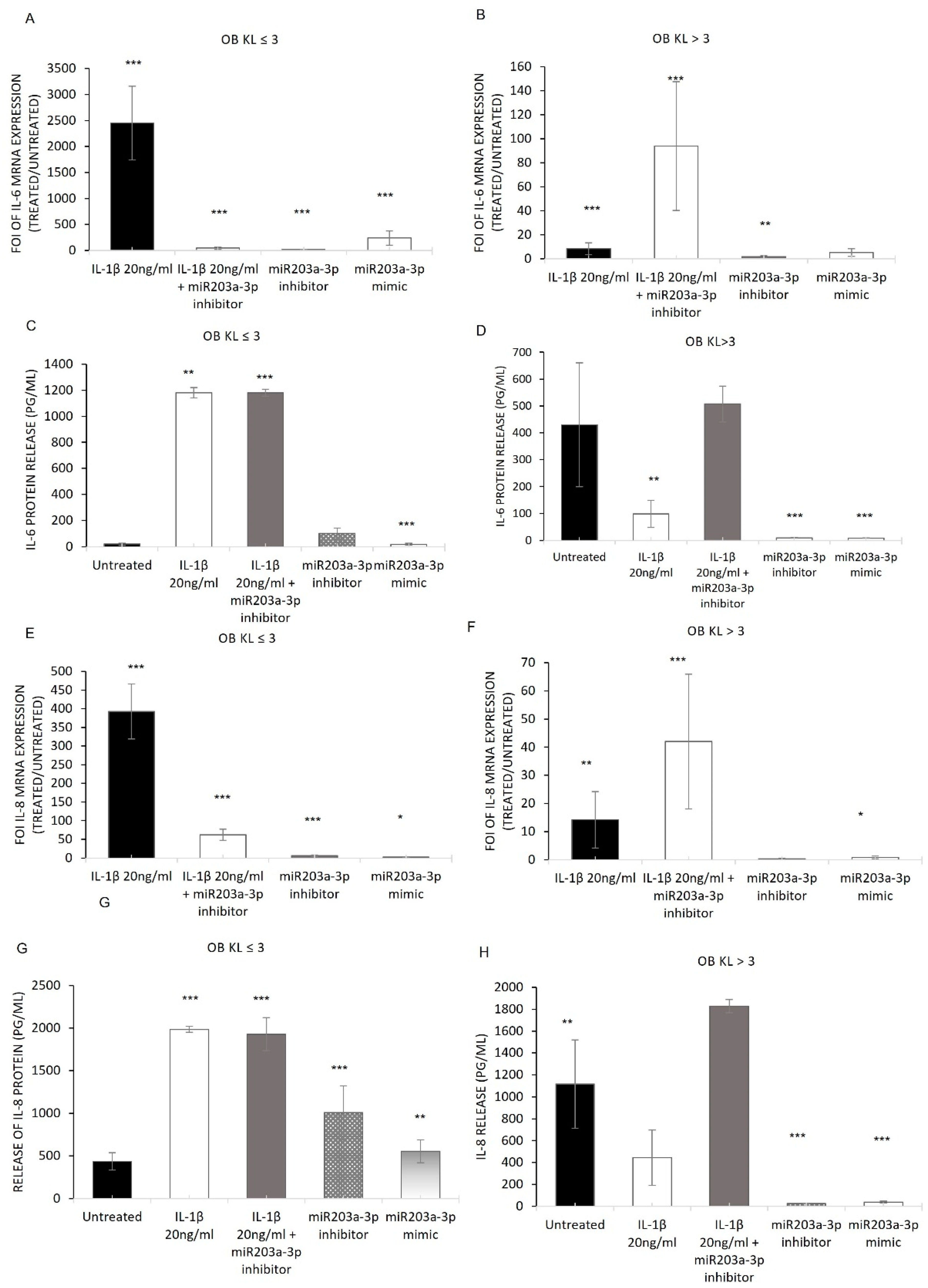
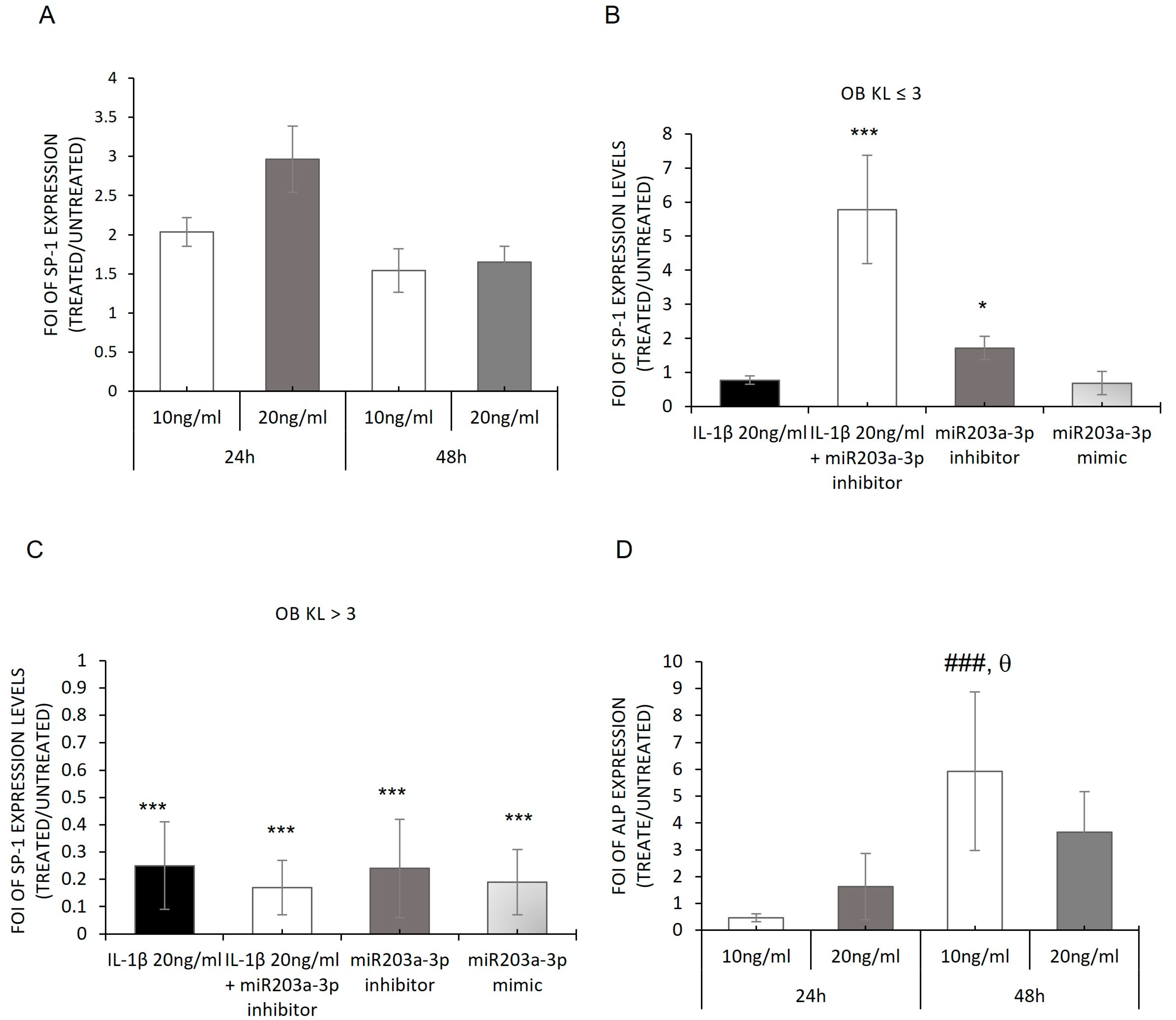
2.3. hMSC Evaluation of IL-1β Stimulation: In Vitro Model of OA Disease
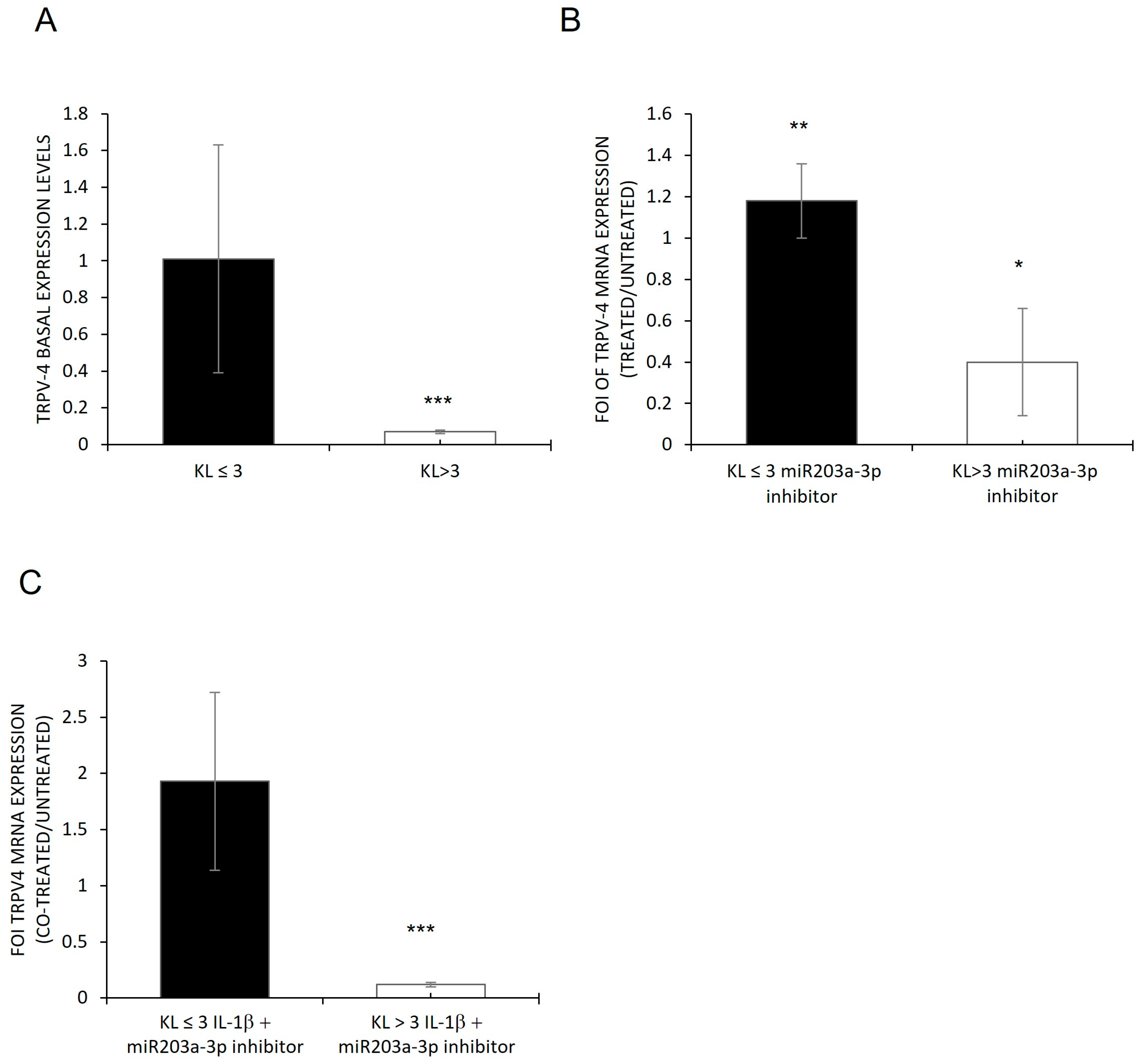
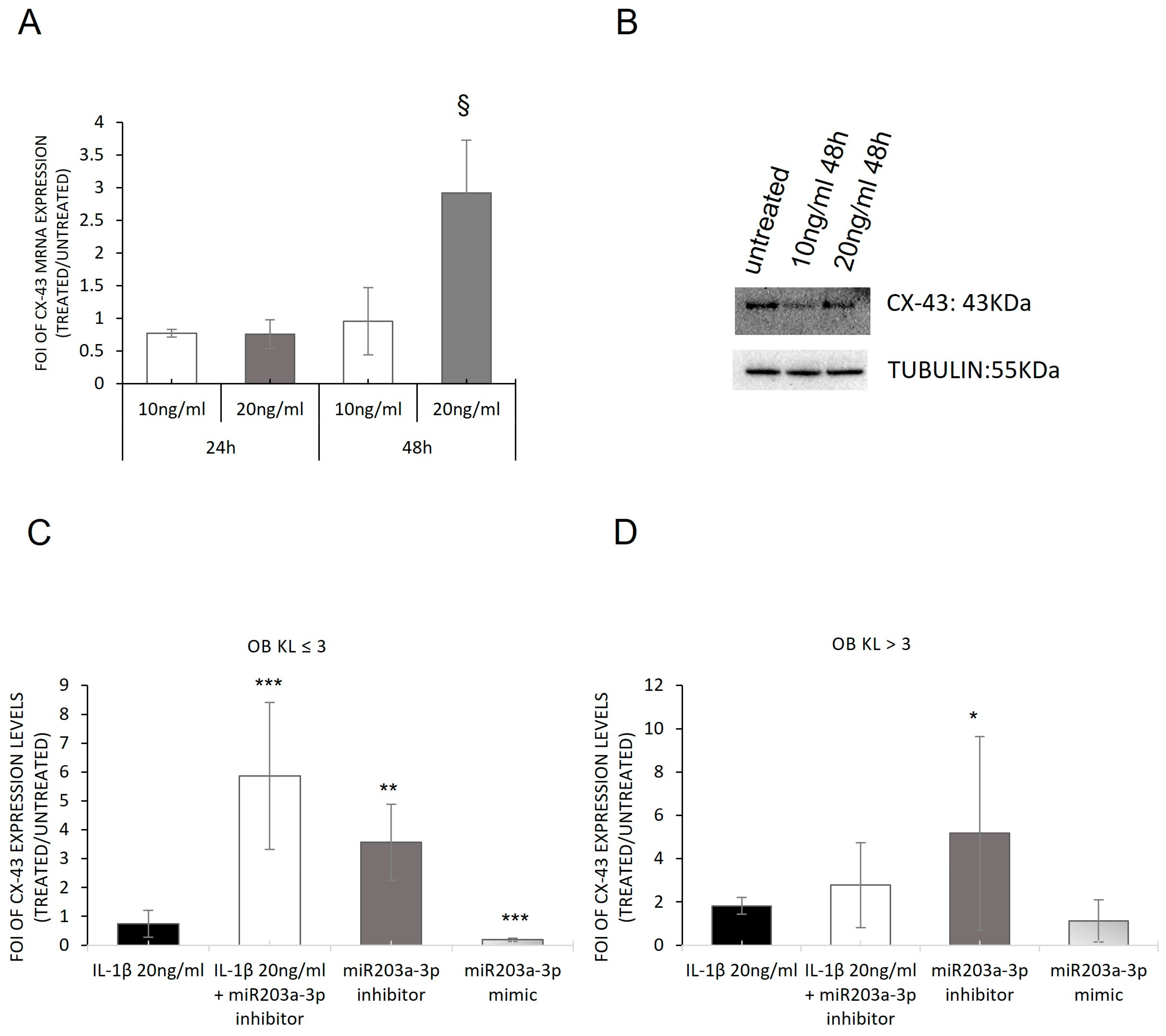
2.4. Involvement of IL1-β and miR203a-3p in CX-43 Expression
2.5. Modulation of SP-1 Expression in OB-Derived OA Patients
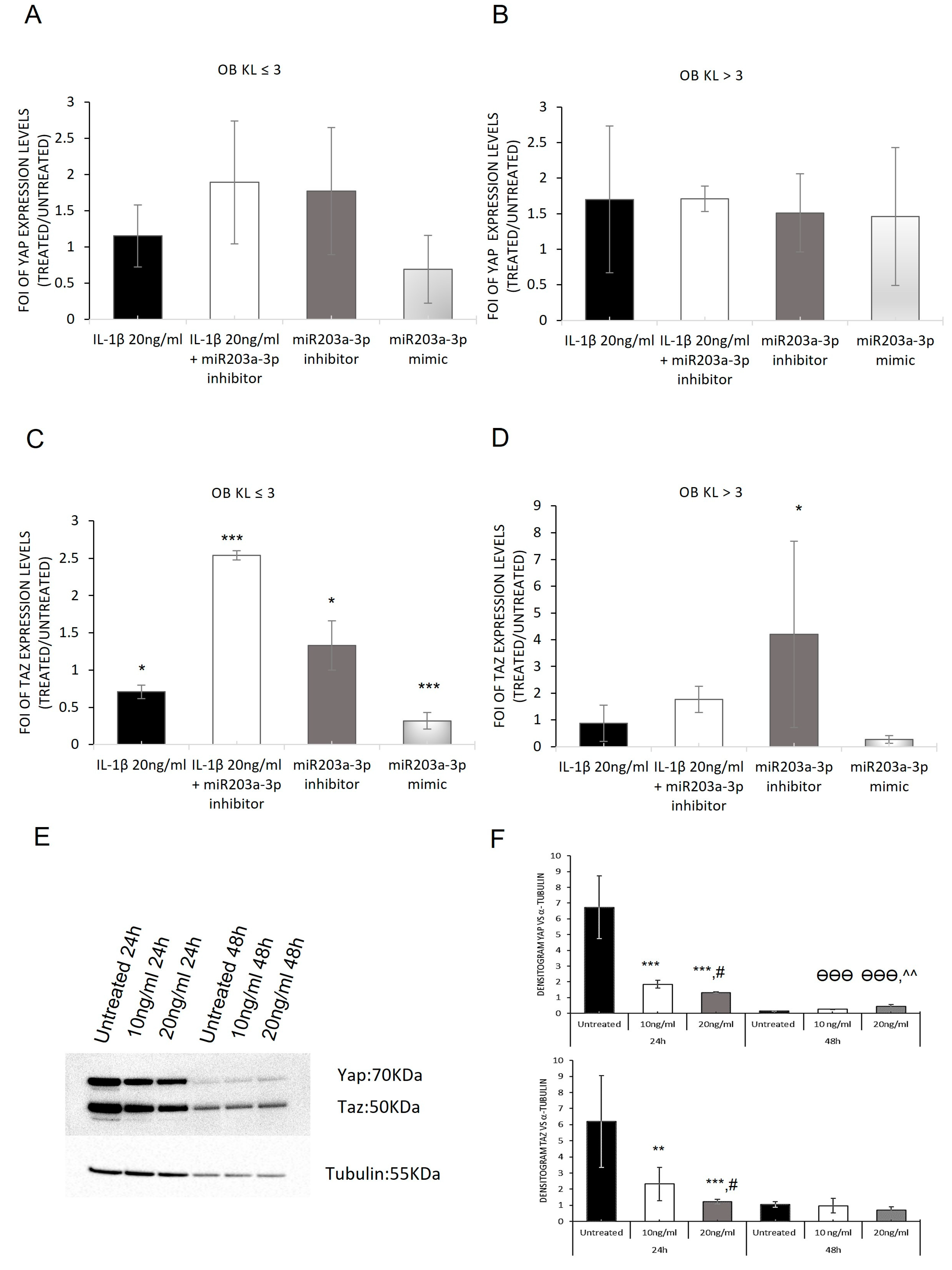
2.6. Modulation of YAP and TAZ Complex by IL-1β Stimulation
3. Discussion
4. Material and Methods
4.1. Cell Cultures and Reagents
4.2. Interleukin 1 Beta (IL-1β) Cell Treatments
4.3. Cell Transfection
4.4. RNA Extraction and Real-Time PCR
4.5. ELISA Assay
4.6. Western Blot Analysis
4.7. MSRE PCR Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hunziker, E.B. Articular cartilage repair: Basic science and clinical progress. A review of the current status and prospects. Osteoarthr. Cartil. 2002, 10, 432–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitaloni, M.; Botto-van Bemden, A.; Sciortino Contreras, R.M.; Scotton, D.; Bibas, M.; Quintero, M.; Monfort, J.; Carné, X.; de Abajo, F.; Oswald, E.; et al. Global management of patients with knee osteoarthritis begins with quality of life assessment: A systematic review. BMC Musculoskelet Disord. 2019, 20, 493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelletier, J.P.; Martel-Pelletier, J.; Abramson, S.B. Osteoarthritis, an inflammatory disease: Potential implication for the selection of new therapeutic targets. Arthritis Rheum. 2001, 44, 1237–1247. [Google Scholar] [CrossRef]
- Hartley, A.; Gregson, C.L.; Paternoster, L.; Tobias, J.H. Osteoarthritis: Insights Offered by the Study of Bone Mass Genetics. Curr. Osteoporos. Rep. 2021, 19, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Runhaar, J.; Bierma-Zeinstra, S.M.A. The Challenges in the Primary Prevention of Osteoarthritis. Clin. Geriatr. Med. 2022, 38, 259–271. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Poulet, C.; Malaise, M.; Abak, A.; Mahmud Hussen, B.; Taheriazam, A.; Taheri, M.; Hallajnejad, M. The Emerging Role of Non-Coding RNAs in Osteoarthritis. Front. Immunol. 2021, 12, 773171. [Google Scholar] [CrossRef]
- Tong, L.; Yu, H.; Huang, X.; Shen, J.; Xiao, G.; Chen, L.; Wang, H.; Xing, L.; Chen, D. Current understanding of osteoarthritis pathogenesis and relevant new approaches. Bone Res. 2022, 10, 60. [Google Scholar] [CrossRef]
- Li, S.; Xie, F.; Shi, K.; Wang, J.; Cao, Y.; Li, Y. Gossypol ameliorates the IL-1β-induced apoptosis and inflammation in chondrocytes by suppressing the activation of TLR4/MyD88/NF-κB pathway via downregulating CX43. Tissue Cell 2021, 73, 101621. [Google Scholar] [CrossRef]
- David, F.; Farley, J.; Huang, H.; Lavoie, J.P.; Laverty, S. Cytokine and chemokine gene expression of IL-1beta stimulated equine articular chondrocytes. Vet. Surg. 2007, 36, 221–227. [Google Scholar] [CrossRef]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.P.; Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 33–42. [Google Scholar] [CrossRef]
- Goldring, M.B.; Birkhead, J.R.; Suen, L.F.; Yamin, R.; Mizuno, S.; Glowacki, J.; Arbiser, J.L.; Apperley, J.F. Interleukin-1 beta-modulated gene expression in immortalized human chondrocytes. J. Clin. Invest. 1994, 94, 2307–2316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Tian, L.; Du, X.; Deng, Z. MiR-203 regulates estrogen receptor α and cartilage degradation in IL-1β-stimulated chondrocytes. J. Bone Miner. Metab. 2020, 38, 346–356. [Google Scholar] [CrossRef]
- Meng, Y.; Hu, X.; Li, S.; Zeng, X.; Qiu, L.; Wei, M.; Wang, Z.; Han, J. miR-203 inhibits cell proliferation and ERK pathway in prostate cancer by targeting IRS-1. BMC Cancer 2020, 20, 1028. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Niger, C.; Buo, A.M.; Eidelman, E.R.; Chen, R.J.; Stains, J.P. Connexin43 enhances the expression of osteoarthritis-associated genes in synovial fibroblasts in culture. BMC Musculoskelet Disord. 2014, 15, 425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gago-Fuentes, R.; Fernández-Puente, P.; Megias, D.; Carpintero-Fernández, P.; Mateos, J.; Acea, B.; Fonseca, E.; Blanco, F.J.; Mayan, M.D. Proteomic Analysis of Connexin 43 Reveals Novel Interactors Related to Osteoarthritis. Mol. Cell Proteom. 2015, 14, 1831–1845. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Fu, Y.; Zhuang, Y.; Zhang, K.; Lu, D. miR-382-3p suppressed IL-1β induced inflammatory response of chondrocytes via the TLR4/MyD88/NF-κB signaling pathway by directly targeting CX43. J. Cell. Physiol. 2019, 234, 23160–23168. [Google Scholar] [CrossRef]
- Martins-Marques, T.; Costa, M.C.; Catarino, S.; Simoes, I.; Aasen, T.; Enguita, F.J.; Girao, H. Cx43-mediated sorting of miRNAs into extracellular vesicles. EMBO Rep. 2022, 23, e54312. [Google Scholar] [CrossRef] [PubMed]
- Varela-Eirin, M.; Loureiro, J.; Fonseca, E.; Corrochano, S.; Caeiro, J.R.; Collado, M.; Mayan, M.D. Cartilage regeneration and ageing: Targeting cellular plasticity in osteoarthritis. Ageing Res. Rev. 2018, 42, 56–71. [Google Scholar] [CrossRef]
- Mayan, M.D.; Carpintero-Fernandez, P.; Gago-Fuentes, R.; Martinez-de-Ilarduya, O.; Wang, H.Z.; Valiunas, V.; Brink, P.; Blanco, F.J. Human articular chondrocytes express multiple gap junction proteins: Differential expression of connexins in normal and osteoarthritic cartilage. Am. J. Pathol. 2013, 182, 1337–1346. [Google Scholar] [CrossRef] [Green Version]
- Costa, V.; De Fine, M.; Carina, V.; Conigliaro, A.; Raimondi, L.; De Luca, A.; Bellavia, D.; Salamanna, F.; Alessandro, R.; Pignatti, G.; et al. How miR-31-5p and miR-33a-5p Regulates SP1/CX43 Expression in Osteoarthritis Disease: Preliminary Insights. Int. J. Mol. Sci. 2021, 22, 2471. [Google Scholar] [CrossRef]
- Varela-Eirín, M.; Carpintero-Fernández, P.; Guitián-Caamaño, A.; Varela-Vázquez, A.; García-Yuste, A.; Sánchez-Temprano, A.; Bravo-López, S.B.; Yañez-Cabanas, J.; Fonseca, E.; Largo, R.; et al. Extracellular vesicles enriched in connexin 43 promote a senescent phenotype in bone and synovial cells contributing to osteoarthritis progression. Cell Death Dis. 2022, 13, 681. [Google Scholar] [CrossRef]
- Shorter, E.; Avelar, R.; Zachariou, M.; Spyrou, G.M.; Raina, P.; Smagul, A.; Ashraf Kharaz, Y.; Peffers, M.; Goljanek-Whysall, K.; de Magalhães, J.P.; et al. Identifying Novel Osteoarthritis-Associated Genes in Human Cartilage Using a Systematic Meta-Analysis and a Multi-Source Integrated Network. Int. J. Mol. Sci. 2022, 23, 4395. [Google Scholar] [CrossRef] [PubMed]
- Fakhry, M.; Hamade, E.; Badran, B.; Buchet, R.; Magne, D. Molecular mechanisms of mesenchymal stem cell differentiation towards osteoblasts. World J. Stem Cells 2013, 5, 136–148. [Google Scholar] [CrossRef]
- Costa, V.; Raimondi, L.; Conigliaro, A.; Salamanna, F.; Carina, V.; De Luca, A.; Bellavia, D.; Alessandro, R.; Fini, M.; Giavaresi, G. Hypoxia-inducible factor 1A may regulate the commitment of mesenchymal stromal cells toward angio-osteogenesis by mirna-675-5P. Cytotherapy 2017, 19, 1412–1425. [Google Scholar] [CrossRef] [PubMed]
- Costa, V.; Carina, V.; Conigliaro, A.; Raimondi, L.; De Luca, A.; Bellavia, D.; Salamanna, F.; Setti, S.; Alessandro, R.; Fini, M.; et al. miR-31-5p Is a LIPUS-Mechanosensitive MicroRNA that Targets HIF-1α Signaling and Cytoskeletal Proteins. Int. J. Mol. Sci. 2019, 20, 1569. [Google Scholar] [CrossRef] [Green Version]
- Costa, V.; Carina, V.; Raimondi, L.; De Luca, A.; Bellavia, D.; Conigliaro, A.; Salamanna, F.; Alessandro, R.; Fini, M.; Giavaresi, G. MiR-33a Controls hMSCS Osteoblast Commitment Modulating the Yap/Taz Expression Through EGFR Signaling Regulation. Cells 2019, 8, 1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, Y.; Zhang, L.; Zhang, X.; Ma, C.; Chen, Z. Expression of MiR-140 and MiR-199 in Synovia and its Correlation with the Progression of Knee Osteoarthritis. Med. Sci. Monit. 2020, 26, e918174. [Google Scholar] [CrossRef]
- Philipot, D.; Guérit, D.; Platano, D.; Chuchana, P.; Olivotto, E.; Espinoza, F.; Dorandeu, A.; Pers, Y.M.; Piette, J.; Borzi, R.M.; et al. p16INK4a and its regulator miR-24 link senescence and chondrocyte terminal differentiation-associated matrix remodeling in osteoarthritis. Arthritis Res. Ther. 2014, 16, R58. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Du, L.; Dong, Z.; Yang, Y.; Zhang, X.; Wang, L.; Li, J.; Zheng, G.; Qu, A.; Wang, C. MiR-203 suppresses ZNF217 upregulation in colorectal cancer and its oncogenicity. PLoS ONE 2015, 10, e0116170. [Google Scholar] [CrossRef]
- Zierau, O.; Helle, J.; Schadyew, S.; Morgenroth, Y.; Bentler, M.; Hennig, A.; Chittur, S.; Tenniswood, M.; Kretzschmar, G. Role of miR-203 in estrogen receptor-mediated signaling in the rat uterus and endometrial carcinoma. J. Cell. Biochem. 2018, 119, 5359–5372. [Google Scholar] [CrossRef]
- Fan, F.Y.; Deng, R.; Qiu, L.; Wen, Q.; Zeng, Y.; Gao, L.; Zhang, C.; Kong, P.; Zhong, J.; Zeng, N.; et al. miR-203a-3p.1 is involved in the regulation of osteogenic differentiation by directly targeting Smad9 in MM-MSCs. Oncol. Lett. 2019, 18, 6339–6346. [Google Scholar] [CrossRef] [Green Version]
- Van Pelt, D.W.; Vechetti, I.J.; Lawrence, M.M.; Van Pelt, K.L.; Patel, P.; Miller, B.F.; Butterfield, T.A.; Dupont-Versteegden, E.E. Serum extracellular vesicle miR-203a-3p content is associated with skeletal muscle mass and protein turnover during disuse atrophy and regrowth. Am. J. Physiol. Cell Physiol. 2020, 319, C419–C431. [Google Scholar] [CrossRef]
- Stanczyk, J.; Ospelt, C.; Karouzakis, E.; Filer, A.; Raza, K.; Kolling, C.; Gay, R.; Buckley, C.D.; Tak, P.P.; Gay, S.; et al. Altered expression of microRNA-203 in rheumatoid arthritis synovial fibroblasts and its role in fibroblast activation. Arthritis Rheum. 2011, 63, 373–381. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Wu, H.; Ye, Y.; Li, Z.; Hao, S.; Kong, L.; Zheng, X.; Lin, S.; Nie, Q.; Zhang, X. The transient expression of miR-203 and its inhibiting effects on skeletal muscle cell proliferation and differentiation. Cell Death Dis. 2014, 5, e1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, L.; Liu, D.; Li, C.G.; Wang, Y.J. MiR-203 is essential for the shift from osteogenic differentiation to adipogenic differentiation of mesenchymal stem cells in postmenopausal osteoporosis. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5804–5814. [Google Scholar] [CrossRef]
- Maneix, L.; Servent, A.; Porée, B.; Ollitrault, D.; Branly, T.; Bigot, N.; Boujrad, N.; Flouriot, G.; Demoor, M.; Boumediene, K.; et al. Up-regulation of type II collagen gene by 17β-estradiol in articular chondrocytes involves Sp1/3, Sox-9, and estrogen receptor α. J. Mol. Med. 2014, 92, 1179–1200. [Google Scholar] [CrossRef]
- KELLGREN, J.H.; LAWRENCE, J.S. Radiological assessment of osteo-arthrosis. Ann. Rheum. Dis. 1957, 16, 494–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Luca, A.; Bellavia, D.; Raimondi, L.; Carina, V.; Costa, V.; Fini, M.; Giavaresi, G. Multiple Effects of Resveratrol on Osteosarcoma Cell Lines. Pharmaceuticals 2022, 15, 342. [Google Scholar] [CrossRef]
- Bellavia, D.; Costa, V.; De Luca, A.; Cordaro, A.; Fini, M.; Giavaresi, G.; Caradonna, F.; Raimondi, L. The Binomial "Inflammation-Epigenetics" in Breast Cancer Progression and Bone Metastasis: IL-1β Actions Are Influenced by TET Inhibitor in MCF-7 Cell Line. Int. J. Mol. Sci. 2022, 23, 15422. [Google Scholar] [CrossRef]
- Iliopoulos, D.; Malizos, K.N.; Oikonomou, P.; Tsezou, A. Integrative microRNA and proteomic approaches identify novel osteoarthritis genes and their collaborative metabolic and inflammatory networks. PLoS ONE 2008, 3, e3740. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, X. Downregulated miR-203 attenuates IL-β, IL-6, and TNF-α activation in TRAF6-treated human renal mesangial and tubular epithelial cells. Int. J. Clin. Exp. Pathol. 2020, 13, 324–331. [Google Scholar] [PubMed]
- Ghafouri-Fard, S.; Abak, A.; Fattahi, F.; Hussen, B.M.; Bahroudi, Z.; Shoorei, H.; Taheri, M. The interaction between miRNAs/lncRNAs and nuclear factor-κB (NF-κB) in human disorders. Biomed. Pharmacother. 2021, 138, 111519. [Google Scholar] [CrossRef] [PubMed]
- McNulty, A.L.; Leddy, H.A.; Liedtke, W.; Guilak, F. TRPV4 as a therapeutic target for joint diseases. Naunyn Schmiedebergs Arch. Pharmacol. 2015, 388, 437–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, F.; Zhu, W.; Wang, L. MicroRNA-203 up-regulates nitric oxide expression in temporomandibular joint chondrocytes via targeting TRPV4. Arch. Oral Biol. 2013, 58, 192–199. [Google Scholar] [CrossRef]
- Tang, C.; Liu, Q.; Zhang, Y.; Liu, G.; Shen, G. Identification of CIRBP and TRPV4 as Immune-Related Diagnostic Biomarkers in Osteoarthritis. Int. J. Gen. Med. 2021, 14, 10235–10245. [Google Scholar] [CrossRef]
- Varela-Eirín, M.; Varela-Vázquez, A.; Guitián-Caamaño, A.; Paíno, C.L.; Mato, V.; Largo, R.; Aasen, T.; Tabernero, A.; Fonseca, E.; Kandouz, M.; et al. Targeting of chondrocyte plasticity via connexin43 modulation attenuates cellular senescence and fosters a pro-regenerative environment in osteoarthritis. Cell Death Dis. 2018, 9, 1166. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Ren, J.; Sun, Y.; Xue, Y.; Zhang, Z.; Gong, A.; Wang, B.; Zhong, Z.; Cui, Z.; Xi, Z.; et al. A connexin43/YAP axis regulates astroglial-mesenchymal transition in hemoglobin induced astrocyte activation. Cell Death Differ. 2018, 25, 1870–1884. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Cao, X.; Li, W.; Liu, P.; Zhao, Y.; Song, L.; Chen, J.; Chen, B.; Yu, W.; Xu, Y. Targeting connexin 43 provides anti-inflammatory effects after intracerebral hemorrhage injury by regulating YAP signaling. J. Neuroinflammation 2020, 17, 322. [Google Scholar] [CrossRef]
- Baglìo, S.R.; Devescovi, V.; Granchi, D.; Baldini, N. MicroRNA expression profiling of human bone marrow mesenchymal stem cells during osteogenic differentiation reveals Osterix regulation by miR-31. Gene 2013, 527, 321–331. [Google Scholar] [CrossRef]
- Finch-Edmondson, M.L.; Strauss, R.P.; Passman, A.M.; Sudol, M.; Yeoh, G.C.; Callus, B.A. TAZ Protein Accumulation Is Negatively Regulated by YAP Abundance in Mammalian Cells. J. Biol. Chem. 2015, 290, 27928–27938. [Google Scholar] [CrossRef] [Green Version]
- Raimondi, L.; De Luca, A.; Costa, V.; Amodio, N.; Carina, V.; Bellavia, D.; Tassone, P.; Pagani, S.; Fini, M.; Alessandro, R.; et al. Circulating biomarkers in osteosarcoma: New translational tools for diagnosis and treatment. Oncotarget 2017, 8, 100831–100851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellavia, D.; Veronesi, F.; Carina, V.; Costa, V.; Raimondi, L.; De Luca, A.; Alessandro, R.; Fini, M.; Giavaresi, G. Gene therapy for chondral and osteochondral regeneration: Is the future now? Cell Mol. Life Sci. 2018, 75, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Raimondo, S.; Urzì, O.; Conigliaro, A.; Bosco, G.L.; Parisi, S.; Carlisi, M.; Siragusa, S.; Raimondi, L.; Luca, A.; Giavaresi, G.; et al. Extracellular Vesicle microRNAs Contribute to the Osteogenic Inhibition of Mesenchymal Stem Cells in Multiple Myeloma. Cancers 2020, 12, 449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amodio, N.; Raimondi, L.; Juli, G.; Stamato, M.A.; Caracciolo, D.; Tagliaferri, P.; Tassone, P. MALAT1: A druggable long non-coding RNA for targeted anti-cancer approaches. J. Hematol. Oncol. 2018, 11, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Zeng, L.; Zhang, T.; Liu, J.; Wang, W. Tenuigenin Prevents IL-1β-induced Inflammation in Human Osteoarthritis Chondrocytes by Suppressing PI3K/AKT/NF-κB Signaling Pathway. Inflammation 2016, 39, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yang, L.; Wan, G.; Zhang, X.; Wang, Y.; Zhao, G. Serum miR-377 Can Be Used as a Diagnostic Marker for Acute Coronary Syndrome and Can Regulate Proinflammatory Factors and Endothelial Injury Markers. Acta Med. Okayama 2022, 76, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Lohcharoenkal, W.; Das Mahapatra, K.; Pasquali, L.; Crudden, C.; Kular, L.; Akkaya Ulum, Y.Z.; Zhang, L.; Xu Landén, N.; Girnita, L.; Jagodic, M.; et al. Genome-Wide Screen for MicroRNAs Reveals a Role for miR-203 in Melanoma Metastasis. J. Invest. Dermatol. 2018, 138, 882–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briem, E.; Budkova, Z.; Sigurdardottir, A.K.; Hilmarsdottir, B.; Kricker, J.; Timp, W.; Magnusson, M.K.; Traustadottir, G.A.; Gudjonsson, T. MiR-203a is differentially expressed during branching morphogenesis and EMT in breast progenitor cells and is a repressor of peroxidasin. Mech. Dev. 2019, 155, 34–47. [Google Scholar] [CrossRef]
- Cai, K.T.; Feng, C.X.; Zhao, J.C.; He, R.Q.; Ma, J.; Zhong, J.C. Upregulated miR-203a-3p and its potential molecular mechanism in breast cancer: A study based on bioinformatics analyses and a comprehensive meta-analysis. Mol. Med. Rep. 2018, 18, 4994–5008. [Google Scholar] [CrossRef] [Green Version]
- Dusílková, N.; Bašová, P.; Polívka, J.; Kodet, O.; Kulvait, V.; Pešta, M.; Trněný, M.; Stopka, T. Plasma miR-155, miR-203, and miR-205 are Biomarkers for Monitoring of Primary Cutaneous T-Cell Lymphomas. Int. J. Mol. Sci. 2017, 18, 2136. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Bai, B.; Wang, J.; Xu, Z.; Yan, S.; Liu, G. Identification of key mRNAs and microRNAs in the pathogenesis and progression of osteoarthritis using microarray analysis. Mol. Med. Rep. 2017, 16, 5659–5666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Yang, L.; Wang, X. Reduced circulating microRNA-203 predicts poor prognosis for glioblastoma. Cancer Biomark. 2017, 20, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Stains, J.P.; Lecanda, F.; Screen, J.; Towler, D.A.; Civitelli, R. Gap junctional communication modulates gene transcription by altering the recruitment of Sp1 and Sp3 to connexin-response elements in osteoblast promoters. J. Biol. Chem. 2003, 278, 24377–24387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strippoli, R.; Sandoval, P.; Moreno-Vicente, R.; Rossi, L.; Battistelli, C.; Terri, M.; Pascual-Antón, L.; Loureiro, M.; Matteini, F.; Calvo, E.; et al. Caveolin1 and YAP drive mechanically induced mesothelial to mesenchymal transition and fibrosis. Cell Death Dis. 2020, 11, 647. [Google Scholar] [CrossRef]
- Liu, W.; Dong, Z.; Liang, J.; Guo, X.; Guo, Y.; Shen, S.; Kuang, G.; Guo, W. Downregulation of Potential Tumor Suppressor miR-203a by Promoter Methylation Contributes to the Invasiveness of Gastric Cardia Adenocarcinoma. Cancer Invest. 2016, 34, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Dong, Z.; Liang, J.; Guo, Y.; Guo, X.; Shen, S.; Kuang, G.; Guo, W. Methylation-mediated repression of potential tumor suppressor miR-203a and miR-203b contributes to esophageal squamous cell carcinoma development. Tumour Biol. 2016, 37, 5621–5632. [Google Scholar] [CrossRef]
- Filippova, E.A.; Loginov, V.I.; Burdennyi, A.M.; Braga, E.A.; Pronina, I.V.; Kazubskaya, T.P.; Kushlinskii, D.N.; Utkin, D.O.; Fridman, M.V.; Khodyrev, D.S.; et al. Hypermethylated Genes of MicroRNA in Ovarian Carcinoma: Metastasis Prediction Marker Systems. Bull. Exp. Biol. Med. 2019, 167, 79–83. [Google Scholar] [CrossRef]
- Loginov, V.I.; Pronina, I.V.; Filippova, E.A.; Burdennyy, A.M.; Lukina, S.S.; Kazubskaya, T.P.; Uroshlev, L.A.; Fridman, M.V.; Brovkina, O.I.; Apanovich, N.V.; et al. Aberrant Methylation of 20 miRNA Genes Specifically Involved in Various Steps of Ovarian Carcinoma Spread: From Primary Tumors to Peritoneal Macroscopic Metastases. Int. J. Mol. Sci. 2022, 23, 1300. [Google Scholar] [CrossRef]
- Ali, A.; Mahla, S.B.; Reza, V.; Alireza, F. Predicting the possible effect of miR-203a-3p and miR-29a-3p on. J. Integr. Bioinform. 2021, 19, 20210016. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 31 October 2022).
- Caradonna, F.; Cruciata, I.; Schifano, I.; La Rosa, C.; Naselli, F.; Chiarelli, R.; Perrone, A.; Gentile, C. Methylation of cytokines gene promoters in IL-1β-treated human intestinal epithelial cells. Inflamm. Res. 2018, 67, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Bellavia, D.; Dimarco, E.; Caradonna, F. Characterization of three different clusters of 18S-26S ribosomal DNA genes in the sea urchin P. lividus: Genetic and epigenetic regulation synchronous to 5S rDNA. Gene 2016, 580, 118–124. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OA KL ≤ 3 (n = 4) | OA KL > 3 (n = 4) | |
|---|---|---|
| Age (yrs) | 60 ± 6 | 64 ± 11 |
| Weight (kg) | 66.8 ± 9.7 | 80.5 ± 24.3 |
| BMI (kg/m²) | 24.3 ± 2.8 | 27.1 ± 4.5 |
| Other pathologies | COPD Hashimoto thyroiditisBreast cancer | Hypertension Gastroesophageal relux |
| WBC (x10³/μL) | 5.89 ± 0.67 | 7.62 ± 1.01 |
| CRP (mg/L) | 0.20 ± 0.07 | 0.28 ± 0.11 |
| Gene | Primer Forward | Primer Reverse |
|---|---|---|
| SP-1 “Specific protein 1” | GCCTCCAGACCATTAACCTCAGT | GCTCCATGATCACCTGGGGCAT |
| CX-43 “Connexin43” | GAACTCAAGGTTGCCCAAAC | TTAGAGATGGTGCTTCCCG |
| Reference Gene | ||
| ACTB “Beta-Actin” | ATCAAGATCATTGCTCCTCCTGA | CTGCTTGCTGATCCACATCTG |
| Gene | Qiagen Number |
|---|---|
| IL-1β Hs_IL-1β_1_SG QuantiTect Primer Assay | QT00021385 |
| IL-6 Hs_IL-6_1_SG QuantiTect Primer Assay | QT00083720 |
| IL-8 Hs IL-8_1_SG QuantiTect Primer Assay | QT00083756 |
| ALPL Hs_ALPL_1_SG QuantiTect Primer Assay | QT00012957 |
| TRPV4 Hs_TRPV4_1_SG QuantiTect Primer Assay | QT00077217 |
| Reference Gene | |
| ACTB Hs_ACTB_1_SG QuantiTect Primer Assay | QT00095431 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, V.; De Fine, M.; Raimondi, L.; Bellavia, D.; Cordaro, A.; Carina, V.; Alessandro, R.; Pignatti, G.; Fini, M.; Giavaresi, G.; et al. Timing Expression of miR203a-3p during OA Disease: Preliminary In Vitro Evidence. Int. J. Mol. Sci. 2023, 24, 4316. https://doi.org/10.3390/ijms24054316
Costa V, De Fine M, Raimondi L, Bellavia D, Cordaro A, Carina V, Alessandro R, Pignatti G, Fini M, Giavaresi G, et al. Timing Expression of miR203a-3p during OA Disease: Preliminary In Vitro Evidence. International Journal of Molecular Sciences. 2023; 24(5):4316. https://doi.org/10.3390/ijms24054316
Chicago/Turabian StyleCosta, Viviana, Marcello De Fine, Lavinia Raimondi, Daniele Bellavia, Aurora Cordaro, Valeria Carina, Riccardo Alessandro, Giovanni Pignatti, Milena Fini, Gianluca Giavaresi, and et al. 2023. "Timing Expression of miR203a-3p during OA Disease: Preliminary In Vitro Evidence" International Journal of Molecular Sciences 24, no. 5: 4316. https://doi.org/10.3390/ijms24054316