
Dysbiosis of Fecal Microbiota in Tg2576 Mice for Alzheimer’s Disease during Pathological Constipation
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Confirmation of the Constipation Phenotypes in Tg2576 Mice
2.2. Alteration of the Profile of the Fecal Microbiota in Tg2576 Mice with the Constipation Phenotypes
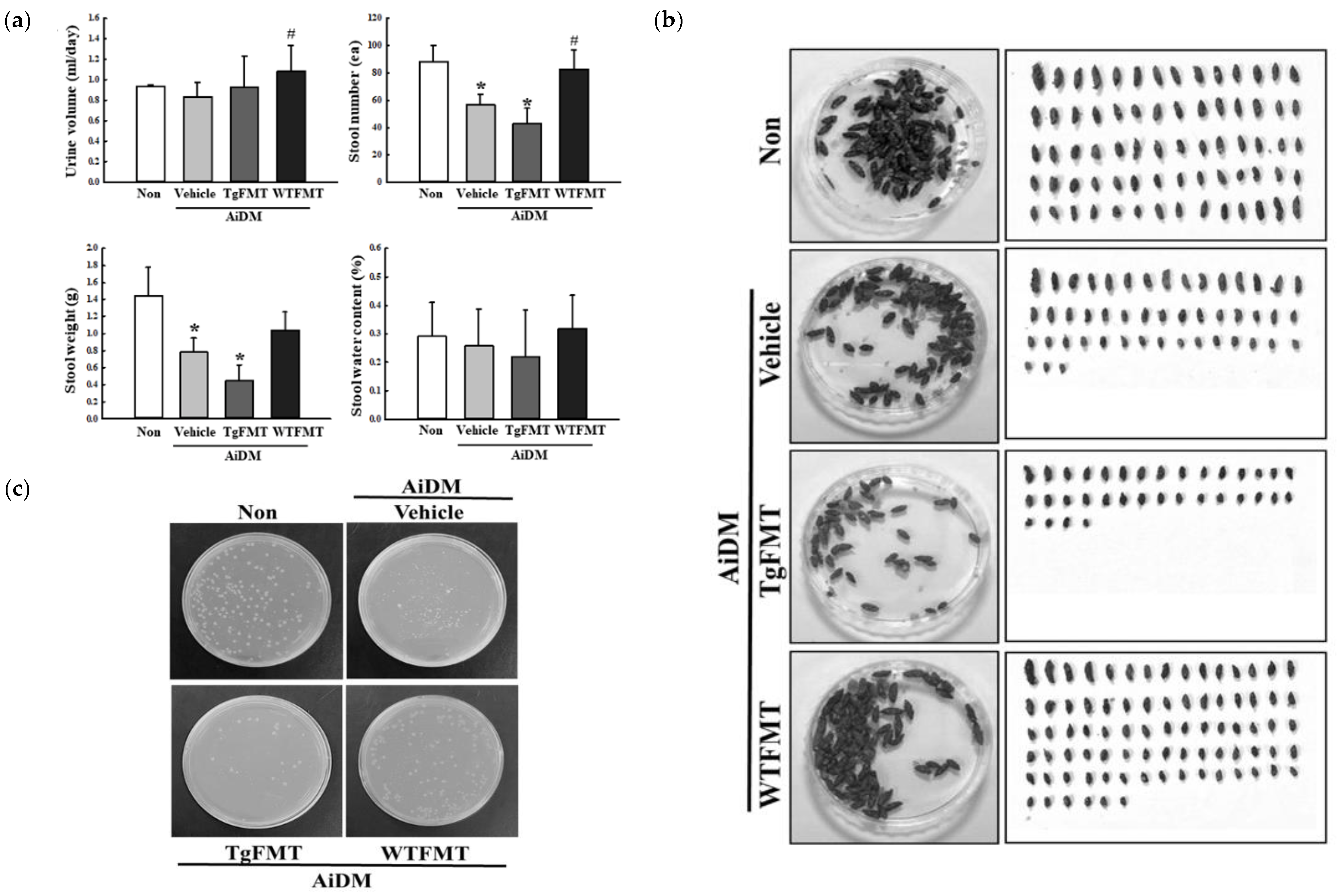
2.3. Effects of TgFMT on Fecal Parameters of AiDM-ICR Mice
2.4. Effects of TgFMT on the Colon Length and Gastrointestinal (GI) Motility in AiDM-ICR Mice
2.5. Effects of TgFMT on the Histopathological Structure of the Mid Colon in AiDM-ICR Mice
2.6. Effects of TgFMT on the Expression of Water Channels in the Mid Colon of AiDM-ICR Mice
2.7. Effects of TgFMT on the Distributions of Neuronal Cells and ICC in the Mid Colon of AiDM-ICR Mice
2.8. Effects of TgFMT on the Excitatory Function of the ENS in the Mid Colon of AiDM-ICR Mice
3. Discussion
4. Materials and Methods
4.1. Management of Animal Study
4.2. Experimental Design of the Tg2576 Mice
4.3. Excretion Parameters Analyses
4.4. Measurement of GI Transit Ratio and Colon Length
4.5. Histopathological Analysis of Mid Colon
4.6. Analysis of Fecal Microbiota
4.7. FMT Analysis
4.8. Culture of Fecal-Derived Microbiota
4.9. Quantitative Real-Time PCR Analysis (RT-qPCR)
4.10. Western Blotting Analysis
4.11. Measurement of 5-HT Concentrations
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Edwards, L.L.; Quigley, E.M.; Harned, R.K.; Hofman, R.; Pfeiffer, R.F. Characterization of swallowing and defecation in Parkinson’s disease. Am. J. Gastroenterol. 1994, 89, 15–25. [Google Scholar]
- Bakke, A.; Myhr, K.M.; Gronning, M.; Nyland, H. Bladder, bowel and sexual dysfunction in patients with multiple sclerosis—A cohort study. Scand. J. Urol. Nephrol. Suppl. 1996, 179, 61–66. [Google Scholar]
- Binnie, N.R.; Smith, A.N.; Creasey, G.H.; Edmond, P. Constipation associated with chronic spinal cord injury: The effect of pelvic parasympathetic stimulation by the Brindley stimulator. Paraplegia 1991, 29, 463–469. [Google Scholar] [CrossRef]
- Zakrzewska-Pniewska, B.; Gawel, M.; Szmidt-Salkowska, E.; Kepczynska, K.; Nojszewska, M. Clinical and functional assessment of dysautonomia and its correlation in Alzheimer’s disease. Am. J. Alzheimers Dis. Other Demen. 2012, 27, 592–599. [Google Scholar] [CrossRef]
- Ullman, T.; Reding, M. Gastrointestinal dysfunction in stroke. Semin. Neurol. 1996, 16, 269–275. [Google Scholar] [CrossRef]
- Jost, W.H. Gastrointestinal motility problems in patients with Parkinson’s disease. Effects of antiparkinsonian treatment and guidelines for management. Drugs Aging 1997, 10, 249–258. [Google Scholar] [CrossRef]
- Kim, J.E.; Park, J.J.; Lee, M.R.; Choi, J.Y.; Song, B.R.; Park, J.W.; Kang, M.J.; Son, H.J.; Hong, J.T.; Hwang, D.Y. Constipation in Tg2576 mice model for Alzheimer’s disease associated with dysregulation of mechanism involving the mAChR signaling pathway and ER stress response. PLoS ONE 2019, 14, e0215205. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.H.; Chang, S.K. Alteration of gut microbiota and efficacy of probiotics in functional constipation. J. Neurogastroenterol. Motil. 2015, 21, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yu, Y.B. Intestinal microbiota and chronic constipation. Springer Plus 2016, 5, 1130. [Google Scholar] [CrossRef] [Green Version]
- Zoppi, G.; Cinquetti, M.; Luciano, A.; Benini, A.; Muner, A.; Bertazzoni, M. The intestinal ecosystem in chronic functional constipation. Acta Paediatr. 1998, 87, 836–841. [Google Scholar] [CrossRef]
- Gerritsen, J.; Smidt, H.; Rijkers, G.T.; de Vos, W.M. Intestinal microbiota in human health and disease: The impact of probiotics. Genes Nutr. 2011, 6, 209–240. [Google Scholar] [CrossRef] [Green Version]
- Kirgizov, I.V.; Sukhorukov, A.M.; Dudarev, V.A.; Istomin, A.A. Hemostasis in children with dysbacteriosis in chronic constipation. Clin. Appl. Thromb. Hemost. 2001, 7, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Khalif, I.L.; Quigley, E.M.; Konovitch, E.A.; Maximova, I.D. Alterations in the colonic flora and intestinal permeability and evidence of immune activation in chronic constipation. Dig. Liver Dis. 2005, 37, 838–849. [Google Scholar] [CrossRef]
- Kim, S.E.; Choi, S.C.; Park, K.S.; Park, M.I.; Shin, J.E.; Lee, T.H.; Jung, K.W.; Koo, H.S.; Myung, S.J. Change of fecal flora and effectiveness of the short-term VSL#3 probiotic treatment in patients with functional constipation. J. Neurofastroenterol. Motil. 2015, 21, 111–120. [Google Scholar]
- Simren, M.; Barbara, G.; Flint, H.J.; Spiegel, B.M.; Spiller, R.C.; Vanner, S.; Verdu, E.F.; Whorwell, P.J.; Zoetendal, E.G.; Committee, R.F. Intestinal microbiota in functional bowel disorders: A Rome foundation report. Gut 2013, 62, 159–176. [Google Scholar] [CrossRef] [PubMed]
- Nourrisson, C.; Scanzi, J.; Pereira, B.; NkoudMongo, C.; Wawrzyniak, I.; Cian, A.; Viscogliosi, E.; Livrelli, V.; Delbac, F.; Dapoigny, M.; et al. Blastocystis is associated with decrease of fecal microbiota protective bacteria: Comparative analysis between patients with irritable bowel syndrome and control subjects. PLoS ONE 2014, 9, e111868. [Google Scholar] [CrossRef]
- Zhu, L.; Liu, W.; Alkhouri, R.; Baker, R.D.; Bard, J.E.; Quigley, E.M.; Baker, S.S. Structural changes in the gut microbiome of constipated patients. Physiol. Genomics 2014, 46, 679–686. [Google Scholar]
- Pryde, S.E.; Duncan, S.H.; Hold, G.L.; Stewart, C.S.; Flint, H.J. The microbiology of butyrate formation in the human colon. FEMS Microbiol. Lett. 2002, 217, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermudez, H.L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Meng, J.; Zhang, L.; Johnson, T.; Chen, C.; Roy, S. Morphine induces changes in the gut microbiome and metabolome in a morphine dependence model. Sci. Rep. 2018, 8, 3596. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.G.; Wen, P.; Fu, H.Z.; Lin, G.Y.; Liao, S.T.; Zou, Y.X. Protective effect of mulberry (Morus atropurpurea) fruit against diphenoxylate-induced constipation in mice through the modulation of gut microbiota. Food Funct. 2019, 10, 1513–1528. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, H.; Zheng, J.; Jiang, N.; Sun, G.; Bao, X.; Lin, A.; Liu, H. Chitosan oligosaccharides attenuate loperamide-induced constipation through regulation of gut microbiota in mice. Carbohydr. Polym. 2021, 253, 117218. [Google Scholar] [CrossRef]
- Zhang, X.; Zheng, J.; Jiang, N.; Sun, G.; Bao, X.; Kong, M.; Cheng, X.; Lin, A.; Liu, H. Modulation of gut microbiota and intestinal metabolites by lactulose improves loperamide-induced constipation in mice. Eur. J. Pharm. Sci. 2021, 158, 105676. [Google Scholar] [CrossRef]
- Li, Y.; Long, S.; Liu, Q.; Ma, H.; Li, J.; Xiaoqing, W.; Yuan, J.; Li, M.; Hou, B. Gut microbiota is involved in the alleviation of loperamide-induced constipation by honey supplementation in mice. Food Sci. Nutr. 2020, 8, 4388–4398. [Google Scholar] [CrossRef]
- Suo, H.; Zhao, X.; Qian, Y.; Li, G.; Liu, Z.; Xie, J.; Li, J. Therapeutic effect of activated carbon-induced constipation mice with Lactobacillus fermentum Suo on treatment. Int. J. Mol. Sci. 2014, 15, 21875–21895. [Google Scholar] [CrossRef]
- Park, J.W.; Kim, J.E.; Choi, Y.J.; Kang, M.J.; Choi, H.J.; Bae, S.J.; Hong, J.T.; Lee, H.; Hwang, D.Y. Deficiency of complement component 3 may be linked to the development of constipation in FVB/N-C3(em1Hlee) /Korl mice. FASEB J. 2021, 35, e21221. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kim, J.E.; Lee, S.J.; Gong, J.E.; Son, H.J.; Hong, J.T.; Hwang, D.Y. Dysbiosis of fecal microbiota from complement 3 knockout mice with constipation phenotypes contributes to development of defecation delay. Front. Physiol. 2021, 12, 650789. [Google Scholar] [CrossRef]
- Quigley, E.M.M. Microflora modulation of motility. J. Neurogastroenterol. Motil. 2011, 17, 140–147. [Google Scholar] [CrossRef]
- Sommer, F.; Backhed, F. The gut microbiota-masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef]
- Hsaio, K.; Chapman, P.; Nilsen, S.; Eckman, C.; Harigaya, Y.; Younkin, S.; Yang, F.; Cole, G. Correlative memory deficits, Aβ elevation, and amyloid plaques in transgenic mice. Science 1996, 274, 99–102. [Google Scholar] [CrossRef]
- Bilkei-Gorzo, A. Genetic mouse models of brain ageing and Alzheimer’s disease. Pharm. Ther. 2014, 142, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Townsend, M.; Mehta, T.; Selkoe, D.J. Soluble Abeta inhibits specific signal transduction cascades common to the insulin receptor pathway. J. Biol. Chem. 2007, 282, 33305–33312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohjima, M.; Sun, Y.; Chan, L. Increased food intake leads to obesity and insulin resistance in the Tg2576 Alzheimer’s disease mouse model. Endocrinology 2010, 151, 1532–1540. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Cao, D.; Garber, D.W.; Kim, H.; Fukuchi, K. Association of aortic atherosclerosis with cerebral beta-amyloidosis and learning deficits in a mouse model of Alzheimer’s disease. Am. J. Pathol. 2003, 163, 2155–2164. [Google Scholar] [CrossRef]
- van Olst, L.; Roks, S.J.M.; Kamermans, A.; Verhaar, B.J.H.; van der Geest, A.M.; Muller, M.; van der Flier, W.M.; de Vries, H.E. Contribution of gut microbiota to immunological changes in Alzheimer’s disease. Front. Immunol. 2021, 12, 683068. [Google Scholar] [CrossRef]
- Cox, L.M.; Schafer, M.J.; Sohn, J.; Vincentini, J.; Weiner, H.L.; Ginsberg, S.D.; Blaser, M.J. Calorie restriction slows age-related microbiota changes in an Alzheimer’s disease model in female mice. Sci. Rep. 2019, 9, 17904. [Google Scholar] [CrossRef] [Green Version]
- Madigan, M.T.; Martinko, J.M.; Brock, T.D. Brock Biology of Microorganisms, 11th ed.; Prentice Hall: Boston, MA, USA, 2005. [Google Scholar]
- Ryan, K.J.; Ray, C.G. Sherris Medical Microbiology, 4th ed.; McGraw Hill: New York, NY, USA, 2004. [Google Scholar]
- Choi, J.S.; Kim, J.W.; Cho, H.R.; Kim, K.Y.; Lee, J.K.; Sohn, J.H.; Ku, S.K. Laxative effects of fermented rice extract in rats with loperamide-induced constipation. Exp. Med. 2014, 8, 1847–1854. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.E.; Choi, Y.J.; Lee, S.J.; Gong, J.E.; Lee, Y.J.; Sung, J.E.; Jung, Y.S.; Lee, H.S.; Hong, J.T.; Hwang, D.Y. Antioxidant activity and laxative effects of tannin-enriched extract of Ecklonia cava in loperamide-induced constipation of SD rats. PLoS ONE 2021, 16, e0246363. [Google Scholar] [CrossRef]
- Kim, J.E.; Choi, Y.J.; Lee, S.J.; Gong, J.E.; Jin, Y.J.; Park, S.H.; Lee, H.S.; Choi, Y.W.; Hong, J.T.; Hwang, D.Y. Laxative effects of phlorotannins derived from Ecklonia cava on loperamide-induced constipation in SD rats. Molecules 2021, 26, 7209. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. J. Bioinform. 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Li, W.; Fu, L.; Niu, B.; Wu, S.; Wooley, J. Ultrafast clustering algorithms for metagenomic sequence analysis. J. Bioinform. 2012, 13, 656–668. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, H.; Liu, X.; An, Y.; Zhou, G.; Liu, Y.; Xu, M.; Dong, W.; Wang, S.; Yan, F.; Jiang, K.; et al. Dysbiosis contributes to chronic constipation development via regulation of serotonin transporter in the intestine. Sci. Rep. 2017, 7, 10322. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C (T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-E.; Roh, Y.-J.; Choi, Y.-J.; Lee, S.-J.; Jin, Y.-J.; Song, H.-J.; Seol, A.-Y.; Son, H.-J.; Hong, J.-T.; Hwang, D.-Y. Dysbiosis of Fecal Microbiota in Tg2576 Mice for Alzheimer’s Disease during Pathological Constipation. Int. J. Mol. Sci. 2022, 23, 14928. https://doi.org/10.3390/ijms232314928
Kim J-E, Roh Y-J, Choi Y-J, Lee S-J, Jin Y-J, Song H-J, Seol A-Y, Son H-J, Hong J-T, Hwang D-Y. Dysbiosis of Fecal Microbiota in Tg2576 Mice for Alzheimer’s Disease during Pathological Constipation. International Journal of Molecular Sciences. 2022; 23(23):14928. https://doi.org/10.3390/ijms232314928
Chicago/Turabian StyleKim, Ji-Eun, Yu-Jeong Roh, Yun-Ju Choi, Su-Jin Lee, You-Jeong Jin, Hee-Jin Song, A-Yun Seol, Hong-Joo Son, Jin-Tae Hong, and Dae-Youn Hwang. 2022. "Dysbiosis of Fecal Microbiota in Tg2576 Mice for Alzheimer’s Disease during Pathological Constipation" International Journal of Molecular Sciences 23, no. 23: 14928. https://doi.org/10.3390/ijms232314928