
Reversal of Peripheral Neuropathic Pain by the Small-Molecule Natural Product Narirutin via Block of Nav1.7 Voltage-Gated Sodium Channel

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Narirutin Shows Antinociceptive Effects in SNI-Induced Rat Models of Neuropathic Pain
2.2. Narirutin Inhibited Veratridine-Triggered Nociceptor Activities of DRG Neurons
2.3. Total Sodium Current (INa) in DRG Sensory Neurons Is Reduced by Narirutin Treatment
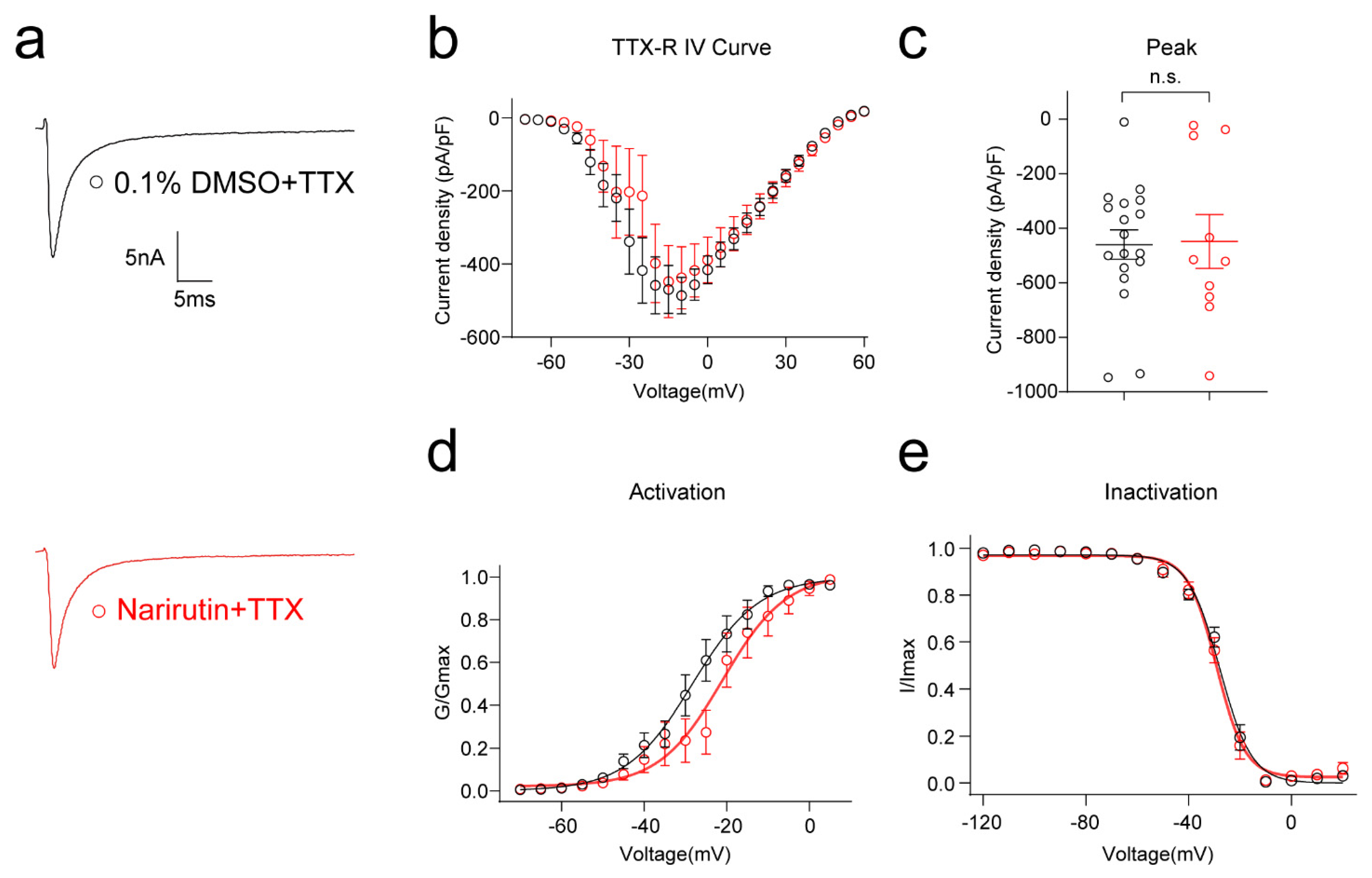
2.4. Narirutin Inhibits TTX-Sensitive Rather Than TTX-Resistant Sodium Currents in DRG Sensory Neurons
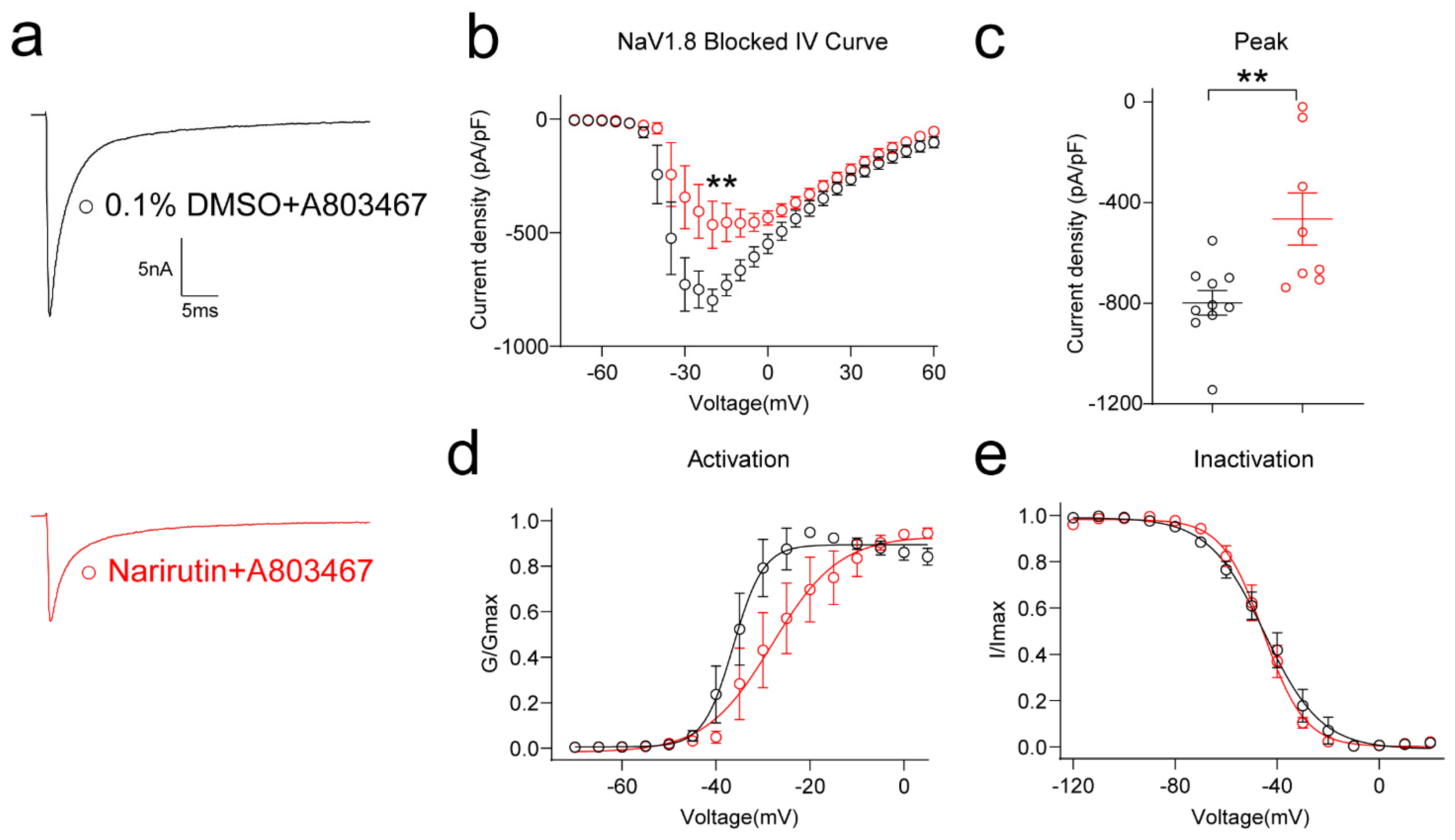
2.5. Narirutin Specifically Inhibits Nav1.7 Currents in Both DRG Sensory Neurons and HEK293 Cell Lines
2.6. Narirutin Blunts the Upregulation of Nav1.7 in CGRP+ DRG Sensory Neurons after SNI Surgery
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Reagents
4.3. Spared Nerve Injury (SNI)
4.4. Assessment of Mechanical Allodynia Using the Von Frey Test
4.5. Assessment of Cold Allodynia Using the Acetone Test
4.6. Cell Culture and Transfection
4.6.1. Cell Culture
4.6.2. Transfection
4.7. Calcium Imaging and Analysis
4.8. Whole-Cell Voltage Clamp Electrophysiology
4.9. Immunoblotting
4.10. Immunofluorescence
4.11. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jensen, T.S.; Baron, R.; Haanpaa, M.; Kalso, E.; Loeser, J.D.; Rice, A.S.C.; Treede, R.D. A new definition of neuropathic pain. Pain 2011, 152, 2204–2205. [Google Scholar] [CrossRef] [PubMed]
- Krebs, E.E.; Gravely, A.; Nugent, S.; Jensen, A.C.; DeRonne, B.; Goldsmith, E.S.; Kroenke, K.; Bair, M.J.; Noorbaloochi, S. Effect of Opioid vs Nonopioid Medications on Pain-Related Function in Patients with Chronic Back Pain or Hip or Knee Osteoarthritis Pain: The SPACE Randomized Clinical Trial. JAMA 2018, 319, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.; Tse, W.C.; Larance, B. Opioid agonist treatment for people who are dependent on pharmaceutical opioids. Cochrane Database Syst. Rev. 2022, 9, CD011117. [Google Scholar] [PubMed]
- Khan, F.A.; Ali, G.; Rahman, K.; Khan, Y.; Ayaz, M.; Mosa, O.F.; Nawaz, A.; Hassan, S.S.U.; Bungau, S. Efficacy of 2-Hydroxyflavanone in Rodent Models of Pain and Inflammation: Involvement of Opioidergic and GABAergic Anti-Nociceptive Mechanisms. Molecules 2022, 27, 5431. [Google Scholar] [CrossRef] [PubMed]
- Khatoon, F.; Ali, S.; Kumar, V.; Elasbali, A.M.; Alhassan, H.H.; Alharethi, S.H.; Islam, A.; Hassan, M.I. Pharmacological features, health benefits and clinical implications of honokiol. J. Biomol. Struct. Dyn. 2022, 1–23. [Google Scholar] [CrossRef]
- Friedrich, L.; Cingolani, G.; Ko, Y.H.; Iaselli, M.; Miciaccia, M.; Perrone, M.G.; Neukirch, K.; Bobinger, V.; Merk, D.; Hofstetter, R.K.; et al. Learning from Nature: From a Marine Natural Product to Synthetic Cyclooxygenase-1 Inhibitors by Automated De Novo Design. Adv. Sci. 2021, 8, e2100832. [Google Scholar] [CrossRef]
- Haroutounian, S.; Nikolajsen, L.; Bendtsen, T.F.; Finnerup, N.B.; Kristensen, A.D.; Hasselstrom, J.B.; Jensen, T.S. Primary afferent input critical for maintaining spontaneous pain in peripheral neuropathy. Pain 2014, 155, 1272–1279. [Google Scholar] [CrossRef]
- Bajaj, S.; Ong, S.T.; Chandy, K.G. Contributions of natural products to ion channel pharmacology. Nat. Prod. Rep. 2020, 37, 703–716. [Google Scholar] [CrossRef]
- Cardoso, F.C.; Lewis, R.J. Sodium channels and pain: From toxins to therapies. Br. J. Pharmacol. 2018, 175, 2138–2157. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Li, T.; Rohou, A.; Arthur, C.P.; Tzakoniati, F.; Wong, E.; Estevez, A.; Kugel, C.; Franke, Y.; Chen, J.; et al. Structural Basis of Nav1.7 Inhibition by a Gating-Modifier Spider Toxin. Cell 2019, 176, 702–715.e14. [Google Scholar] [CrossRef]
- Shen, H.; Liu, D.; Wu, K.; Lei, J.; Yan, N. Structures of human Nav1.7 channel in complex with auxiliary subunits and animal toxins. Science 2019, 363, 1303–1308. [Google Scholar] [CrossRef]
- Wu, Y.; Zou, B.; Liang, L.; Li, M.; Tao, Y.X.; Yu, H.; Wang, X.; Li, M. Loperamide inhibits sodium channels to alleviate inflammatory hyperalgesia. Neuropharmacology 2017, 117, 282–291. [Google Scholar] [CrossRef]
- Foudah, A.I.; Alqarni, M.H.; Devi, S.; Singh, A.; Alam, A.; Alam, P.; Singh, S. Analgesic Action of Catechin on Chronic Constriction Injury-Induced Neuropathic Pain in Sprague-Dawley Rats. Front. Pharmacol. 2022, 13, 895079. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.S.; Soto Tellini, V.H.; Benjumea Gutierrez, D.M. Study of the dermal anti-inflammatory, antioxidant, and analgesic activity of pinostrobin. Heliyon 2022, 8, e10413. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, D.Q.; Tian, Y.K.; Mei, W.; Tian, X.B.; Xu, A.J.; Zhou, Y.Q. The emerging role of quercetin in the treatment of chronic pain. Curr. Neuropharmacol. 2022, 20, 2346–2353. [Google Scholar]
- Zhou, Y.; Cai, S.; Moutal, A.; Yu, J.; Gomez, K.; Madura, C.L.; Shan, Z.; Pham, N.Y.N.; Serafini, M.J.; Dorame, A.; et al. The Natural Flavonoid Naringenin Elicits Analgesia through Inhibition of NaV1.8 Voltage-Gated Sodium Channels. ACS Chem. Neurosci. 2019, 10, 4834–4846. [Google Scholar] [CrossRef] [PubMed]
- Shammugasamy, B.; Valtchev, P.; Dong, Q.; Dehghani, F. Effect of citrus peel extracts on the cellular quiescence of prostate cancer cells. Food Funct. 2019, 10, 3727–3737. [Google Scholar] [CrossRef] [PubMed]
- Islam, F.; Mitra, S.; Nafady, M.H.; Rahman, M.T.; Tirth, V.; Akter, A.; Emran, T.B.; Mohamed, A.A.; Algahtani, A.; El-Kholy, S.S. Neuropharmacological and Antidiabetic Potential of Lannea Coromandelica (Houtt.) Merr. Leaves Extract: An Experimental Analysis. Evid. Based Complement. Altern. Med. 2022, 2022, 6144733. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.K.; Park, H.Y.; Eom, H.; Kim, Y.; Choi, I. Narirutin fraction from citrus peels attenuates LPS-stimulated inflammatory response through inhibition of NF-kappaB and MAPKs activation. Food Chem. Toxicol. 2012, 50, 3498–3504. [Google Scholar] [CrossRef]
- Camarda, L.; Di Stefano, V.; Del Bosco, S.F.; Schillaci, D. Antiproliferative activity of citrus juices and HPLC evaluation of their flavonoid composition. Fitoterapia 2007, 78, 426–429. [Google Scholar] [CrossRef]
- Mitra, S.; Lami, M.S.; Uddin, T.M.; Das, R.; Islam, F.; Anjum, J.; Hossain, M.J.; Emran, T.B. Prospective multifunctional roles and pharmacological potential of dietary flavonoid narirutin. Biomed. Pharmacother. 2022, 150, 112932. [Google Scholar] [CrossRef] [PubMed]
- Decosterd, I.; Woolf, C.J. Spared nerve injury: An animal model of persistent peripheral neuropathic pain. Pain 2000, 87, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Eijkenboom, I.; Sopacua, M.; Hoeijmakers, J.G.J.; de Greef, B.T.A.; Lindsey, P.; Almomani, R.; Marchi, M.; Vanoevelen, J.; Smeets, H.J.M.; Waxman, S.G.; et al. Yield of peripheral sodium channels gene screening in pure small fibre neuropathy. J. Neurol. Neurosurg. Psychiatry 2019, 90, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Fouillet, A.; Watson, J.F.; Piekarz, A.D.; Huang, X.; Li, B.; Priest, B.; Nisenbaum, E.; Sher, E.; Ursu, D. Characterisation of Nav1.7 functional expression in rat dorsal root ganglia neurons by using an electrical field stimulation assay. Mol. Pain 2017, 13, 1744806917745179. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Ding, X.; Wang, T.; Mou, S.; Sun, H.; Hou, T. Voltage-gated sodium channels: Structures, functions, and molecular modeling. Drug Discov. Today 2019, 24, 1389–1397. [Google Scholar] [CrossRef]
- Mohammed, Z.A.; Kaloyanova, K.; Nassar, M.A. An unbiased and efficient assessment of excitability of sensory neurons for analgesic drug discovery. Pain 2020, 161, 1100–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammed, Z.A.; Doran, C.; Grundy, D.; Nassar, M.A. Veratridine produces distinct calcium response profiles in mouse dorsal root ganglia neurons. Sci. Rep. 2017, 7, 45221. [Google Scholar] [CrossRef] [Green Version]
- Dustrude, E.T.; Moutal, A.; Yang, X.; Wang, Y.; Khanna, M.; Khanna, R. Hierarchical CRMP2 posttranslational modifications control NaV1.7 function. Proc. Natl. Acad. Sci. USA 2016, 113, E8443–E8452. [Google Scholar] [CrossRef] [Green Version]
- Akopian, A.N.; Souslova, V.; England, S.; Okuse, K.; Ogata, N.; Ure, J.; Smith, A.; Kerr, B.J.; McMahon, S.B.; Boyce, S.; et al. The tetrodotoxin-resistant sodium channel SNS has a specialized function in pain pathways. Nat. Neurosci. 1999, 2, 541–548. [Google Scholar] [CrossRef]
- Renganathan, M.; Cummins, T.R.; Waxman, S.G. Contribution of Na(v)1.8 sodium channels to action potential electrogenesis in DRG neurons. J. Neurophysiol. 2001, 86, 629–640. [Google Scholar] [CrossRef] [Green Version]
- Rush, A.M.; Cummins, T.R.; Waxman, S.G. Multiple sodium channels and their roles in electrogenesis within dorsal root ganglion neurons. J. Physiol. 2007, 579, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Bean, B.P. Lidocaine Binding Enhances Inhibition of Nav1.7 Channels by the Sulfonamide PF-05089771. Mol. Pharmacol. 2020, 97, 377–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akin, E.J.; Alsaloum, M.; Higerd, G.P.; Liu, S.; Zhao, P.; Dib-Hajj, F.B.; Waxman, S.G.; Dib-Hajj, S.D. Paclitaxel increases axonal localization and vesicular trafficking of Nav1.7. Brain 2021, 144, 1727–1737. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhu, L.; Zhang, W.; Tang, Q.; Zhong, Y. IL-10 alleviates radicular pain by inhibiting TNF-alpha/p65 dependent Nav1.7 up-regulation in DRG neurons of rats. Brain Res. 2022, 1791, 147997. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Suzuki, T.; Karnup, S.V.; Gu, B.; Yoshimura, N. Nerve growth factor-mediated Na(+) channel plasticity of bladder afferent neurons in mice with spinal cord injury. Life Sci. 2022, 298, 120524. [Google Scholar] [CrossRef]
- Li, Y.; North, R.Y.; Rhines, L.D.; Tatsui, C.E.; Rao, G.; Edwards, D.D.; Cassidy, R.M.; Harrison, D.S.; Johansson, C.A.; Zhang, H.; et al. DRG Voltage-Gated Sodium Channel 1.7 Is Upregulated in Paclitaxel-Induced Neuropathy in Rats and in Humans with Neuropathic Pain. J. Neurosci. 2018, 38, 1124–1136. [Google Scholar] [CrossRef] [Green Version]
- Laedermann, C.J.; Cachemaille, M.; Kirschmann, G.; Pertin, M.; Gosselin, R.D.; Chang, I.; Albesa, M.; Towne, C.; Schneider, B.L.; Kellenberger, S.; et al. Dysregulation of voltage-gated sodium channels by ubiquitin ligase NEDD4-2 in neuropathic pain. J. Clin. Investig. 2013, 123, 3002–3013. [Google Scholar] [CrossRef] [Green Version]
- Franken, G.; Douven, P.; Debets, J.; Joosten, E.A.J. Conventional Dorsal Root Ganglion Stimulation in an Experimental Model of Painful Diabetic Peripheral Neuropathy: A Quantitative Immunocytochemical Analysis of Intracellular Gamma-Aminobutyric Acid in Dorsal Root Ganglion Neurons. Neuromodulation 2021, 24, 639–645. [Google Scholar] [CrossRef]
- Noguri, T.; Hatakeyama, D.; Kitahashi, T.; Oka, K.; Ito, E. Profile of dorsal root ganglion neurons: Study of oxytocin expression. Mol. Brain 2022, 15, 44. [Google Scholar] [CrossRef]
- Xu, J.; Wu, S.; Wang, J.; Wang, J.; Yan, Y.; Zhu, M.; Zhang, D.; Jiang, C.; Liu, T. Oxidative stress induced by NOX2 contributes to neuropathic pain via plasma membrane translocation of PKCepsilon in rat dorsal root ganglion neurons. J. Neuroinflamm. 2021, 18, 106. [Google Scholar] [CrossRef]
- Cohen, F.; Yuan, H.; Silberstein, S.D. Calcitonin Gene-Related Peptide (CGRP)-Targeted Monoclonal Antibodies and Antagonists in Migraine: Current Evidence and Rationale. BioDrugs 2022, 36, 341–358. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Lauritsen, C.G.; Kaiser, E.A.; Silberstein, S.D. CGRP Monoclonal Antibodies for Migraine: Rationale and Progress. BioDrugs 2017, 31, 487–501. [Google Scholar] [CrossRef]
- Rabiller, L.; Labit, E.; Guissard, C.; Gilardi, S.; Guiard, B.P.; Mouledous, L.; Silva, M.; Mithieux, G.; Penicaud, L.; Lorsignol, A.; et al. Pain sensing neurons promote tissue regeneration in adult mice. NPJ Regen. Med. 2021, 6, 63. [Google Scholar] [CrossRef] [PubMed]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Roth, E.; Linehan, A.; Weihrauch, D.; Stucky, C.; Hogan, Q.; Hoben, G. Targeted muscle reinnervation prevents and reverses rat pain behaviors following nerve transection. Pain 2022. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Jin, X.; Huang, G.; Huang, J.; Wu, T.; Li, Z.; Chen, J.; Kong, F.; Pan, X.; Yan, N. Structural basis for high-voltage activation and subtype-specific inhibition of human Nav1.8. Proc. Natl. Acad. Sci. USA 2022, 119, e2208211119. [Google Scholar] [CrossRef]
- Chernov-Rogan, T.; Li, T.; Lu, G.; Verschoof, H.; Khakh, K.; Jones, S.W.; Beresini, M.H.; Liu, C.; Ortwine, D.F.; McKerrall, S.J.; et al. Mechanism-specific assay design facilitates the discovery of Nav1.7-selective inhibitors. Proc. Natl. Acad. Sci. USA 2018, 115, E792–E801. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total NaV (Narirutin-20 μM) | 0.1% DMSO | Narirutin | p Value 2 | |
| Treated | Overnight | Overnight | ||
| Activation | V1/2 | −39.2 ± 0.7 | −28.8 ± 0.7 | **** p < 0.0001 |
| k | 3.0 ± 0.6 | 3.7 ± 0.6 | p = 0.4585 | |
| Inactivation | V1/2 | −39.5 ± 2.0 | −50.3 ± 2.0 | ** p = 0.0013 |
| k | −14.9 ± 1.9 | −16.0 ± 2.0 | p = 0.7018 | |
| Blocked TTX-S (TTX-1 μM) | 0.1% DMSO + TTX | Narirutin + TTX | ||
| Treated | Another Overnight TTX Acute | Another Overnight TTX Acute | ||
| Activation | V1/2 | −28.3 ± 1.2 | −21.0 ± 1.9 | ** p = 0.0045 |
| k | 8.0 ± 1.2 | 7.7 ± 1.7 | p = 0.8655 | |
| Inactivation | V1/2 | −27.8 ± 0.5 | −29.1 ± 0.6 | p = 0.1025 |
| k | −6.2 ± 0.4 | −5.6 ± 0.5 | p = 0.3450 | |
| Blocked Nav1.8 (A-803467-500 nM) | 0.1%DMSO + A-803467 | Narirutin + A-803467 | ||
| Treated | Another Overnight A-803467 Acute | Another Overnight A-803467 Acute | ||
| Activation | V1/2 | −36.3 ± 0.8 | −28.0 ± 1.9 | ** p = 0.0026 |
| k | 3.2 ± 0.7 | 7.1 ± 1.8 | p = 0.0681 | |
| Inactivation | V1/2 | −44.9 ± 1.2 | −45.3 ± 0.8 | p = 0.7653 |
| k | −11.1 ± 1.1 | −8.1 ± 0.7 | * p = 0.0412 | |
| Blocked Nav1.7 (PF-05089771-100 nM) | 0.1%DMSO + PF-05089771 | Narirutin + PF-05089771 | ||
| Treated | Another Overnight PF-05089771 Acute | Another Overnight PF-05089771 Acute | ||
| Activation | V1/2 | −23.8 ± 0.9 | −24.9 ± 0.6 | p = 0.3620 |
| k | 5.5 ± 0.8 | 3.9 ± 0.6 | p = 0.1084 | |
| Inactivation | V1/2 | −35.8 ± 1.1 | −39.9 ± 1.0 | * p = 0.0398 |
| k | −9.6 ± 0.9 | −10.8 ± 0.9 | p = 0.3740 | |
| Nav1.7 (Narirutin 20 μM) | 0.1% DMSO | Narirutin (20 μM) | ||
| Treated | Overnight | Overnight | ||
| Activation | V1/2 | −15.2 ± 0.4 | −8.0 ± 2.9 | * p = 0.0342 |
| k | 4.1 ± 0.3 | 8.3 ± 1.6 | * p = 0.0295 | |
| Inactivation | V1/2 | −66.5 ± 0.6 | −65.8 ± 1.1 | p = 0.5900 |
| k | −7.4 ± 0.6 | −7.5 ± 0.9 | p = 0.9140 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Shan, Z.; Guo, W.; Wang, Y.; Cai, S.; Li, F.; Huang, Q.; Liu, J.A.; Cheung, C.W.; Cai, S. Reversal of Peripheral Neuropathic Pain by the Small-Molecule Natural Product Narirutin via Block of Nav1.7 Voltage-Gated Sodium Channel. Int. J. Mol. Sci. 2022, 23, 14842. https://doi.org/10.3390/ijms232314842
Yang H, Shan Z, Guo W, Wang Y, Cai S, Li F, Huang Q, Liu JA, Cheung CW, Cai S. Reversal of Peripheral Neuropathic Pain by the Small-Molecule Natural Product Narirutin via Block of Nav1.7 Voltage-Gated Sodium Channel. International Journal of Molecular Sciences. 2022; 23(23):14842. https://doi.org/10.3390/ijms232314842
Chicago/Turabian StyleYang, Haoyi, Zhiming Shan, Weijie Guo, Yuwei Wang, Shuxian Cai, Fuyi Li, Qiaojie Huang, Jessica Aijia Liu, Chi Wai Cheung, and Song Cai. 2022. "Reversal of Peripheral Neuropathic Pain by the Small-Molecule Natural Product Narirutin via Block of Nav1.7 Voltage-Gated Sodium Channel" International Journal of Molecular Sciences 23, no. 23: 14842. https://doi.org/10.3390/ijms232314842