
Inhibition of Myeloperoxidase Pro-Fibrotic Effect by Noscapine in Equine Endometrium
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Viability of Equine Endometrial Explants Data
2.2. The Independent Effect of MPO, NOSC, Time of Treatment and Estrous Cycle Phase, and Their Interaction Combinations
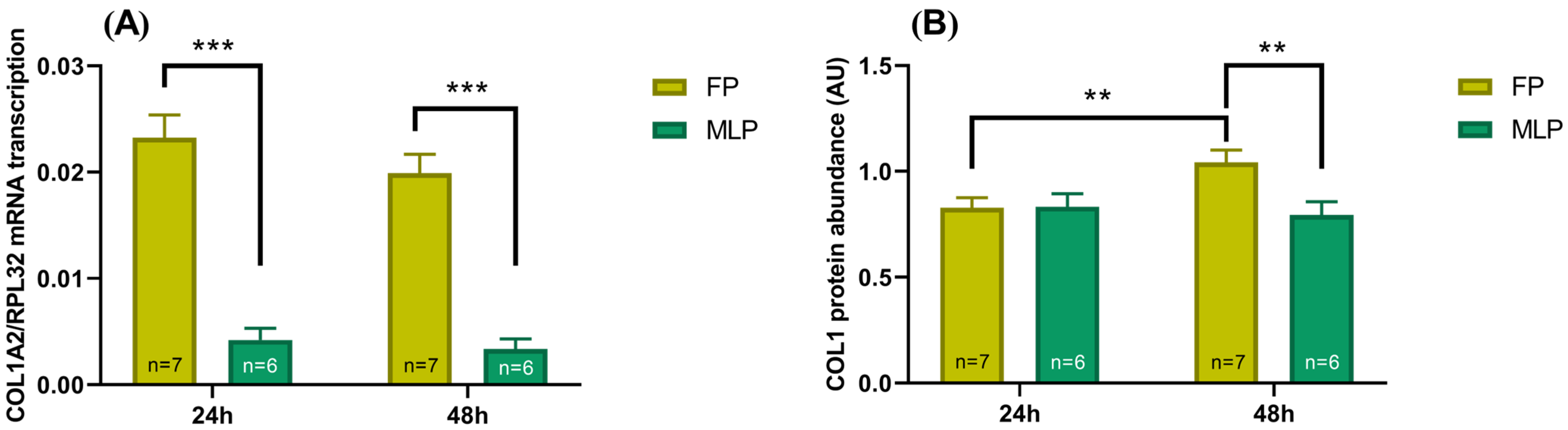
2.3. COL1 Expression Is Different, Depending on Time of Treatment and the Estrous Cycle Phase
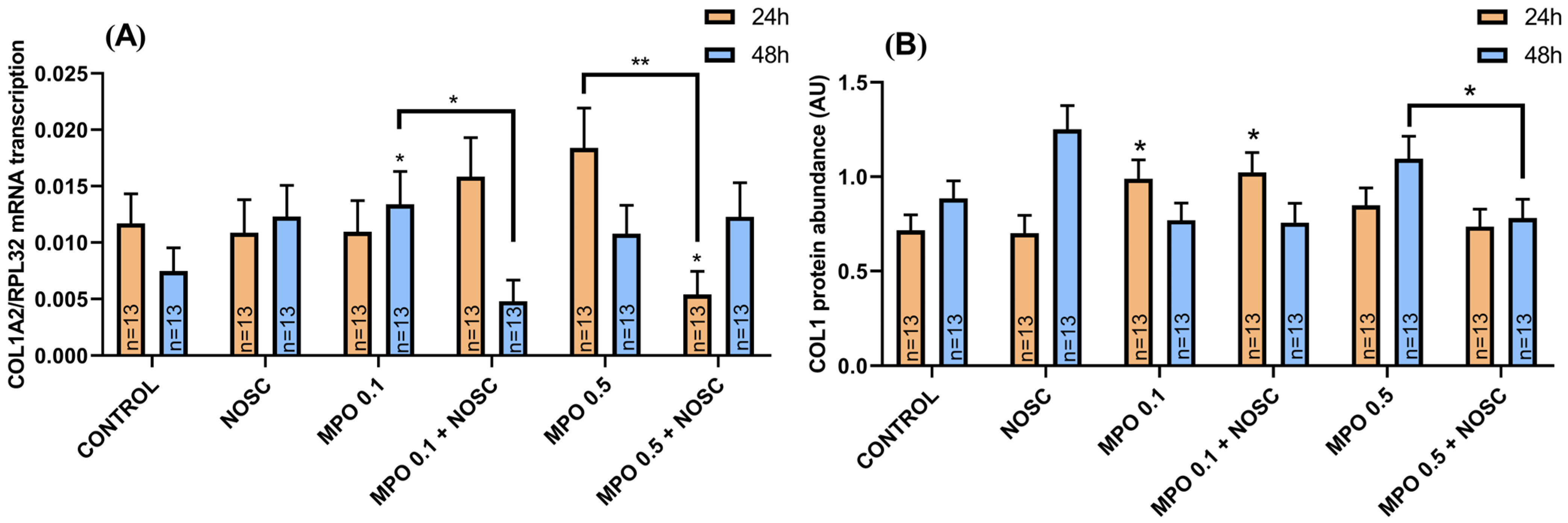
2.4. Noscapine Inhibition on MPO-Induced COL1 Expression Is Dependent on Time of Treatment
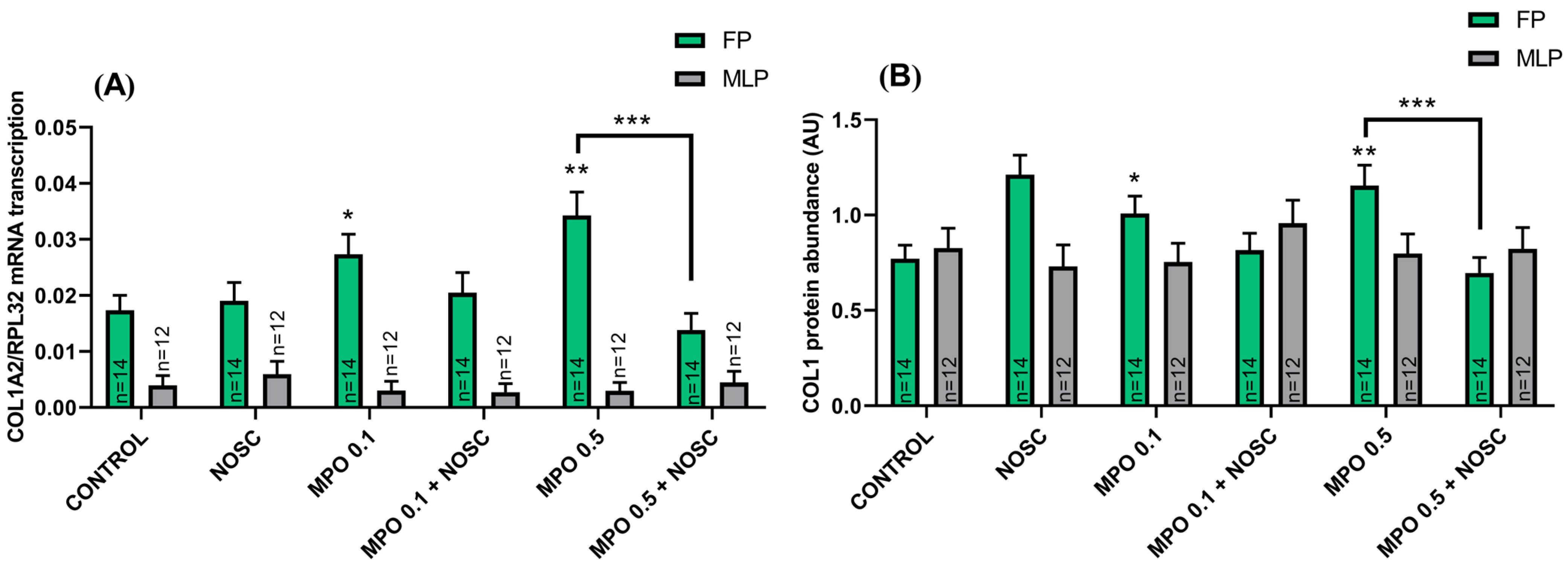
2.5. Estrous Cycle Phase Affects the Equine Endometrial Explant Response to MPO and NOSC Treatments
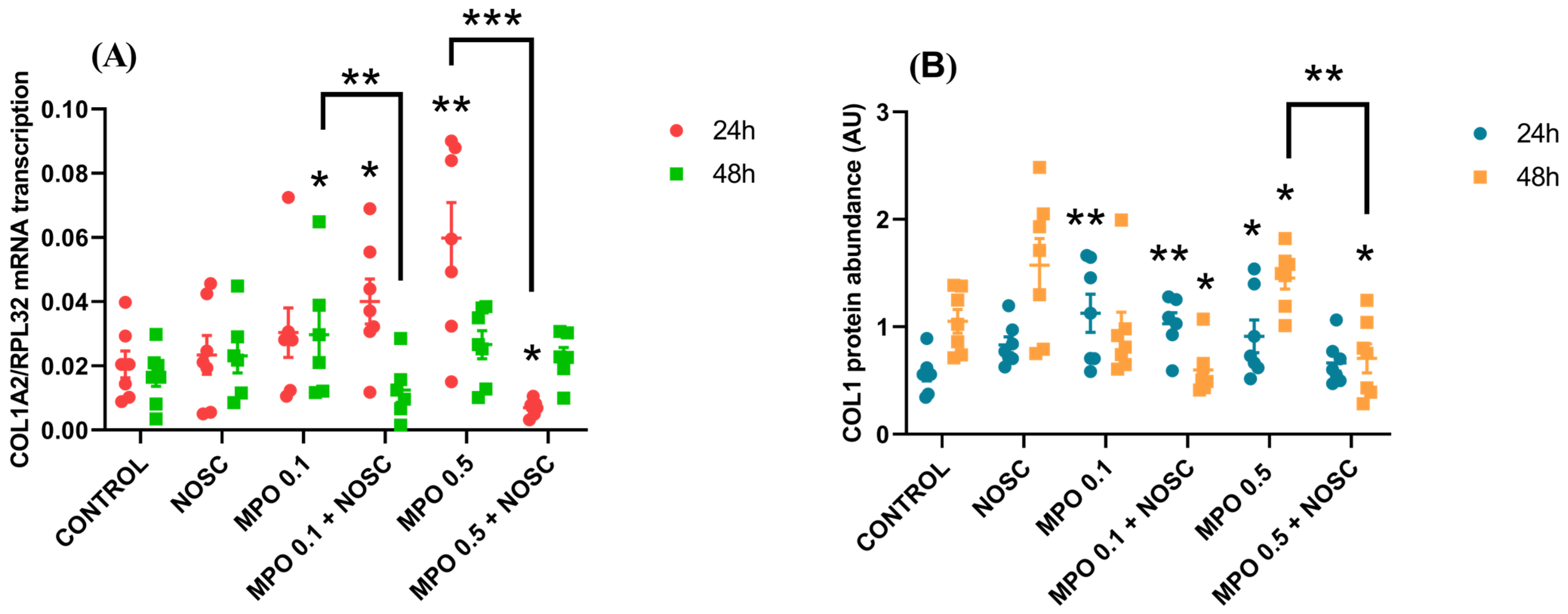
2.6. NOSC Inhibition of MPO-Induced COL1 Is Dependent on the Time of Treatment and Estrous Cycle Phase
3. Discussion
4. Materials and Methods
4.1. Mares Collection at Abattoir
4.2. Equine Endometrial Explant In Vitro Culture
4.3. Determination of Equine Endometrial Explant Viability
4.4. Real-Time Polymerase Chain Reaction (qPCR) was Used to Determine COL1A2 mRNA Transcription
4.5. Determination of COL1 Protein Relative Abundance by Western Blot Assay
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cowland, J.B.; Borregaard, N. Granulopoiesis and Granules of Human Neutrophils. Immunol. Rev. 2016, 273, 11–28. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- O’Donoghue, A.J.; Jin, Y.; Knudsen, G.M.; Perera, N.C.; Jenne, D.E.; Murphy, J.E.; Craik, C.S.; Hermiston, T.W. Global Substrate Profiling of Proteases in Human Neutrophil Extracellular Traps Reveals Consensus Motif Predominantly Contributed by Elastase. PLoS ONE 2013, 8, e75141. [Google Scholar] [CrossRef]
- Hoeksema, M.; van Eijk, M.; Haagsman, H.P.; Hartshorn, K.L. Histones as Mediators of Host Defense, Inflammation and Thrombosis. Future Microbiol. 2016, 11, 441–453. [Google Scholar] [CrossRef]
- Mangold, A.; Alias, S.; Scherz, T.; Hofbauer, T.M.; Jakowitsch, J.; Panzenböck, A.; Simon, D.; Laimer, D.; Bangert, C.; Kammerlander, A.; et al. Coronary Neutrophil Extracellular Trap Burden and Deoxyribonuclease Activity in ST-Elevation Acute Coronary Syndrome Are Predictors of ST-Segment Resolution and Infarct Size. Circ. Res. 2015, 116, 1182–1192. [Google Scholar] [CrossRef]
- Ribon, M.; Seninet, S.; Mussard, J.; Sebbag, M.; Clavel, C.; Serre, G.; Boissier, M.-C.; Semerano, L.; Decker, P. Neutrophil Extracellular Traps Exert Both Pro- and Anti-Inflammatory Actions in Rheumatoid Arthritis That Are Modulated by C1q and LL-37. J. Autoimmun. 2019, 98, 122–131. [Google Scholar] [CrossRef]
- Bonaventura, A.; Vecchié, A.; Abbate, A.; Montecucco, F. Neutrophil Extracellular Traps and Cardiovascular Diseases: An Update. Cells 2020, 9, 231. [Google Scholar] [CrossRef]
- Klopf, J.; Brostjan, C.; Eilenberg, W.; Neumayer, C. Neutrophil Extracellular Traps and Their Implications in Cardiovascular and Inflammatory Disease. Int. J. Mol. Sci. 2021, 22, 559. [Google Scholar] [CrossRef]
- Rebordão, M.R.; Amaral, A.; Lukasik, K.; Szóstek-Mioduchowska, A.; Pinto-Bravo, P.; Galvão, A.; Skarzynski, D.J.; Ferreira-Dias, G. Constituents of Neutrophil Extracellular Traps Induce In Vitro Collagen Formation in Mare Endometrium. Theriogenology 2018, 113, 8–18. [Google Scholar] [CrossRef]
- Amaral, A.; Fernandes, C.; Lukasik, K.; Szóstek-Mioduchowska, A.; Baclawska, A.; Rebordão, M.R.; Aguiar-Silva, J.; Pinto-Bravo, P.; Skarzynski, D.J.; Ferreira-Dias, G. Elastase Inhibition Affects Collagen Transcription and Prostaglandin Secretion in Mare Endometrium during the Estrous Cycle. Reprod. Domest. Anim. 2018, 53 (Suppl. S2), 66–69. [Google Scholar] [CrossRef]
- Amaral, A.; Fernandes, C.; Rebordão, M.R.; Szóstek-Mioduchowska, A.; Lukasik, K.; Gawronska-Kozak, B.; Telo da Gama, L.; Skarzynski, D.J.; Ferreira-Dias, G. The In Vitro Inhibitory Effect of Sivelestat on Elastase Induced Collagen and Metallopeptidase Expression in Equine Endometrium. Animals 2020, 10, 863. [Google Scholar] [CrossRef]
- Amaral, A.; Fernandes, C.; Morazzo, S.; Rebordão, M.R.; Szóstek-Mioduchowska, A.; Lukasik, K.; Gawronska-Kozak, B.; Telo da Gama, L.; Skarzynski, D.J.; Ferreira-Dias, G. The Inhibition of Cathepsin G on Endometrial Explants With Endometrosis in the Mare. Front Vet. Sci. 2020, 7, 582211. [Google Scholar] [CrossRef]
- Amaral, A.; Fernandes, C.; Rebordão, M.R.; Szóstek-Mioduchowska, A.; Lukasik, K.; Pinto-Bravo, P.; Telo da Gama, L.; Jan Skarzynski, D.; Ferreira-Dias, G. Myeloperoxidase Inhibition Decreases the Expression of Collagen and Metallopeptidase in Mare Endometria under In Vitro Conditions. Animals 2021, 11, 208. [Google Scholar] [CrossRef] [PubMed]
- Rada, B. Neutrophil Extracellular Traps Are Present in the Airways of ENaC-Overexpressing Mice with Cystic Fibrosis-like Lung Disease. BMC Immunol. 2021, 22, 7. [Google Scholar] [CrossRef]
- Pandolfi, L.; Bozzini, S.; Frangipane, V.; Percivalle, E.; De Luigi, A.; Violatto, M.B.; Lopez, G.; Gabanti, E.; Carsana, L.; D’Amato, M.; et al. Neutrophil Extracellular Traps Induce the Epithelial-Mesenchymal Transition: Implications in Post-COVID-19 Fibrosis. Front. Immunol. 2021, 12, 663303. [Google Scholar] [CrossRef]
- Urban, C.F.; Ermert, D.; Schmid, M.; Abu-Abed, U.; Goosmann, C.; Nacken, W.; Brinkmann, V.; Jungblut, P.R.; Zychlinsky, A. Neutrophil Extracellular Traps Contain Calprotectin, a Cytosolic Protein Complex Involved in Host Defense against Candida Albicans. PLoS Pathog. 2009, 5, e1000639. [Google Scholar] [CrossRef]
- Brinkmann, V. Neutrophil Extracellular Traps in the Second Decade. J. Innate. Immun. 2018, 10, 414–421. [Google Scholar] [CrossRef]
- Aratani, Y. Myeloperoxidase: Its Role for Host Defense, Inflammation, and Neutrophil Function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef]
- Klebanoff, S.J. Myeloperoxidase: Friend and Foe. J. Leukoc. Biol. 2005, 77, 598–625. [Google Scholar] [CrossRef]
- Nauseef, W.M. Myeloperoxidase in Human Neutrophil Host Defence. Cell Microbiol. 2014, 16, 1146–1155. [Google Scholar] [CrossRef]
- Sagel, S.D.; Wagner, B.D.; Anthony, M.M.; Emmett, P.; Zemanick, E.T. Sputum Biomarkers of Inflammation and Lung Function Decline in Children with Cystic Fibrosis. Am. J. Respir. Crit. Care Med. 2012, 186, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Sly, P.D.; Gangell, C.L.; Chen, L.; Ware, R.S.; Ranganathan, S.; Mott, L.S.; Murray, C.P.; Stick, S.M.; AREST CF Investigators. Risk Factors for Bronchiectasis in Children with Cystic Fibrosis. N. Engl. J. Med. 2013, 368, 1963–1970. [Google Scholar] [CrossRef]
- Pulli, B.; Ali, M.; Iwamoto, Y.; Zeller, M.W.G.; Schob, S.; Linnoila, J.J.; Chen, J.W. Myeloperoxidase-Hepatocyte-Stellate Cell Cross Talk Promotes Hepatocyte Injury and Fibrosis in Experimental Nonalcoholic Steatohepatitis. Antioxid. Redox. Signal. 2015, 23, 1255–1269. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; Alsahli, M.A.; Rahmani, A.H. Myeloperoxidase as an Active Disease Biomarker: Recent Biochemical and Pathological Perspectives. Med. Sci. 2018, 6, 33. [Google Scholar] [CrossRef]
- Kenney, R.M. The aetiology, diagnosis, and classification of chronic degenerative endometritis. In Workshop on Equine Endometritis; Hughes, J.P., Ed.; Newmarket Press: New York, NY, USA, 1992; Volume 125, p. 186. [Google Scholar]
- Hoffmann, C.; Ellenberger, C.; Mattos, R.C.; Aupperle, H.; Dhein, S.; Stief, B.; Schoon, H.-A. The Equine Endometrosis: New Insights into the Pathogenesis. Anim. Reprod. Sci. 2009, 111, 261–278. [Google Scholar] [CrossRef]
- Troedsson, M.H.; Liu, I.K.; Thurmond, M. Function of Uterine and Blood-Derived Polymorphonuclear Neutrophils in Mares Susceptible and Resistant to Chronic Uterine Infection: Phagocytosis and Chemotaxis. Biol. Reprod. 1993, 49, 507–514. [Google Scholar] [CrossRef]
- Troedsson, M.H.T. Breeding-Induced Endometritis in Mares. Vet. Clin. North Am. Equine. Pract. 2006, 22, 705–712. [Google Scholar] [CrossRef]
- Kotilainen, T.; Huhtinen, M.; Katila, T. Sperm-Induced Leukocytosis in the Equine Uterus. Theriogenology 1994, 41, 629–636. [Google Scholar] [CrossRef]
- Katila, T. Onset and Duration of Uterine Inflammatory Response of Mares after Insemination with Fresh Semen. Biol. Reprod. 1995, 52, 515–517. [Google Scholar] [CrossRef]
- Amaral, A.; Fernandes, C.; Szóstek-Mioduchowska, A.; Rebordão, M.R.; Skarzynski, D.J.; Ferreira-Dias, G. Noscapine Acts as a Protease Inhibitor of In Vitro Elastase-Induced Collagen Deposition in Equine Endometrium. Int. J. Mol. Sci. 2021, 22, 5333. [Google Scholar] [CrossRef]
- Amaral, A.; Fernandes, C.; Szóstek-Mioduchowska, A.; Lukasik, K.; Rebordão, M.R.; Pinto-Bravo, P.; Skarzynski, D.J.; Ferreira-Dias, G. The Inhibitory Effect of Noscapine on the In Vitro Cathepsin G-Induced Collagen Expression in Equine Endometrium. Life 2021, 11, 1107. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, M.O.; Dahlström, B.; Eckernäs, S.A.; Johansson, M.; Alm, A.T. Pharmacokinetics of Oral Noscapine. Eur. J. Clin. Pharmacol. 1990, 39, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Rida, P.C.G.; LiVecche, D.; Ogden, A.; Zhou, J.; Aneja, R. The Noscapine Chronicle: A Pharmaco-Historic Biography of the Opiate Alkaloid Family and Its Clinical Applications. Med. Res. Rev. 2015, 35, 1072–1096. [Google Scholar] [CrossRef] [PubMed]
- Rahmanian-Devin, P.; Baradaran Rahimi, V.; Jaafari, M.R.; Golmohammadzadeh, S.; Sanei-Far, Z.; Askari, V.R. Noscapine, an Emerging Medication for Different Diseases: A Mechanistic Review. Evid. Based. Complement Altern. Med. 2021, 2021, 8402517. [Google Scholar] [CrossRef] [PubMed]
- Quisbert-Valenzuela, E.O.; Calaf, G.M. Apoptotic Effect of Noscapine in Breast Cancer Cell Lines. Int. J. Oncol. 2016, 48, 2666–2674. [Google Scholar] [CrossRef] [PubMed]
- Esnaashari, S.S.; Muhammadnejad, S.; Amanpour, S.; Amani, A. A Combinational Approach Towards Treatment of Breast Cancer: An Analysis of Noscapine-Loaded Polymeric Nanoparticles and Doxorubicin. AAPS PharmSciTech 2020, 21, 166. [Google Scholar] [CrossRef]
- Kach, J.; Sandbo, N.; La, J.; Denner, D.; Reed, E.B.; Akimova, O.; Koltsova, S.; Orlov, S.N.; Dulin, N.O. Antifibrotic Effects of Noscapine through Activation of Prostaglandin E2 Receptors and Protein Kinase A. J. Biol. Chem. 2014, 289, 7505–7513. [Google Scholar] [CrossRef]
- Doddapaneni, R.; Patel, K.; Chowdhury, N.; Singh, M. Noscapine Chemosensitization Enhances Docetaxel Anticancer Activity and Nanocarrier Uptake in Triple Negative Breast Cancer. Exp. Cell Res. 2016, 346, 65–73. [Google Scholar] [CrossRef]
- Rebordão, M.R.; Carneiro, C.; Alexandre-Pires, G.; Brito, P.; Pereira, C.; Nunes, T.; Galvão, A.; Leitão, A.; Vilela, C.; Ferreira-Dias, G. Neutrophil Extracellular Traps Formation by Bacteria Causing Endometritis in the Mare. J. Reprod. Immunol. 2014, 106, 41–49. [Google Scholar] [CrossRef]
- Aurich, C. Reproductive Cycles of Horses. Anim. Reprod. Sci. 2011, 124, 220–228. [Google Scholar] [CrossRef]
- Koop, A.C.; Thiele, N.D.; Steins, D.; Michaëlsson, E.; Wehmeyer, M.; Scheja, L.; Steglich, B.; Huber, S.; Schulze Zur Wiesch, J.; Lohse, A.W.; et al. Therapeutic Targeting of Myeloperoxidase Attenuates NASH in Mice. Hepatol. Commun. 2020, 4, 1441–1458. [Google Scholar] [CrossRef]
- Chandler, J.D.; Margaroli, C.; Horati, H.; Kilgore, M.B.; Veltman, M.; Liu, H.K.; Taurone, A.J.; Peng, L.; Guglani, L.; Uppal, K.; et al. Myeloperoxidase Oxidation of Methionine Associates with Early Cystic Fibrosis Lung Disease. Eur. Respir. J. 2018, 52, 1801118. [Google Scholar] [CrossRef]
- Kawadkar, M.; Mandloi, A.S.; Saxena, V.; Tamadaddi, C.; Sahi, C.; Dhote, V.V. Noscapine Alleviates Cerebral Damage in Ischemia-Reperfusion Injury in Rats. Naunyn. Schmiedebergs Arch. Pharm. 2021, 394, 669–683. [Google Scholar] [CrossRef]
- Frangie, C.; Daher, J. Role of Myeloperoxidase in Inflammation and Atherosclerosis (Review). Biomed. Rep. 2022, 16, 53. [Google Scholar] [CrossRef]
- Awasthi, A.; Kumar, N.; Mishra, A.; Ravi, R.; Dalal, A.; Shankar, S.; Chandra, R. Noscapine–Amino Acid Conjugates Suppress the Progression of Cancer Cells. ACS Pharmacol. Transl. Sci. 2022, 5, 1292–1304. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.; Marcotte, E.M. Insights into the Regulation of Protein Abundance from Proteomic and Transcriptomic Analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, R.I. Collagen I and the Fibroblast: High Protein Expression Requires a New Paradigm of Post-Transcriptional, Feedback Regulation. Biochem. Biophys. Rep. 2015, 3, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; Kettle, A.J.; Hampton, M.B. Reactive Oxygen Species and Neutrophil Function. Annu. Rev. Biochem. 2016, 85, 765–792. [Google Scholar] [CrossRef]
- Chelombitko, M.A. Role of Reactive Oxygen Species in Inflammation: A Minireview. Mosc. Univ. Biol. Sci. Bull. 2018, 73, 199–202. [Google Scholar] [CrossRef]
- Nambiar, N.; Nagireddy, P.K.R.; Pedapati, R.; Kantevari, S.; Lopus, M. Tubulin- and ROS-Dependent Antiproliferative Mechanism of a Potent Analogue of Noscapine, N-Propargyl Noscapine. Life Sci. 2020, 258, 118238. [Google Scholar] [CrossRef]
- Newcomb, E.W.; Lukyanov, Y.; Smirnova, I.; Schnee, T.; Zagzag, D. Noscapine Induces Apoptosis in Human Glioma Cells by an Apoptosis-Inducing Factor-Dependent Pathway. Anti-Cancer Drugs 2008, 19, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Khanmoradi, M.; Ali Mard, S.; Aboutaleb, N.; Nobakht, M.; Mahmoudian, M. The Protective Activity of Noscapine on Renal Ischemia-Reperfusion Injury in Male Wistar Rat. Iran. J. Basic Med. Sci. 2014, 17, 244–249. [Google Scholar] [PubMed]
- Nour, M.; Scalzo, F.; Liebeskind, D.S. Ischemia-Reperfusion Injury in Stroke. Interv. Neurol. 2013, 1, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Dobrivojević, M.; Špiranec, K.; Sinđić, A. Involvement of Bradykinin in Brain Edema Development after Ischemic Stroke. Pflug. Arch. 2015, 467, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Yao, P.; Ma, S.; Zhang, C. Bradykinin Activates the Bradykinin B2 Receptor to Ameliorate Neuronal Injury in a Rat Model of Spinal Cord Ischemia-Reperfusion Injury. ACS Chem. Neurosci. 2021, 12, 1031–1038. [Google Scholar] [CrossRef]
- Mahmoudian, M.; Rezvani, M.; Rohani, M.; Benaissa, F.; Jalili, M.; Ghourchian, S. A Novel Effect of Noscapine on Patients with Massive Ischemic Stroke: A Pseudo-Randomized Clinical Trial. Iran. J. Neurol. 2015, 14, 12–16. [Google Scholar]
- Altinoz, M.A.; Topcu, G.; Hacimuftuoglu, A.; Ozpinar, A.; Ozpinar, A.; Hacker, E.; Elmaci, İ. Noscapine, a Non-Addictive Opioid and Microtubule-Inhibitor in Potential Treatment of Glioblastoma. Neurochem. Res. 2019, 44, 1796–1806. [Google Scholar] [CrossRef]
- Sung, B.; Ahn, K.S.; Aggarwal, B.B. Noscapine, a Benzylisoquinoline Alkaloid, Sensitizes Leukemic Cells to Chemotherapeutic Agents and Cytokines by Modulating the NF-ΚB Signaling Pathway. Cancer Res. 2010, 70, 3259–3268. [Google Scholar] [CrossRef]
- Chougule, M.B.; Patel, A.R.; Jackson, T.; Singh, M. Antitumor Activity of Noscapine in Combination with Doxorubicin in Triple Negative Breast Cancer. PLoS ONE 2011, 6, e17733. [Google Scholar] [CrossRef]
- Canisso, I.F.; Segabinazzi, L.G.T.M.; Fedorka, C.E. Persistent Breeding-Induced Endometritis in Mares—A Multifaceted Challenge: From Clinical Aspects to Immunopathogenesis and Pathobiology. Int. J. Mol. Sci. 2020, 21, 1432. [Google Scholar] [CrossRef]
- Woodward, E.M.; Christoffersen, M.; Campos, J.; Betancourt, A.; Horohov, D.; Scoggin, K.E.; Squires, E.L.; Troedsson, M.H.T. Endometrial Inflammatory Markers of the Early Immune Response in Mares Susceptible or Resistant to Persistent Breeding-Induced Endometritis. Reproduction 2013, 145, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Domino, M.; Jasinski, T.; Kautz, E.; Juszczuk-Kubiak, E.; Ferreira-Dias, G.; Zabielski, R.; Sady, M.; Gajewski, Z. Expression of Genes Involved in the NF-ΚB-Dependent Pathway of the Fibrosis in the Mare Endometrium. Theriogenology 2020, 147, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Rebordão, M.R.; Amaral, A.; Lukasik, K.; Szóstek-Mioduchowska, A.; Pinto-Bravo, P.; Galvão, A.; Skarzynski, D.J.; Ferreira-Dias, G. Impairment of the Antifibrotic Prostaglandin E2 Pathway May Influence Neutrophil Extracellular Traps–Induced Fibrosis in the Mare Endometrium. Domest. Anim. Endocrinol. 2019, 67, 1–10. [Google Scholar] [CrossRef]
- Cabezas, J.; Rojas, D.; Wong, Y.; Telleria, F.; Manriquez, J.; Mançanares, A.C.F.; Rodriguez-Alvarez, L.; Castro, F.O. In Vitro Preconditioning of Equine Adipose Mesenchymal Stem Cells with Prostaglandin E2, Substance P and Their Combination Changes the Cellular Protein Secretomics and Improves Their Immunomodulatory Competence without Compromising Stemness. Vet. Immunol. Immunopathol. 2020, 228, 110100. [Google Scholar] [CrossRef] [PubMed]
- Ye, K.; Ke, Y.; Keshava, N.; Shanks, J.; Kapp, J.A.; Tekmal, R.R.; Petros, J.; Joshi, H.C. Opium Alkaloid Noscapine Is an Antitumor Agent That Arrests Metaphase and Induces Apoptosis in Dividing Cells. Proc. Natl. Acad. Sci. USA 1998, 95, 1601–1606. [Google Scholar] [CrossRef] [PubMed]
- Kenney, R.M.; Doig, P.A. Equine endometrial biopsy. In Current Therapy in Theriogenology 2: Diagnosis, Treatment, and Prevention of Reproductive Diseases in Small and Large Animals, 2nd ed.; Morrow, D.A., Ed.; W.B. Saunders: Philadelphia, PA, USA, 1986; pp. 723–729. [Google Scholar]
- Schafer, W.R.; Fischer, L.; Roth, K.; Jullig, A.K.; Stuckenschneider, J.E.; Schwartz, P.; Weimer, M.; Orlowska-Volk, M.; Hanjalic-Beck, A.; Kranz, I.; et al. Critical Evaluation of Human Endometrial Explants as an Ex Vivo Model System: A Molecular Approach. Mol. Hum. Reprod. 2011, 17, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Fernald, R.D. Comprehensive Algorithm for Quantitative Real-Time Polymerase Chain Reaction. J. Comput. Biol. 2005, 12, 1047–1064. [Google Scholar] [CrossRef]
- Posch, A.; Kohn, J.; Oh, K.; Hammond, M.; Liu, N. V3 Stain-Free Workflow for a Practical, Convenient, and Reliable Total Protein Loading Control in Western Blotting. J. Vis. Exp. 2013, 82, 50948. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time of Incubation | LDH Activity (%) |
|---|---|
| 1 h | 95.19 ± 0.40 a |
| 24 h | 90.24 ± 0.70 b |
| 48 h | 87.93 ± 0.86 b |
| COL1A2 Gene | COL1 Protein | |||
|---|---|---|---|---|
| Factor/Factors Interaction | Mean (COL1A2/RPL32 mRNA Transcription) | p Value | Mean (COL1 Protein Abundance—AU) | p Value |
| MPO | 0.000425 | 0.744 | 0.007296 | 0.754 |
| NOSC | 0.007189 | 0.027 * | 0.000790 | 0.862 |
| Time of treatment | 0.001246 | 0.353 | 0.089686 | 0.066 |
| Estrous cycle phase | 0.196106 | <0.00001 *** | 0.088325 | 0.068 |
| MPO × NOSC | 0.004735 | 0.041 * | 0.155415 | 0.0035 ** |
| MPO × time of treatment | 0.001531 | 0.347 | 0.320149 | <0.0001 *** |
| MPO × estrous cycle phase | 0.002762 | 0.015 | 0.006394 | 0.781 |
| NOSC × time of treatment | 0.000826 | 0.450 | 0.000009 | 0.985 |
| NOSC × estrous cycle phase | 0.007010 | 0.029 * | 0.018122 | 0.404 |
| Time of treatment × estrous cycle phase | 0.000040 | 0.868 | 0.137148 | 0.023 * |
| MPO × NOSC × time of treatment | 0.009824 | 0.00017 ** | 0.046136 | 0.173 |
| MPO × NOSC × estrous cycle phase | 0.003486 | 0.093 | 0.187116 | 0.0012 ** |
| MPO × time of treatment × estrous cycle phase | 0.001567 | 0.339 | 0.053447 | 0.131 |
| NOSC × time of treatment × estrous cycle phase | 0.001222 | 0.358 | 0.093944 | 0.060 |
| Gene (Accession Number) | Sequence 5′-3′ | Amplicon |
|---|---|---|
| COL1A2 (XM_001492939.3) | Forward: CAAGGGCATTAGGGGACACA | 196 |
| Reverse: ACCCACACTTCCATCGCTTC | ||
| RPL32 (XM_001492042.6) | Forward: AGCCATCTACTCGGCGTCA | 144 |
| Reverse: GTCAATGCCTCTGGGTTTCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amaral, A.; Cebola, N.; Szóstek-Mioduchowska, A.; Rebordão, M.R.; Kordowitzki, P.; Skarzynski, D.; Ferreira-Dias, G. Inhibition of Myeloperoxidase Pro-Fibrotic Effect by Noscapine in Equine Endometrium. Int. J. Mol. Sci. 2023, 24, 3593. https://doi.org/10.3390/ijms24043593
Amaral A, Cebola N, Szóstek-Mioduchowska A, Rebordão MR, Kordowitzki P, Skarzynski D, Ferreira-Dias G. Inhibition of Myeloperoxidase Pro-Fibrotic Effect by Noscapine in Equine Endometrium. International Journal of Molecular Sciences. 2023; 24(4):3593. https://doi.org/10.3390/ijms24043593
Chicago/Turabian StyleAmaral, Ana, Nélio Cebola, Anna Szóstek-Mioduchowska, Maria Rosa Rebordão, Paweł Kordowitzki, Dariusz Skarzynski, and Graça Ferreira-Dias. 2023. "Inhibition of Myeloperoxidase Pro-Fibrotic Effect by Noscapine in Equine Endometrium" International Journal of Molecular Sciences 24, no. 4: 3593. https://doi.org/10.3390/ijms24043593