
Cloning and Molecular Characterization of Hsp Genes from Anoplophora glabripennis and Their Responses to Cold Stress

Abstract
:1. Introduction
2. Results
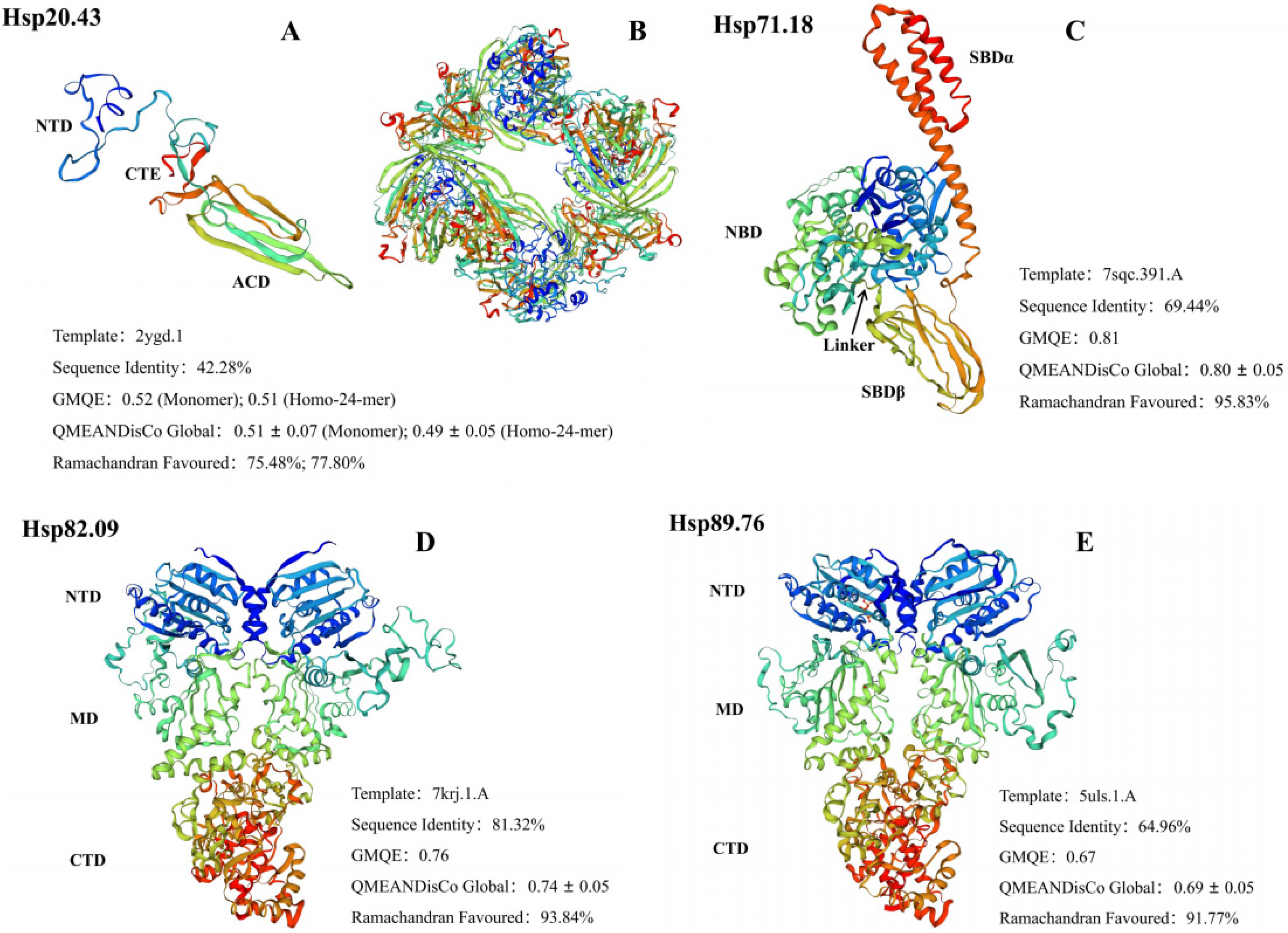
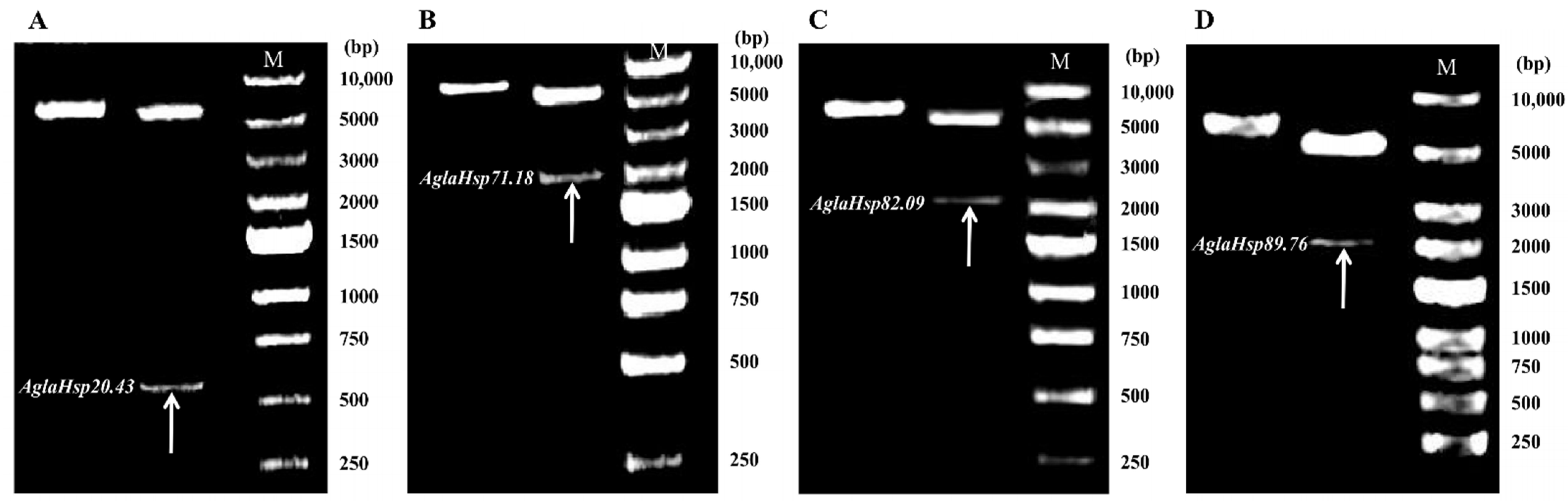
2.1. Cloning and Sequence Analysis of AglaHsps
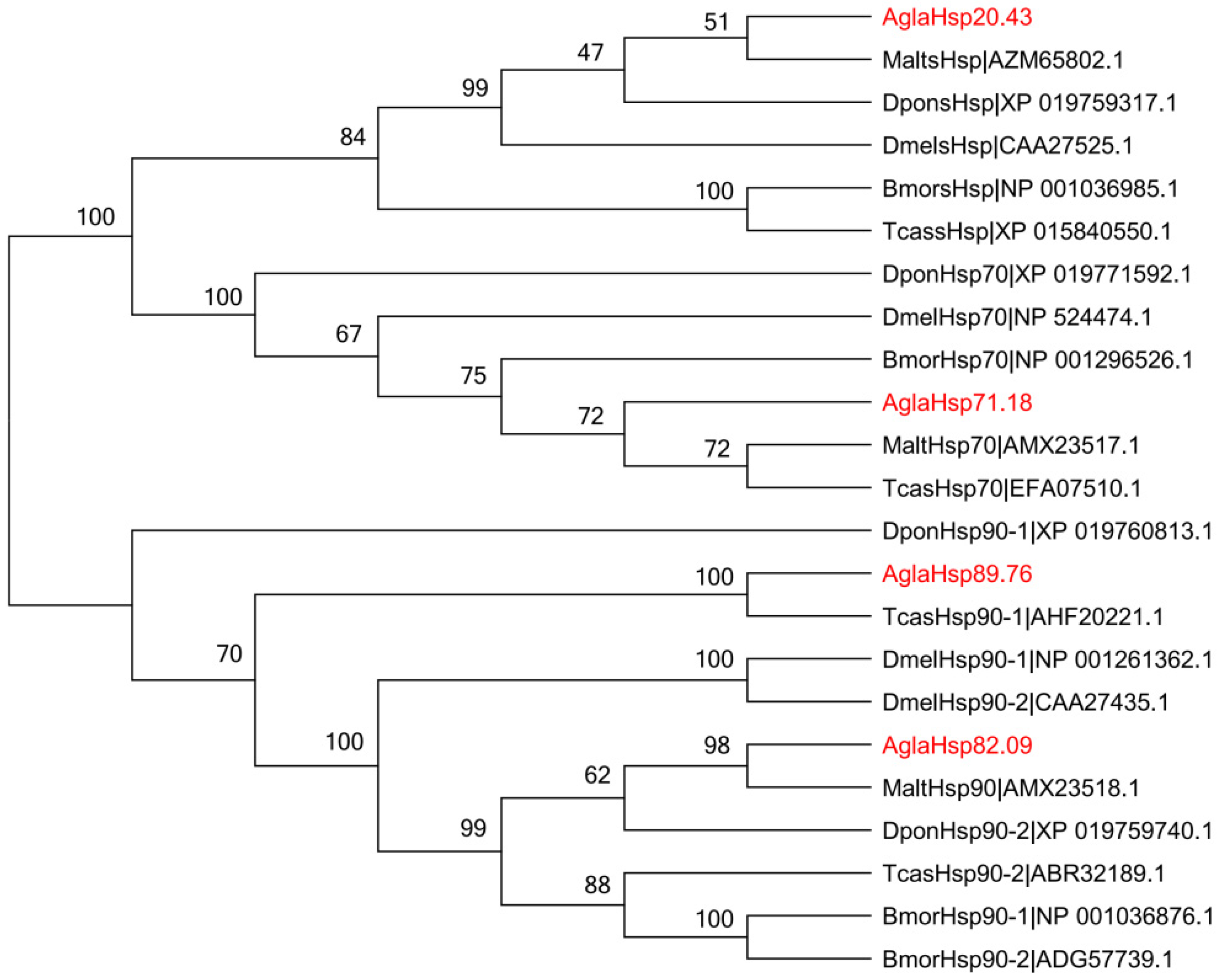
2.2. Multiple Sequence Alignment and Phylogenetic Analysis of AglaHsps
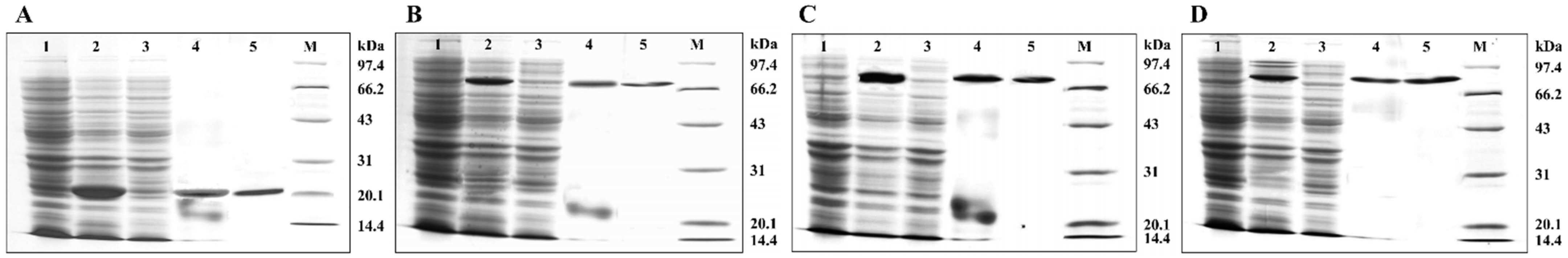
2.3. Prokaryotic Expression and Purification of AglaHsps
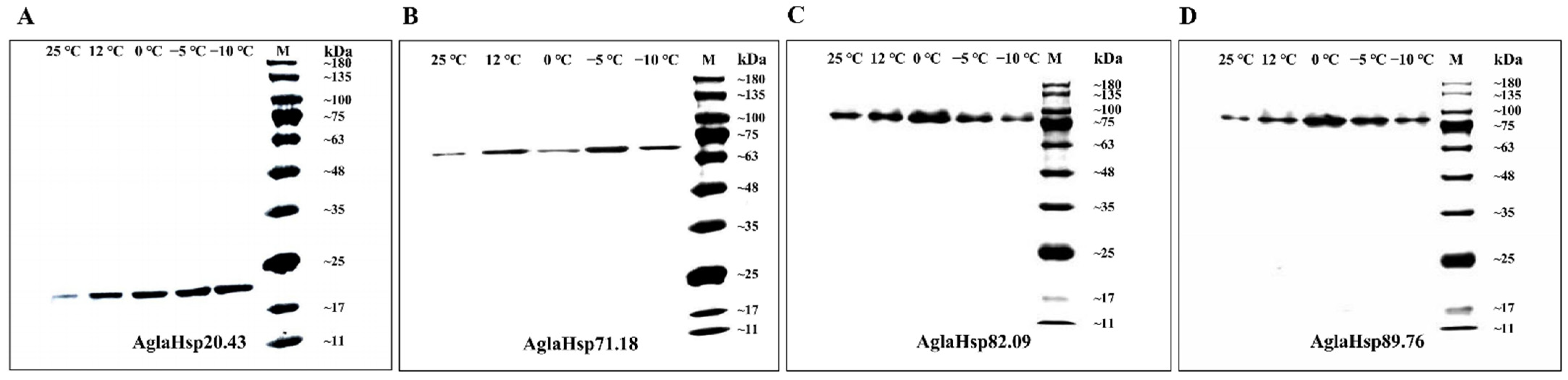
2.4. Antibody Titer Determination and Western Blotting Analysis
3. Discussion
3.1. The Diversified Structures of AglaHsps Underlie Their Functional Specificities
3.2. Evolutionary Trends and Environmental Adaptabilities of AglaHsps
3.3. AglaHsps Function in the Regulatory Networks
4. Materials and Methods
4.1. Insect Collection and Temperature Treatments
4.2. RNA Extraction and cDNA Library Construction
4.3. Cloning of AglaHsp Genes
4.4. Bioinformatic Analysis
4.5. Prokaryotic Expression and Protein Purification
4.6. Preparation of AglaHsps Antibody
4.7. Western Blotting Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [Green Version]
- Lindquist, S.; Craig, E.A. The heat-shock proteins. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef]
- Parsell, D.A.; Lindquist, S. The function of heat-shock proteins in stress tolerance: Degradation and reactivation of damaged proteins. Annu. Rev. Genet. 1993, 27, 437–496. [Google Scholar] [CrossRef]
- Lindquist, S. The heat-shock response. Annu. Rev. Biochem. 1986, 55, 1151–1191. [Google Scholar] [CrossRef] [PubMed]
- Evgen’ev, M.B.; Garbuz, D.G.; Zatsepina, O.G. Heat Shock Proteins and Whole Body Adaptation to Extreme Environments; Springer: Dordrecht, The Netherlands, 2014. [Google Scholar]
- Chen, B.; Feder, M.E.; Kang, L. Evolution of heat-shock protein expression underlying adaptive responses to environmental stress. Mol. Ecol. 2018, 27, 3040–3054. [Google Scholar] [CrossRef] [Green Version]
- King, A.M.; MacRae, T.H. Insect heat shock proteins during stress and diapause. Annu. Rev. Entomol. 2015, 60, 59–75. [Google Scholar] [CrossRef]
- Haslbeck, M.; Franzmann, T.; Weinfurtner, D.; Buchner, J. Some like it hot: The structure and function of small heat-shock proteins. Nat. Struct. Mol. Biol. 2005, 12, 842–846. [Google Scholar] [CrossRef] [PubMed]
- Basha, E.; O’Neill, H.; Vierling, E. Small heat shock proteins and α-crystallins: Dynamic proteins with flexible functions. Trends Biochem. Sci. 2012, 37, 106–117. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M.P. Gymnastics of molecular chaperones. Mol. Cell 2010, 39, 321–331. [Google Scholar] [CrossRef] [Green Version]
- Zhuravleva, A.; Gierasch, L.M. Allosteric signal transmission in the nucleotide-binding domain of 70-kDa heat shock protein (Hsp70) molecular chaperones. Proc. Natl. Acad. Sci. USA 2011, 108, 6987–6992. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.P. Hsp70 chaperone dynamics and molecular mechanism. Trends Biochem. Sci. 2013, 38, 507–514. [Google Scholar] [CrossRef]
- Richter, K.; Haslbeck, M.; Buchner, J. The heat shock response: Life on the verge of death. Mol. Cell 2010, 40, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Kityk, R.; Kopp, J.; Sinning, I.; Mayer, M.P. Structure and dynamics of the ATP-bound open conformation of Hsp70 chaperones. Mol. Cell 2012, 48, 863–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clare, D.K.; Saibil, H.R. ATP-driven molecular chaperone machines. Biopolymers 2013, 99, 846–859. [Google Scholar] [CrossRef] [PubMed]
- Wegele, H.; Müller, L.; Buchner, J. Hsp70 and Hsp90—A relay team for protein folding. Rev. Physiol. Biochem. Pharmacol. 2004, 151, 1–44. [Google Scholar] [PubMed]
- Bai, J.; Wang, Y.C.; Liu, Y.C.; Chang, Y.W.; Liu, X.N.; Gong, W.R.; Du, Y.Z. Isolation of two new genes encoding heat shock protein 70 in Bemisia tabaci and analysis during thermal stress. Int. J. Biol. Macromol. 2021, 193, 933–940. [Google Scholar] [CrossRef]
- Li, H.; Qiao, H.; Liu, Y.J.; Li, S.Y.; Tan, J.J.; Hao, D.J. Characterization, expression profiling, and thermal tolerance analysis of heat shock protein 70 in pine sawyer beetle, Monochamus alternatus hope (Coleoptera: Cerambycidae). Bull. Entomol. Res. 2021, 111, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.L.; Zhu, F.; Lu, M.X.; Du, Y.Z. Characterization and functional analysis of Cshsp19.0 encoding a small heat shock protein in Chilo suppressalis (Walker). Int. J. Biol. Macromol. 2021, 188, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.W.; Zhang, X.X.; Lu, M.X.; Du, Y.Z.; Zhu-Salzman, K. Molecular cloning and characterization of small heat shock protein genes in the invasive leaf miner fly, Liriomyza trifolii. Genes 2019, 10, 775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haack, R.A.; Hérard, F.; Sun, J.H.; Turgeon, J.J. Managing invasive populations of Asian longhorned beetle and citrus longhorned beetle: A worldwide perspective. Annu. Rev. Entomol. 2010, 55, 521–546. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.T.; Bancroft, J.; Li, G.H.; Gao, R.T.; Teale, S. Dispersal of Anoplophora glabripennis (Cerambycidae). Environ. Entomol. 2001, 30, 1036–1040. [Google Scholar] [CrossRef] [Green Version]
- Javal, M.; Roux, G.; Roques, A.; Sauvard, D. Asian Long-horned Beetle dispersal potential estimated in computer-linked flight mills. J. Appl. Entomol. 2018, 142, 282–286. [Google Scholar] [CrossRef]
- Javal, M.; Roques, A.; Haran, J.; Hérard, F.; Keena, M.; Roux, G. Complex invasion history of the Asian long-horned beetle: Fifteen years after first detection in Europe. J. Pest Sci. 2019, 92, 173–187. [Google Scholar] [CrossRef]
- Hérard, F.; Ciampitti, M.; Maspero, M.; Krehan, H.; Benker, U.; Boegel, C.; Schrage, R.; Bouhot-Delduc, L.; Bialooki, P. Anoplophora species in Europe: Infestations and management processes. EPPO Bull. 2006, 36, 470–474. [Google Scholar] [CrossRef]
- Javal, M.; Lombaert, E.; Tsykun, T.; Courtin, C.; Kerdelhué, C.; Prospero, S.; Roques, A.; Roux, G. Deciphering the worldwide invasion of the Asian long-horned beetle: A recurrent invasion process from the native area together with a bridgehead effect. Mol. Ecol. 2019, 28, 951–967. [Google Scholar] [CrossRef]
- Asian Longhorned Beetle Response Guidelines. 2014. Available online: https://www.aphis.usda.gov/plant_health/plant_pest_info/asian_lhb/downloads/response-guidelines.pdf (accessed on 15 June 2022).
- EPPO A1 and A2 Lists of Pests Recommended for Regulation as Quarantine Pests. Available online: https://gd.eppo.int/taxon/ANOLGL/documents (accessed on 15 June 2022).
- Feng, Y.Q.; Xu, L.L.; Li, W.B.; Xu, Z.C.; Cao, M.; Wang, J.L.; Tao, J.; Zong, S.X. Seasonal changes in supercooling capacity and major cryoprotectants of overwintering Asian longhorned beetle (Anoplophora glabripennis) larvae. Agric. For. Entomol. 2016, 18, 302–312. [Google Scholar] [CrossRef]
- Huang, J.X.; Qu, B.R.; Fang, G.F.; Li, X.D.; Zong, S.X. The drivers of the Asian longhorned beetle disaster show significant spatial heterogeneity. Ecol. Indic. 2020, 117, 106680. [Google Scholar] [CrossRef]
- Wang, Z.G. Study on the Occurrence Dynamics of Anoplophora glabripennis (Coleoptera: Cerambycidae) and Its Control Measures. Ph.D. Thesis, Northeast Forestry University, Harbin, China, 2004. [Google Scholar]
- Feng, Y.Q. Cold Hardiness and Adaptive Mechanism of the Anoplophora glabripennis Larva. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2017. [Google Scholar]
- Xu, Y.B.; Li, Y.R.; Wang, Q.Q.; Zheng, C.C.; Zhao, D.F.; Shi, F.M.; Liu, X.H.; Tao, J.; Zong, S.X. Identification of key genes associated with overwintering in Anoplophora glabripennis larva using gene co-expression network analysis. Pest Manag. Sci. 2021, 77, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.B.; Shi, F.M.; Li, Y.R.; Tao, J.; Zong, S.X. Genome-wide identification and expression analysis of the Hsp gene superfamily in Asian long-horned beetle (Anoplophora glabripennis). Int. J. Biol. Macromol. 2022, 200, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Toxopeus, J.; Sinclair, B.J. Mechanisms underlying insect freeze tolerance. Biol. Rev. 2018, 93, 1891–1914. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.H.; Wang, C.Z.; Kang, L. Cloning and expression of five heat shock protein genes in relation to cold hardening and development in the leafminer, Liriomyza sativa. J. Insect Physiol. 2009, 55, 279–285. [Google Scholar] [CrossRef]
- Zhang, G.J.; Storey, J.M.; Storey, K.B. Chaperone proteins and winter survival by a freeze tolerant insect. J. Insect Physiol. 2011, 57, 1115–1122. [Google Scholar] [CrossRef]
- Sun, Y.; MacRae, T.H. Small heat shock proteins: Molecular structure and chaperone function. Cell. Mol. Life Sci. 2005, 62, 2460–2476. [Google Scholar] [CrossRef] [PubMed]
- Mchaourab, H.S.; Godar, J.A.; Stewart, P.L. Structure and mechanism of protein stability sensors: Chaperone activity of small heat shock proteins. Biochemistry 2009, 48, 3828–3837. [Google Scholar] [CrossRef] [Green Version]
- Nakamoto, H.; Vígh, L. The small heat shock proteins and their clients. Cell. Mol. Life Sci. 2007, 64, 294–306. [Google Scholar] [CrossRef]
- Waters, E.R. The evolution, function, structure, and expression of the plant sHSPs. J. Exp. Bot. 2013, 64, 391–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, E.R.; Vierling, E. Plant small heat shock proteins—Evolutionary and functional diversity. New Phytol. 2020, 227, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Haley, D.A.; Bova, M.P.; Huang, Q.L.; Mchaourab, H.S.; Stewart, P.L. Small heat-shock protein structures reveal a continuum from symmetric to variable assemblies. J. Mol. Biol. 2000, 298, 261–272. [Google Scholar] [CrossRef]
- Bar-Lavan, Y.; Shemesh, N.; Ben-Zvi, A. Chaperone families and interactions in metazoa. Essays Biochem. 2016, 60, 237–253. [Google Scholar]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Rev. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Vogel, M.; Mayer, M.P.; Bukau, B. Allosteric regulation of hsp70 chaperones involves a conserved interdomain linker. J. Biol. Chem. 2006, 281, 38705–38711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.E.; Hipp, M.S.; Bracher, A.; Hayer-Hartl, M.; Ulrich Hartl, F. Molecular chaperone functions in protein folding and proteostasis. Annu. Rev. Biochem. 2013, 82, 323–355. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S. Phylogenetic analysis of the 90 kD heat shock family of protein sequences and an examination of the relationship among animals, plants, and fungi species. Mol. Biol. Evol. 1995, 12, 1063–1073. [Google Scholar] [PubMed] [Green Version]
- Csermely, P.; Schnaider, T.; Soti, C.; Prohászka, Z.; Nardai, G. The 90-kDa Molecular Chaperone Family: Structure, Function, and Clinical Applications. A Comprehensive Review. Pharmacol. Ther. 1998, 79, 129–168. [Google Scholar] [CrossRef]
- Chen, B.; Zhong, D.B.; Monteiro, A. Comparative genomics and evolution of the HSP90 family of genes across all kingdoms of organisms. BMC Genomics 2006, 7, 156. [Google Scholar] [CrossRef] [Green Version]
- Kriehuber, T.; Rattei, T.; Weinmaier, T.; Bepperling, A.; Haslbeck, M.; Buchner, J. Independent evolution of the core domain and its flanking sequences in small heat shock proteins. FASEB J. 2010, 24, 3633–3642. [Google Scholar] [CrossRef]
- Thomas, J.H. Rapid Birth-Death Evolution Specific to Xenobiotic Cytochrome P450 Genes in Vertebrates. PLoS Genet. 2007, 3, e67. [Google Scholar] [CrossRef] [Green Version]
- Daugaard, M.; Rohde, M.; Jäättelä, M. The heat shock protein 70 family: Highly homologous proteins with overlapping and distinct functions. FEBS Lett. 2007, 581, 3702–3710. [Google Scholar] [CrossRef] [Green Version]
- Pelham, H.R.B. Hsp70 accelerates the recovery of nucleolar morphology after heat shock. EMBO J. 1984, 3, 3095–3100. [Google Scholar] [CrossRef]
- Chen, Z.C.; Wu, W.S.; Lin, M.T.; Hsu, C.C. Protective effect of transgenic expression of porcine heat shock protein 70 on hypothalamic ischemic and oxidative damage in a mouse model of heatstroke. BMC Neurosci. 2009, 10, 111. [Google Scholar] [CrossRef] [PubMed]
- Taipale, M.; Jarosz, D.F.; Lindquist, S. HSP90 at the hub of protein homeostasis: Emerging mechanistic insights. Nat. Rev. Mol. Cell Biol. 2010, 11, 515–528. [Google Scholar] [CrossRef]
- Krishna, P.; Gloor, G. The Hsp90 family of proteins in Arabidopsis thaliana. Cell Stress Chaperones 2001, 6, 238–246. [Google Scholar] [CrossRef]
- Guillen, A.C.; Borges, M.E.; Herrerias, T.; Kandalski, P.K.; de Arruda Marins, E.; Viana, D.; de Souza, M.R.D.P.; Oliveira do Carmo Daloski, L.; Donatti, L. Effect of gradual temperature increase on the carbohydrate energy metabolism responses of the Antarctic fish Notothenia rossii. Mar. Environ. Res. 2019, 150, 104779. [Google Scholar] [CrossRef]
- Pfister, T.D.; Storey, K.B. Insect freeze tolerance: Roles of protein phosphatases and protein kinase A. Insect Biochem. Mol. Biol. 2006, 36, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, M.; Pollex, T.; Hanna, K.; Tuorto, F.; Meusburger, M.; Helm, M.; Lyko, F. RNA methylation by Dnmt2 protects transfer RNAs against stress-induced cleavage. Genes Dev. 2010, 24, 1590–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Chang, G.F.; Li, Z.Z.; Abouzaid, M.; Du, X.Y.; Hull, J.J.; Ma, W.H.; Lin, Y.J. The HSP/co-chaperone network in environmental cold adaptation of Chilo suppressalis. Int. J. Biol. Macromol. 2021, 187, 780–788. [Google Scholar] [CrossRef]
- Kang, Z.W.; Liu, F.H.; Liu, X.; Yu, W.B.; Tan, X.L.; Zhang, S.Z.; Tian, H.G.; Liu, T.X. The Potential Coordination of the Heat-Shock Proteins and Antioxidant Enzyme Genes of Aphidius gifuensis in Response to Thermal Stress. Front. Physiol. 2017, 8, 976. [Google Scholar] [CrossRef] [Green Version]
- Buckley, B.A.; Hofmann, G.E. Magnitude and duration of thermal stress determine kinetics of hsp gene regulation in the goby Gillichthys mirabilis. Physiol. Biochem. Zool. 2004, 77, 570–581. [Google Scholar] [CrossRef] [Green Version]
- Krebs, R.A.; Feder, M.E. Deleterious consequences of Hsp70 overexpression in Drosophila melanogaster larvae. Cell Stress Chaperones 1997, 2, 60–71. [Google Scholar] [CrossRef]
- Feder, J.H.; Rossi, J.M.; Solomon, J.; Solomon, N.; Lindquist, S. The consequences of expressing hsp70 in Drosophila cells at normal temperatures. Genes. Dev. 1992, 6, 1402–1413. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Huang, J.F. Artificial feeding of Anoplophora glabripennis. J. Beijing For. Univ. 1992, 14, 61–67. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AglaHsp20.43 | AglaHsp71.18 | AglaHsp82.09 | AglaHsp89.76 | |
|---|---|---|---|---|
| Coding sequence | 546 bp | 1950 bp | 2154 bp | 2355 bp |
| Amino acid residues | 181 aa | 649 aa | 717 aa | 784 aa |
| Molecular weight (kDa) | 20.43 | 71.18 | 82.09 | 89.76 |
| Theoretical isoelectric point | 6.50 | 5.52 | 5.00 | 4.88 |
| Instability index | 44.10 | 37.03 | 38.39 | 40.06 |
| Hydrophilic protein | −0.631 | −0.427 | −0.665 | −0.600 |
| Transmembrane structures | No | No | No | No |
| Signal peptide | No | No | No | aa 1-20 |
| Subcellular localization | Cytoplasmic | Cytoplasmic | Cytoplasmic | Endoplasmic reticulum |
| Alpha Helix | Extended Strand | Beta Turn | Random Coil | |
|---|---|---|---|---|
| AglaHsp20.43 | 24.86% | 16.02% | 5.52% | 53.59% |
| AglaHsp71.18 | 41.29% | 18.80% | 7.40% | 32.51% |
| AglaHsp82.09 | 48.81% | 15.76% | 5.86% | 29.57% |
| AglaHsp89.76 | 51.02% | 13.78% | 3.70% | 31.51% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Li, Y.; Shi, F.; Zhang, S.; Zong, S.; Tao, J. Cloning and Molecular Characterization of Hsp Genes from Anoplophora glabripennis and Their Responses to Cold Stress. Int. J. Mol. Sci. 2022, 23, 11958. https://doi.org/10.3390/ijms231911958
Xu Y, Li Y, Shi F, Zhang S, Zong S, Tao J. Cloning and Molecular Characterization of Hsp Genes from Anoplophora glabripennis and Their Responses to Cold Stress. International Journal of Molecular Sciences. 2022; 23(19):11958. https://doi.org/10.3390/ijms231911958
Chicago/Turabian StyleXu, Yabei, Yurong Li, Fengming Shi, Sainan Zhang, Shixiang Zong, and Jing Tao. 2022. "Cloning and Molecular Characterization of Hsp Genes from Anoplophora glabripennis and Their Responses to Cold Stress" International Journal of Molecular Sciences 23, no. 19: 11958. https://doi.org/10.3390/ijms231911958