

Transcriptome-Based Traits of Radioresistant Sublines of Non-Small Cell Lung Cancer Cells
 , and
, and
Abstract
:1. Introduction
2. Results
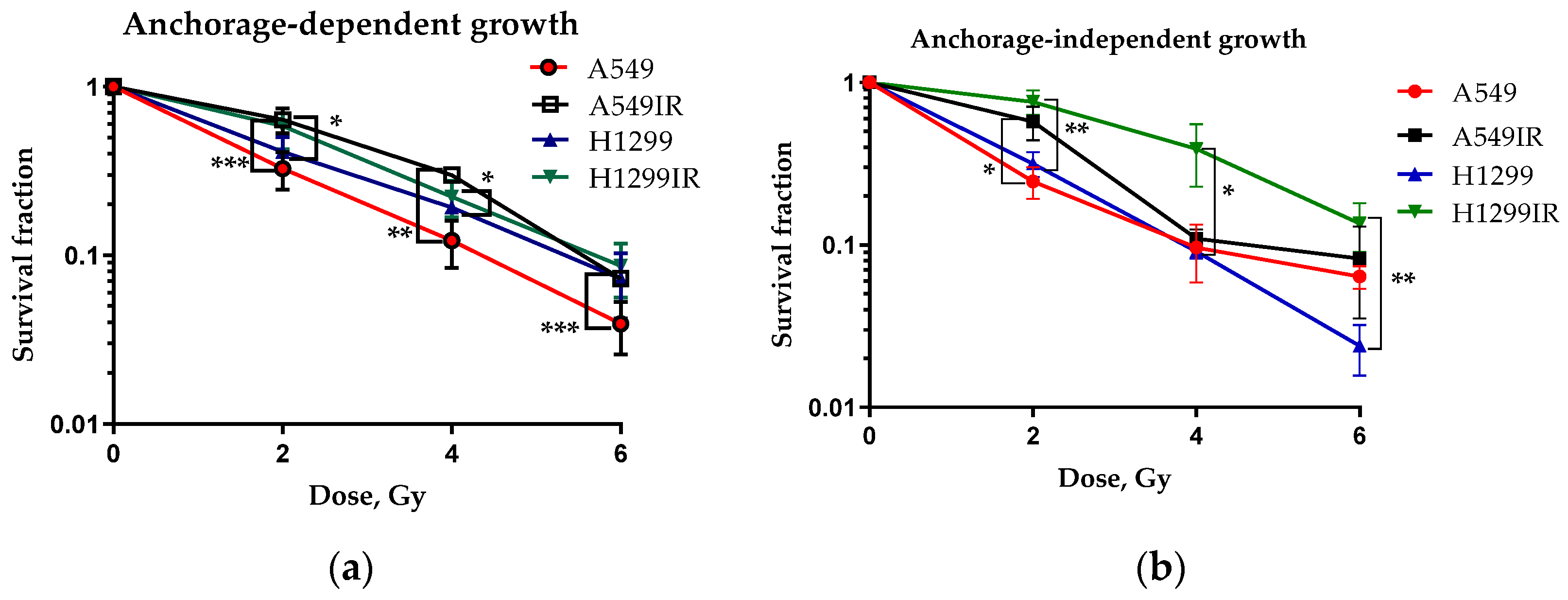
2.1. Establishment of the Radioresistant NSCLC Cells
2.2. Differential Gene Expression of NSCLC Cells
2.3. Radiation-Induced Transcriptome Alteration in Radioresistant NSCLC Cells through Pathway Activation Level (PAL) Analysis
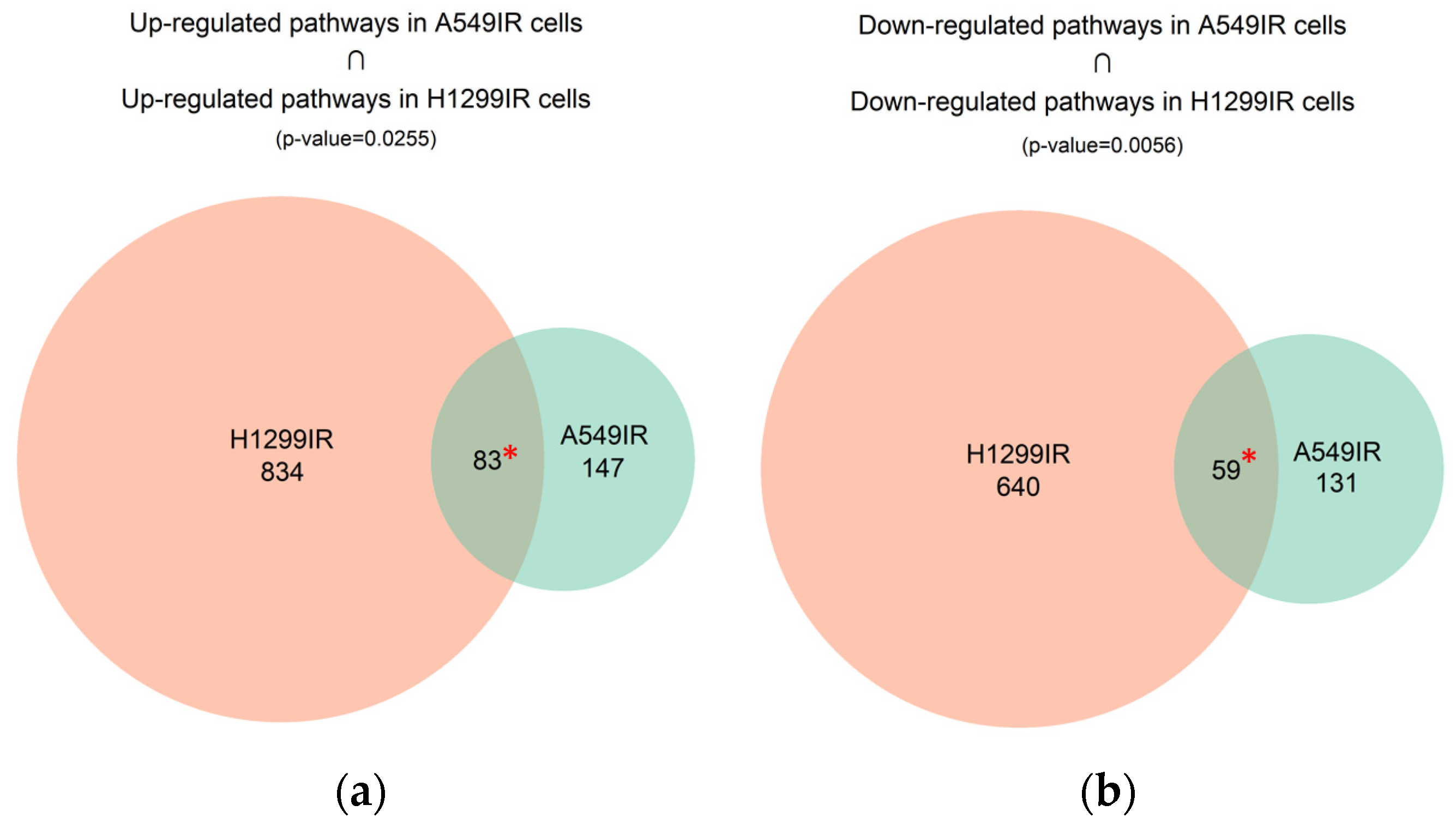
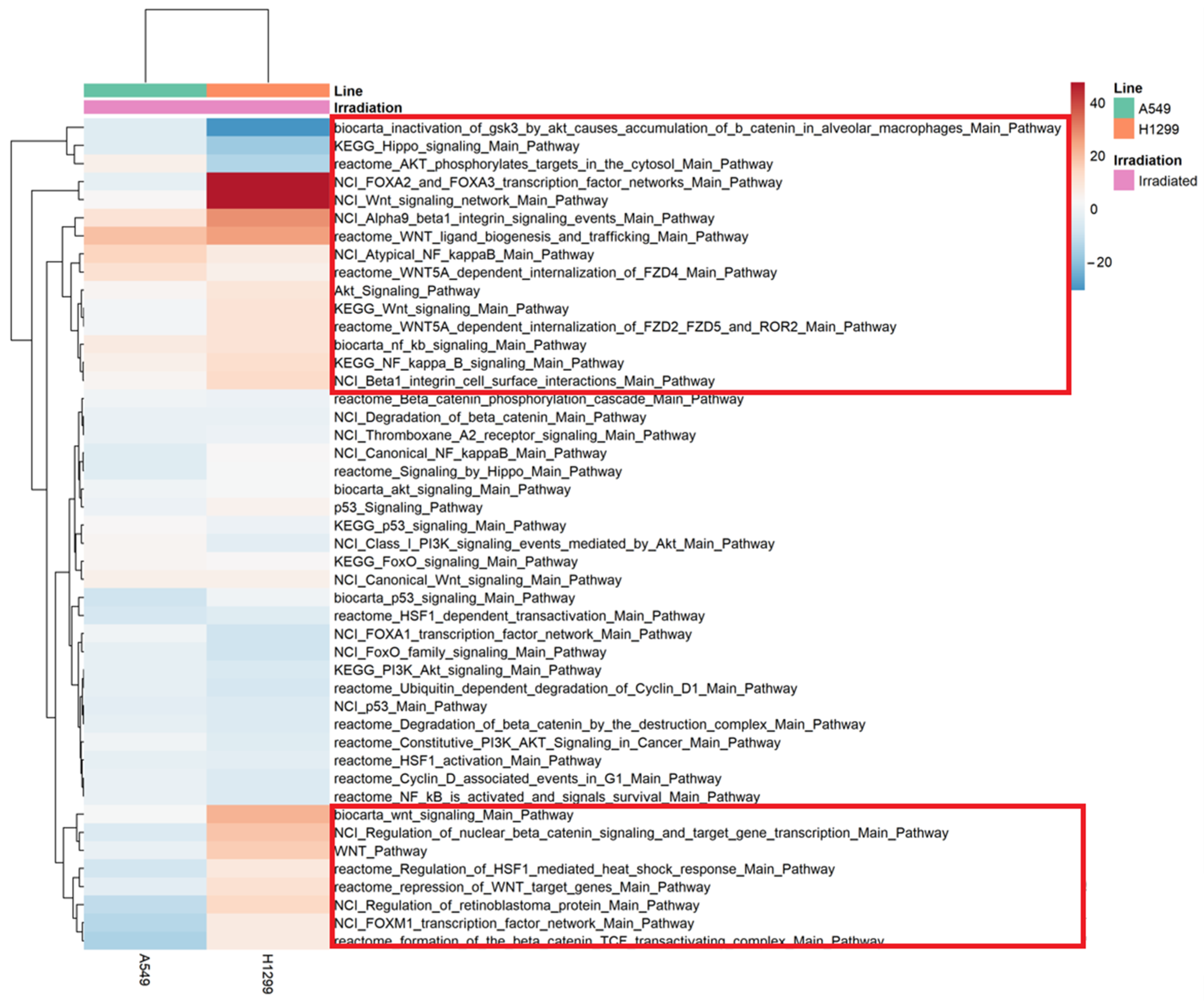
2.4. Differential Pathway Changes in NSCLC Cells
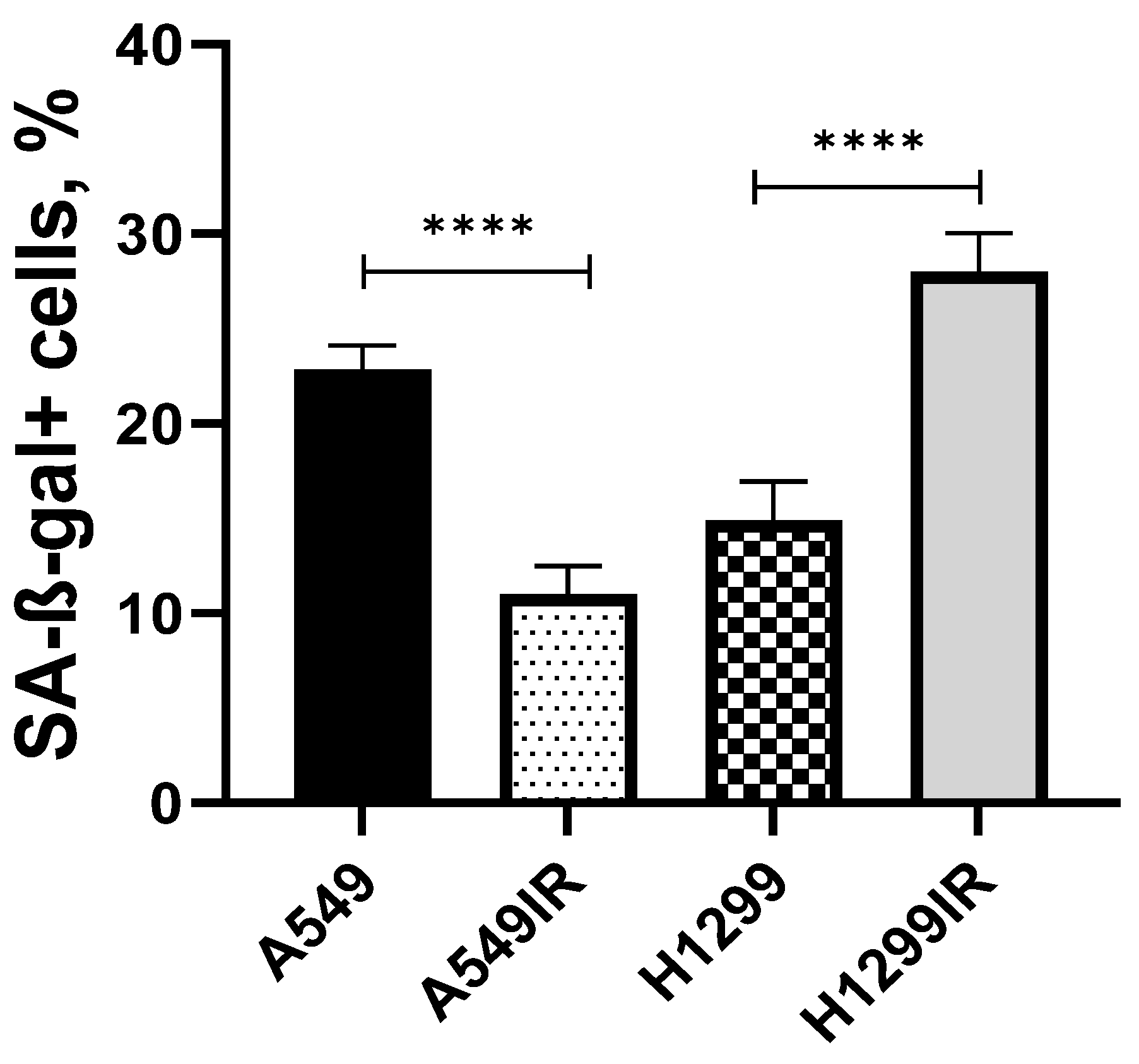
2.5. Charachteristics of Senescence-Associated Radioresistance in NSCLC Cells
3. Discussion
4. Materials and Methods
4.1. Cell Cultures and Irradiation
4.2. Clonogenic Test and Soft Agar Colony Formation
4.3. Transcriptomic Analysis
4.4. Bioinformatics Analysis
4.5. Gene Ontology (GO) Analysis
4.6. Analysis of Senescence-Associated β-Galactosidase (SA-β-Gal) Positive Cells
4.7. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thandra, K.C.; Barsouk, A.; Saginala, K.; Aluru, J.S.; Barsouk, A. Epidemiology of lung cancer. Contemp. Oncol. 2021, 25, 45–52. [Google Scholar] [CrossRef]
- Herbst, R.S.; Morgensztern, D.; Boshoff, C. The biology and management of non-small cell lung cancer. Nature 2018, 553, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Prabavathy, D.; Swarnalatha, Y.; Ramadoss, N. Lung cancer stem cells—Origin, characteristics and therapy. Stem Cell Investig. 2018, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Césaire, M.; Montanari, J.; Curcio, H.; Lerouge, D.; Gervais, R.; Demontrond, P.; Balosso, J.; Chevalier, F. Radioresistance of Non-Small Cell Lung Cancers and Therapeutic Perspectives. Cancers 2022, 14, 2829. [Google Scholar] [CrossRef]
- Borisov, N.M.; Terekhanova, N.V.; Aliper, A.M.; Venkova, L.S.; Smirnov, P.Y.; Roumiantsev, S.; Korzinkin, M.B.; Zhavoronkov, A.A.; Buzdin, A.A. Signaling pathways activation profiles make better markers of cancer than expression of individual genes. Oncotarget 2014, 5, 10198–10205. [Google Scholar] [CrossRef]
- Kamashev, D.; Sorokin, M.; Kochergina, I.; Drobyshev, A.; Vladimirova, U.; Zolotovskaia, M.; Vorotnikov, I.; Shaban, N.; Raevskiy, M.; Kuzmin, D.; et al. Human blood serum can donor-specifically antagonize effects of EGFR-targeted drugs on squamous carcinoma cell growth. Heliyon 2021, 7, e06394. [Google Scholar] [CrossRef]
- Crispo, F.; Notarangelo, T.; Pietrafesa, M.; Lettini, G.; Storto, G.; Sgambato, A.; Maddalena, F.; Landriscina, M. BRAF Inhibitors in Thyroid Cancer: Clinical Impact, Mechanisms of Resistance and Future Perspectives. Cancers 2019, 11, 1388. [Google Scholar] [CrossRef]
- Pustovalova, M.; Alhaddad, L.; Smetanina, N.; Chigasova, A.; Blokhina, T.; Chuprov-Netochin, R.; Osipov, A.N.; Leonov, S. The p53–53BP1-Related Survival of A549 and H1299 Human Lung Cancer Cells after Multifractionated Radiotherapy Demonstrated Different Response to Additional Acute X-ray Exposure. Int. J. Mol. Sci. 2020, 21, 3342. [Google Scholar] [CrossRef]
- Pustovalova, M.; Alhaddad, L.; Blokhina, T.; Smetanina, N.; Chigasova, A.; Chuprov-Netochin, R.; Eremin, P.; Gilmutdinova, I.; Osipov, A.; Leonov, S. The CD44high Subpopulation of Multifraction Irradiation-Surviving NSCLC Cells Exhibits Partial EMT-Program Activation and DNA Damage Response Depending on Their p53 Status. Int. J. Mol. Sci. 2021, 22, 2369. [Google Scholar] [CrossRef]
- Pustovalova, M.V.; Guryanova, A.A.; Sorokin, M.I.; Suntsova, M.V.; Buzdin, A.A.; Alhaddad, L.; Osipov, A.N.; Leonov, S.V. Transcriptomic Analysis of DNA Repair Pathways in Human Non-Small Cell Lung Cancer Cells Surviving Multifraction X-ray Irradiation. Bull. Exp. Biol. Med. 2022, 173, 454–458. [Google Scholar] [CrossRef]
- Zheng, Q.; Wang, X.-J. GOEAST: A web-based software toolkit for Gene Ontology enrichment analysis. Nucleic Acids Res. 2008, 36, W358–W363. [Google Scholar] [CrossRef]
- Borisov, N.; Sorokin, M.; Garazha, A.; Buzdin, A. Quantitation of Molecular Pathway Activation Using RNA Sequencing Data. In Nucleic Acid Detection and Structural Investigations; Humana: New York, NY, USA, 2019; Volume 2063, pp. 189–206. [Google Scholar] [CrossRef]
- Croft, D.; Mundo, A.F.; Haw, R.; Milacic, M.; Weiser, J.; Wu, G.; Caudy, M.; Garapati, P.; Gillespie, M.; Kamdar, M.R.; et al. The Reactome pathway knowledgebase. Nucleic Acids Res. 2013, 42, D472–D477. [Google Scholar] [CrossRef]
- Schaefer, C.F.; Anthony, K.; Krupa, S.; Buchoff, J.; Day, M.; Hannay, T.; Buetow, K.H. PID: The Pathway Interaction Database. Nucleic Acids Res. 2008, 37, D674–D679. [Google Scholar] [CrossRef]
- Zhu, W.; Wang, H.; Zhu, D. Wnt/β-catenin signaling pathway in lung cancer. Med. Drug Discov. 2021, 13, 100113. [Google Scholar] [CrossRef]
- Schoetz, U.; Klein, D.; Hess, J.; Shnayien, S.; Spoerl, S.; Orth, M.; Mutlu, S.; Hennel, R.; Sieber, A.; Ganswindt, U.; et al. Early senescence and production of senescence-associated cytokines are major determinants of radioresistance in head-and-neck squamous cell carcinoma. Cell Death Dis. 2021, 12, 1162. [Google Scholar] [CrossRef]
- Chien, Y.; Scuoppo, C.; Wang, X.; Fang, X.; Balgley, B.; Bolden, J.E.; Premsrirut, P.; Luo, W.; Chicas, A.; Lee, C.S.; et al. Control of the senescence-associated secretory phenotype by NF-κB promotes senescence and enhances chemosensitivity. Genes Dev. 2011, 25, 2125–2136. [Google Scholar] [CrossRef]
- Pustovalova, M.; Blokhina, T.; Alhaddad, L.; Chigasova, A.; Chuprov-Netochin, R.; Veviorskiy, A.; Filkov, G.; Osipov, A.N.; Leonov, S. CD44+ and CD133+ Non-Small Cell Lung Cancer Cells Exhibit DNA Damage Response Pathways and Dormant Polyploid Giant Cancer Cell Enrichment Relating to Their p53 Status. Int. J. Mol. Sci. 2022, 23, 4922. [Google Scholar] [CrossRef]
- Alhaddad, L.; Pustovalova, M.; Blokhina, T.; Chuprov-Netochin, R.; Osipov, A.; Leonov, S. IR-Surviving NSCLC Cells Exhibit Different Patterns of Molecular and Cellular Reactions Relating to the Multifraction Irradiation Regimen and p53-Family Proteins Expression. Cancers 2021, 13, 2669. [Google Scholar] [CrossRef]
- Zhao, X.; Jin, X.; Zhang, Q.; Liu, R.; Luo, H.; Yang, Z.; Geng, Y.; Feng, S.; Li, C.; Wang, L.; et al. Silencing of the lncRNA H19 enhances sensitivity to X-ray and carbon-ions through the miR-130a-3p /WNK3 signaling axis in NSCLC cells. Cancer Cell Int. 2021, 21, 644. [Google Scholar] [CrossRef]
- Liu, B.; Li, H.; Liu, X.; Li, F.; Chen, W.; Kuang, Y.; Zhao, X.; Li, L.; Yu, B.; Jin, X.; et al. CircZNF208 enhances the sensitivity to X-rays instead of carbon-ions through the miR-7-5p /SNCA signal axis in non-small-cell lung cancer cells. Cell. Signal. 2021, 84, 110012. [Google Scholar] [CrossRef]
- Guo, W.; Xie, L.; Zhao, L.; Zhao, Y. mRNA and microRNA expression profiles of radioresistant NCI-H520 non-small cell lung cancer cells. Mol. Med. Rep. 2015, 12, 1857–1867. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Cao, Q.; Ding, X.; Gao, D.; Yang, Q.; Li, B. Radiotranscriptomics signature-based predictive nomograms for radiotherapy response in patients with nonsmall cell lung cancer: Combination and association of CT features and serum miRNAs levels. Cancer Med. 2020, 9, 5065–5074. [Google Scholar] [CrossRef] [PubMed]
- Mboge, M.Y.; Mahon, B.P.; McKenna, R.; Frost, S.C. Carbonic Anhydrases: Role in pH Control and Cancer. Metabolites 2018, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Annan, D.A.; Maishi, N.; Soga, T.; Dawood, R.; Li, C.; Kikuchi, H.; Hojo, T.; Morimoto, M.; Kitamura, T.; Alam, M.T.; et al. Carbonic anhydrase 2 (CAII) supports tumor blood endothelial cell survival under lactic acidosis in the tumor microenvironment. Cell Commun. Signal. 2019, 17, 169. [Google Scholar] [CrossRef]
- Hermawan, A.; Putri, H. Integrative bioinformatics analysis reveals miR-494 and its target genes as predictive biomarkers of trastuzumab-resistant breast cancer. J. Egypt. Natl. Cancer Inst. 2020, 32, 16. [Google Scholar] [CrossRef]
- Kim, M.; Kim, D.J. GFRA1: A Novel Molecular Target for the Prevention of Osteosarcoma Chemoresistance. Int. J. Mol. Sci. 2018, 19, 1078. [Google Scholar] [CrossRef]
- Wang, L.; Shen, X.; Wang, Z.; Xiao, X.; Wei, P.; Wang, Q.; Ren, F.; Wang, Y.; Liu, Z.; Sheng, W.; et al. A molecular signature for the prediction of recurrence in colorectal cancer. Mol. Cancer 2015, 14, 22. [Google Scholar] [CrossRef]
- Liu, J.; Yu, N.; Feng, X.; He, Y.; Lv, K.; Zhu, H.; Wang, J. Loss of EphA7 Expression in Basal Cell Carcinoma by Hypermethylation of CpG Islands in the Promoter Region. Anal. Cell. Pathol. 2022, 2022, 4220786. [Google Scholar] [CrossRef]
- Wang, T.-H.; Hsia, S.-M.; Shieh, T.-M. Lysyl Oxidase and the Tumor Microenvironment. Int. J. Mol. Sci. 2016, 18, 62. [Google Scholar] [CrossRef] [Green Version]
- Abdulkareem, N.M.; Bhat, R.; Qin, L.; Vasaikar, S.; Gopinathan, A.; Mitchell, T.; Shea, M.J.; Nanda, S.; Thangavel, H.; Zhang, B.; et al. A novel role of ADGRF1 (GPR110) in promoting cellular quiescence and chemoresistance in human epidermal growth factor receptor 2-positive breast cancer. FASEB J. 2021, 35, e21719. [Google Scholar] [CrossRef]
- Zhong, L.; Gan, X.; Deng, X.; Shen, F.; Feng, J.; Cai, W.; Liu, Q.; Miao, J.; Zheng, B.; Xu, B. Potential five-mRNA signature model for the prediction of prognosis in patients with papillary thyroid carcinoma. Oncol. Lett. 2020, 20, 2302–2310. [Google Scholar] [CrossRef]
- Upadhyay, G. Emerging Role of Lymphocyte Antigen-6 Family of Genes in Cancer and Immune Cells. Front. Immunol. 2019, 10, 819. [Google Scholar] [CrossRef]
- Alves, C.L.; Elias, D.; Lyng, M.B.; Bak, M.; Ditzel, H.J. SNAI2 upregulation is associated with an aggressive phenotype in fulvestrant-resistant breast cancer cells and is an indicator of poor response to endocrine therapy in estrogen receptor-positive metastatic breast cancer. Breast Cancer Res. 2018, 20, 60. [Google Scholar] [CrossRef]
- Luan, F.; Li, X.; Cheng, X.; Huangfu, L.; Han, J.; Guo, T.; Du, H.; Wen, X.; Ji, J. TNFRSF11B activates Wnt/β-catenin signaling and promotes gastric cancer progression. Int. J. Biol. Sci. 2020, 16, 1956–1971. [Google Scholar] [CrossRef]
- Xie, W.; Qiao, X.; Shang, L.; Dou, J.; Yang, X.; Qiao, S.; Wu, Y. Knockdown of ZNF233 suppresses hepatocellular carcinoma cell proliferation and tumorigenesis. Gene 2018, 679, 179–185. [Google Scholar] [CrossRef]
- Tepedelen, B.E.; Kirmizibayrak, P.B. Endoplasmic Reticulum-Associated Degradation (ERAD). In Endoplasmic Reticulum; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, J.; Liu, L.; Li, J. The Crucial Role of Demannosylating Asparagine-Linked Glycans in ERADicating Misfolded Glycoproteins in the Endoplasmic Reticulum. Front. Plant Sci. 2021, 11, 625033. [Google Scholar] [CrossRef]
- Habiba, U.; Rafiq, M.; Khawar, M.B.; Nazir, B.; Haider, G.; Nazir, N. The multifaceted role of IL-12 in cancer. Adv. Cancer Biol.-Metastasis 2022, 5, 100053. [Google Scholar] [CrossRef]
- Wu, M.-F.; Wang, Y.-C.; Li, H.-T.; Chen, W.-C.; Liao, C.-H.; Shih, T.-C.; Chang, W.-S.; Tsai, C.-W.; Hsia, T.-C.; Bau, D.-T. The Contribution of Interleukin-12 Genetic Variations to Taiwanese Lung Cancer. Anticancer Res. 2018, 38, 6321–6327. [Google Scholar] [CrossRef]
- Zhang, W.; Dang, S.; Zhang, G.; He, H.; Wen, X. Genetic polymorphisms of IL-10, IL-18 and IL12B are associated with risk of non-small cell lung cancer in a Chinese Han population. Int. Immunopharmacol. 2019, 77, 105938. [Google Scholar] [CrossRef]
- Núñez-Marrero, A.; Arroyo, N.; Godoy-Munoz, L.; Rahman, M.Z.; Matta, J.L.; Dutil, J. SNPs in the interleukin-12 signaling pathway are associated with breast cancer risk in Puerto Rican women. Oncotarget 2020, 11, 3420–3431. [Google Scholar] [CrossRef]
- Herranz, N.; Gil, J. Mechanisms and functions of cellular senescence. J. Clin. Investig. 2018, 128, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lankhorst, L.; Bernards, R. Exploiting senescence for the treatment of cancer. Nat. Rev. Cancer 2022, 22, 340–355. [Google Scholar] [CrossRef] [PubMed]
- Ruhland, M.K.; Loza, A.J.; Capietto, A.-H.; Luo, X.; Knolhoff, B.L.; Flanagan, K.C.; Belt, B.A.; Alspach, E.; Leahy, K.; Luo, J.; et al. Stromal senescence establishes an immunosuppressive microenvironment that drives tumorigenesis. Nat. Commun. 2016, 7, 11762. [Google Scholar] [CrossRef] [PubMed]
- Marin, I.; Boix, O.; Garcia-Garijo, A.; Sirois, I.; Caballe, A.; Zarzuela, E.; Ruano, I.; Stephan-Otto Attolini, C.; Prats, N.; Lopez-Dominguez, J.A.; et al. Cellular senescence is immunogenic and promotes anti-tumor immunity. Cancer Discov 2022, 12, 2154115. [Google Scholar] [CrossRef]
- Hu, X.; Guo, L.; Liu, G.; Dai, Z.; Wang, L.; Zhang, J.; Wang, J. Novel cellular senescence-related risk model identified as the prognostic biomarkers for lung squamous cell carcinoma. Front. Oncol. 2022, 12, 997702. [Google Scholar] [CrossRef]
- Xiong, J.; Jiang, P.; Zhong, L.; Wang, Y. The Novel Tumor Suppressor Gene ZNF24 Induces THCA Cells Senescence by Regulating Wnt Signaling Pathway, Resulting in Inhibition of THCA Tumorigenesis and Invasion. Front. Oncol. 2021, 11, 646511. [Google Scholar] [CrossRef]
- Pang, B.; Wang, Y.; Chang, X. A Novel Tumor Suppressor Gene, ZNF24, Inhibits the Development of NSCLC by Inhibiting the WNT Signaling Pathway to Induce Cell Senescence. Front. Oncol. 2021, 11, 664369. [Google Scholar] [CrossRef]
- Cuollo, L.; Antonangeli, F.; Santoni, A.; Soriani, A. The Senescence-Associated Secretory Phenotype (SASP) in the Challenging Future of Cancer Therapy and Age-Related Diseases. Biology 2020, 9, 485. [Google Scholar] [CrossRef]
- Cong, L.; Qiu, Z.-Y.; Zhao, Y.; Wang, W.-B.; Wang, C.-X.; Shen, H.-C.; Han, J.-Q. Loss of β-arrestin-2 and Activation of CXCR2 Correlate with Lymph Node Metastasis in Non-small Cell Lung Cancer. J. Cancer 2017, 8, 2785–2792. [Google Scholar] [CrossRef]
- Giampieri, S.; Manning, C.; Hooper, S.; Jones, L.; Hill, C.; Sahai, E. Localized and reversible TGFβ signalling switches breast cancer cells from cohesive to single cell motility. Nature 2009, 11, 1287–1296. [Google Scholar] [CrossRef]
- Jackstadt, R.; van Hooff, S.R.; Leach, J.D.; Cortes-Lavaud, X.; Lohuis, J.O.; Ridgway, R.A.; Wouters, V.M.; Roper, J.; Kendall, T.J.; Roxburgh, C.S.; et al. Epithelial NOTCH Signaling Rewires the Tumor Microenvironment of Colorectal Cancer to Drive Poor-Prognosis Subtypes and Metastasis. Cancer Cell 2019, 36, 319–336. [Google Scholar] [CrossRef]
- Del Valle, L.; Dai, L.; Lin, H.; Lin, Z.; Chen, J.; Post, S.R.; Qin, Z. Role of EIF4G1 network in non-small cell lung cancers (NSCLC) cell survival and disease progression. J. Cell. Mol. Med. 2021, 25, 2795–2805. [Google Scholar] [CrossRef]
- Chua, Y.L.; Ito, Y.; Pole, J.C.M.; Newman, S.; Chin, S.F.; Stein, R.C.; Ellis, I.O.; Caldas, C.; O’Hare, M.J.; Murrell, A.; et al. The NRG1 gene is frequently silenced by methylation in breast cancers and is a strong candidate for the 8p tumour suppressor gene. Oncogene 2009, 28, 4041–4052. [Google Scholar] [CrossRef]
- Piomelli, D. Endocannabinoids. In Encyclopedia of Biological Chemistry; Academic Press: Cambridge, MA, USA, 2013; pp. 194–196. [Google Scholar] [CrossRef]
- Maccarrone, M. Metabolism of the Endocannabinoid Anandamide: Open Questions after 25 Years. Front. Mol. Neurosci. 2017, 10, 166. [Google Scholar] [CrossRef]
- Kozak, K.R.; Crews, B.C.; Morrow, J.D.; Wang, L.-H.; Ma, Y.H.; Weinander, R.; Jakobsson, P.-J.; Marnett, L.J. Metabolism of the Endocannabinoids, 2-Arachidonylglycerol and Anandamide, into Prostaglandin, Thromboxane, and Prostacyclin Glycerol Esters and Ethanolamides. J. Biol. Chem. 2002, 277, 44877–44885. [Google Scholar] [CrossRef]
- Qiao, L.; Kozoni, V.; Tsioulias, G.J.; Koutsos, M.I.; Hanif, R.; Shiff, S.J.; Rigas, B. Selected eicosanoids increase the proliferation rate of human colon carcinoma cell lines and mouse colonocytes in vivo. Biochim. Biophys. Acta BBA Lipids Lipid Metab. 1995, 1258, 215–223. [Google Scholar] [CrossRef]
- Tsujii, M.; Kawano, S.; Tsuji, S.; Sawaoka, H.; Hori, M.; DuBois, R.N. Cyclooxygenase Regulates Angiogenesis Induced by Colon Cancer Cells. Cell 1998, 93, 705–716. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.J.; Kim, N.; Seong, K.M.; Youn, H.; Youn, B. Investigation of Radiation-induced Transcriptome Profile of Radioresistant Non-small Cell Lung Cancer A549 Cells Using RNA-seq. PLoS ONE 2013, 8, e59319. [Google Scholar] [CrossRef]
- Li, W.; Yue, W.; Zhang, L.; Zhao, X.; Ma, L.; Yang, X.; Zhang, C.; Wang, Y.; Gu, M. COX-2 silencing inhibits cell proliferation in A549 cell. Chin.-Ger. J. Clin. Oncol. 2011, 10, 423–427. [Google Scholar] [CrossRef]
- Ding, C.; Liu, J.; Zhang, J.; Wan, Y.; Hu, L.; Charwudzi, A.; Zhan, H.; Meng, Y.; Zheng, H.; Wang, H.; et al. Tumor Endothelial Marker 8 Promotes Proliferation and Metastasis via the Wnt/β-Catenin Signaling Pathway in Lung Adenocarcinoma. Front. Oncol. 2021, 11, 712371. [Google Scholar] [CrossRef]
- Chen, D.; Bhat-Nakshatri, P.; Goswami, C.; Badve, S.; Nakshatri, H. ANTXR1, a Stem Cell-Enriched Functional Biomarker, Connects Collagen Signaling to Cancer Stem-like Cells and Metastasis in Breast Cancer. Cancer Res 2013, 73, 5821–5833. [Google Scholar] [CrossRef] [PubMed]
- Alcalá, S.; Martinelli, P.; Hermann, P.C.; Heeschen, C.; Sainz, B. The Anthrax Toxin Receptor 1 (ANTXR1) Is Enriched in Pancreatic Cancer Stem Cells Derived from Primary Tumor Cultures. Stem Cells Int. 2019, 2019, 1378639. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int. J. Biol. Sci. 2018, 14, 1483–1496. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Li, J.; Qiang, H.; Lei, Y.; Chang, Q.; Zhong, R.; Stella, G.M.; Gelsomino, F.; Kim, Y.W.; Abed, A.; et al. A retrospective study for prognostic significance of type II diabetes mellitus and hemoglobin A1c levels in non-small cell lung cancer patients treated with pembrolizumab. Transl. Lung Cancer Res. 2021, 11, 1619–1630. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-H.; Chen, W.-M.; Chen, M.; Shia, B.-C.; Wu, S.-Y. Association of Diabetes Severity and Mortality with Lung Squamous Cell Carcinoma. Cancers 2022, 14, 2553. [Google Scholar] [CrossRef]
- Zhu, L.; Cao, H.; Zhang, T.; Shen, H.; Dong, W.; Wang, L.; Du, J. The Effect of Diabetes Mellitus on Lung Cancer Prognosis: A PRISMA-compliant meta-analysis of cohort studies. Medicine 2016, 95, e3528. [Google Scholar] [CrossRef]
- Luo, J.; Hendryx, M.; Qi, L.; Ho, G.Y.; Margolis, K. Pre-existing diabetes and lung cancer prognosis. Br. J. Cancer 2016, 115, 76–79. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Bai, Y. IGF-1 activates the P13K/AKT signaling pathway via upregulation of secretory clusterin. Mol. Med. Rep. 2012, 6, 1433–1437. [Google Scholar] [CrossRef]
- Vafopoulou, X.; Steel, C.G. Insulin-like and testis ecdysiotropin neuropeptides are regulated by the circadian timing system in the brain during larval–adult development in the insect Rhodnius prolixus (Hemiptera). Gen. Comp. Endocrinol. 2012, 179, 277–288. [Google Scholar] [CrossRef]
- Henriksen, E.J.; Dokken, B.B. Role of glycogen synthase kinase-3 in insulin resistance and type 2 diabetes. Curr. Drug Targets 2006, 7, 1435–1441. [Google Scholar] [CrossRef]
- Silva-Filho, J.L.; Caruso-Neves, C.; Pinheiro, A.A.S. IL-4: An important cytokine in determining the fate of T cells. Biophys. Rev. 2014, 6, 111–118. [Google Scholar] [CrossRef]
- Bankaitis, K.V.; Fingleton, B. Targeting IL4/IL4R for the treatment of epithelial cancer metastasis. Clin. Exp. Metastasis 2015, 32, 847–856. [Google Scholar] [CrossRef]
- Shi, X.; Dong, A.; Jia, X.; Zheng, G.; Wang, N.; Wang, Y.; Yang, C.; Lu, J.; Yang, Y. Integrated analysis of single-cell and bulk RNA-sequencing identifies a signature based on T-cell marker genes to predict prognosis and therapeutic response in lung squamous cell carcinoma. Front. Immunol. 2022, 13, 992990. [Google Scholar] [CrossRef]
- Rapp, J.; Jaromi, L.; Kvell, K.; Miskei, G.; Pongracz, J.E. WNT signaling—Lung cancer is no exception. Respir. Res. 2017, 18, 167. [Google Scholar] [CrossRef]
- Takahashi-Yanaga, F.; Kahn, M. Targeting Wnt Signaling: Can We Safely Eradicate Cancer Stem Cells? Clin. Cancer Res. 2010, 16, 3153–3162. [Google Scholar] [CrossRef]
- Stewart, D.J. Wnt Signaling Pathway in Non-Small Cell Lung Cancer. JNCI J. Natl. Cancer Inst. 2014, 106, djt356. [Google Scholar] [CrossRef]
- Nalbantoglu, B.; Durmu, S.; Ülgen, K.Ö. Wnt Signaling Network in Homo Sapiens. In Cell Metabolism—Cell Homeostasis and Stress Response; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.; Wang, X.; Wang, Y.; Ma, D. Wnt/β-catenin signaling regulates cancer stem cells in lung cancer A549 cells. Biochem. Biophys. Res. Commun. 2010, 392, 373–379. [Google Scholar] [CrossRef]
- Mazieres, J.; He, B.; You, L.; Xu, Z.; Jablons, D.M. Wnt signaling in lung cancer. Cancer Lett. 2005, 222, 1–10. [Google Scholar] [CrossRef]
- Jaiswal, P.K.; Goel, A.; Mittal, R.D. Survivin: A molecular biomarker in cancer. Indian J. Med. Res. 2015, 141, 389–397. [Google Scholar] [CrossRef]
- Tenbaum, S.P.; Ordóñez-Morán, P.; Puig, I.; Chicote, I.; Arqués, O.; Landolfi, S.; Fernández, Y.; Herance, J.R.; Gispert, J.D.; Mendizabal, L.; et al. β-catenin confers resistance to PI3K and AKT inhibitors and subverts FOXO3a to promote metastasis in colon cancer. Nat. Med. 2012, 18, 892–901. [Google Scholar] [CrossRef]
- Pangon, L.; Mladenova, D.; Watkins, L.; Van Kralingen, C.; Currey, N.; Al-Sohaily, S.; Lecine, P.; Borg, J.-P.; Kohonen-Corish, M.R. MCC inhibits β-catenin transcriptional activity by sequestering DBC1 in the cytoplasm. Int. J. Cancer 2014, 136, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Borowicz, S.; Van Scoyk, M.; Avasarala, S.; Karuppusamy Rathinam, M.K.; Tauler, J.; Bikkavilli, R.K.; Winn, R.A. The soft agar colony formation assay. JoVE J. Vis. Exp. 2014, 92, e51998. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Sorokin, M.; Kholodenko, R.; Suntsova, M.; Malakhova, G.; Garazha, A.; Kholodenko, I.; Poddubskaya, E.; Lantsov, D.; Stilidi, I.; Arhiri, P.; et al. Oncobox Bioinformatical Platform for Selecting Potentially Effective Combinations of Target Cancer Drugs Using High-Throughput Gene Expression Data. Cancers 2018, 10, 365. [Google Scholar] [CrossRef]
- Sorokin, M.; Borisov, N.; Kuzmin, D.; Gudkov, A.; Zolotovskaia, M.; Garazha, A.; Buzdin, A. Algorithmic Annotation of Functional Roles for Components of 3044 Human Molecular Pathways. Front. Genet. 2021, 12, 617059. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Description | p-Value Adjusted | Genes |
|---|---|---|---|
| GO:0002934 | desmosome organization | 0.019522 | SNAI2 |
| GO:0030656 | regulation of vitamin metabolic process | 0.019522 | SNAI2 |
| GO:0042368 | vitamin D biosynthetic process | 0.019522 | SNAI2 |
| GO:0048251 | elastic fiber assembly | 0.019522 | LOX |
| GO:0048755 | branching morphogenesis of a nerve | 0.019522 | EPHA7 |
| GO:0070561 | vitamin D receptor signaling pathway | 0.019522 | SNAI2 |
| GO:0072178 | nephric duct morphogenesis | 0.019522 | EPHA7 |
| GO:1905809 | negative regulation of synapse organization | 0.019522 | EPHA7 |
| GO:0061314 | Notch signaling involved in heart development | 0.019522 | SNAI2 |
| GO:0033629 | negative regulation of cell adhesion mediated by integrin | 0.019522 | SNAI2 |
| GO:0042362 | fat-soluble vitamin biosynthetic process | 0.019522 | SNAI2 |
| GO:0072176 | nephric duct development | 0.019522 | EPHA7 |
| GO:0018158 | protein oxidation | 0.019522 | LOX |
| GO:0035791 | platelet-derived growth factor receptor-beta signaling pathway | 0.019522 | LOX |
| GO:0010839 | negative regulation of keratinocyte proliferation | 0.019522 | SNAI2 |
| GO:0042481 | regulation of odontogenesis | 0.019522 | TNFRSF11B |
| GO:0043518 | negative regulation of DNA damage response, signal transduction by p53 class mediator | 0.019522 | SNAI2 |
| GO:0003198 | epithelial to mesenchymal transition involved in endocardial cushion formation | 0.019522 | SNAI2 |
| GO:0045779 | negative regulation of bone resorption | 0.019522 | TNFRSF11B |
| GO:0048670 | regulation of collateral sprouting | 0.019522 | EPHA7 |
| GO:2000811 | negative regulation of anoikis | 0.019522 | SNAI2 |
| GO:0032026 | response to magnesium ion | 0.019522 | TNFRSF11B |
| GO:0046851 | negative regulation of bone remodeling | 0.019522 | TNFRSF11B |
| GO:0060973 | cell migration involved in heart development | 0.019522 | SNAI2 |
| GO:0009110 | vitamin biosynthetic process | 0.019522 | SNAI2 |
| GO:0031290 | retinal ganglion cell axon guidance | 0.019522 | EPHA7 |
| GO:0032793 | positive regulation of CREB transcription factor activity | 0.019522 | ADGRF1 |
| GO:0034104 | negative regulation of tissue remodeling | 0.019522 | TNFRSF11B |
| GO:0046716 | muscle cell cellular homeostasis | 0.019522 | LOX |
| GO:0150146 | cell junction disassembly | 0.019522 | SNAI2 |
| Genes | A549IR LFC | A549IR p-Value | H1299IR LFC | H1299IR p-Value |
|---|---|---|---|---|
| CCL20 | −1.325 | 0.246 | −0.085 | 0.791 |
| CCL3 | 0.079 | 0.810 | −0.085 | 0.791 |
| CXCL1 | 0.043 | 0.895 | 0.739 | 0.134 |
| CXCL2 | −0.448 | 0.348 | −0.037 | 0.917 |
| CXCL3 | −0.247 | 0.348 | 0.054 | 0.924 |
| CXCL5 | 0.513 | 0.119 | −0.085 | 0.791 |
| CXCL8 | −0.299 | 0.424 | 3.544 | 0.000 |
| FGF2 | −0.066 | 0.760 | −0.194 | 0.560 |
| IL1A | 0.773 | 0.354 | −0.085 | 0.791 |
| IL1B | 0.299 | 0.582 | 0.283 | 0.738 |
| IL6 | 1.494 | 0.006 | 1.500 | 0.004 |
| NRG1 | −0.322 | 0.212 | −2.212 | 0.009 |
| TGFB2 | −0.037 | 0.863 | 1.579 | 0.003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pustovalova, M.; Malakhov, P.; Guryanova, A.; Sorokin, M.; Suntsova, M.; Buzdin, A.; Osipov, A.N.; Leonov, S. Transcriptome-Based Traits of Radioresistant Sublines of Non-Small Cell Lung Cancer Cells. Int. J. Mol. Sci. 2023, 24, 3042. https://doi.org/10.3390/ijms24033042
Pustovalova M, Malakhov P, Guryanova A, Sorokin M, Suntsova M, Buzdin A, Osipov AN, Leonov S. Transcriptome-Based Traits of Radioresistant Sublines of Non-Small Cell Lung Cancer Cells. International Journal of Molecular Sciences. 2023; 24(3):3042. https://doi.org/10.3390/ijms24033042
Chicago/Turabian StylePustovalova, Margarita, Philipp Malakhov, Anastasia Guryanova, Maxim Sorokin, Maria Suntsova, Anton Buzdin, Andreyan N. Osipov, and Sergey Leonov. 2023. "Transcriptome-Based Traits of Radioresistant Sublines of Non-Small Cell Lung Cancer Cells" International Journal of Molecular Sciences 24, no. 3: 3042. https://doi.org/10.3390/ijms24033042