

Mapping QTLs for Reproductive Stage Salinity Tolerance in Rice Using a Cross between Hasawi and BRRI dhan28

Abstract
:1. Introduction
2. Results
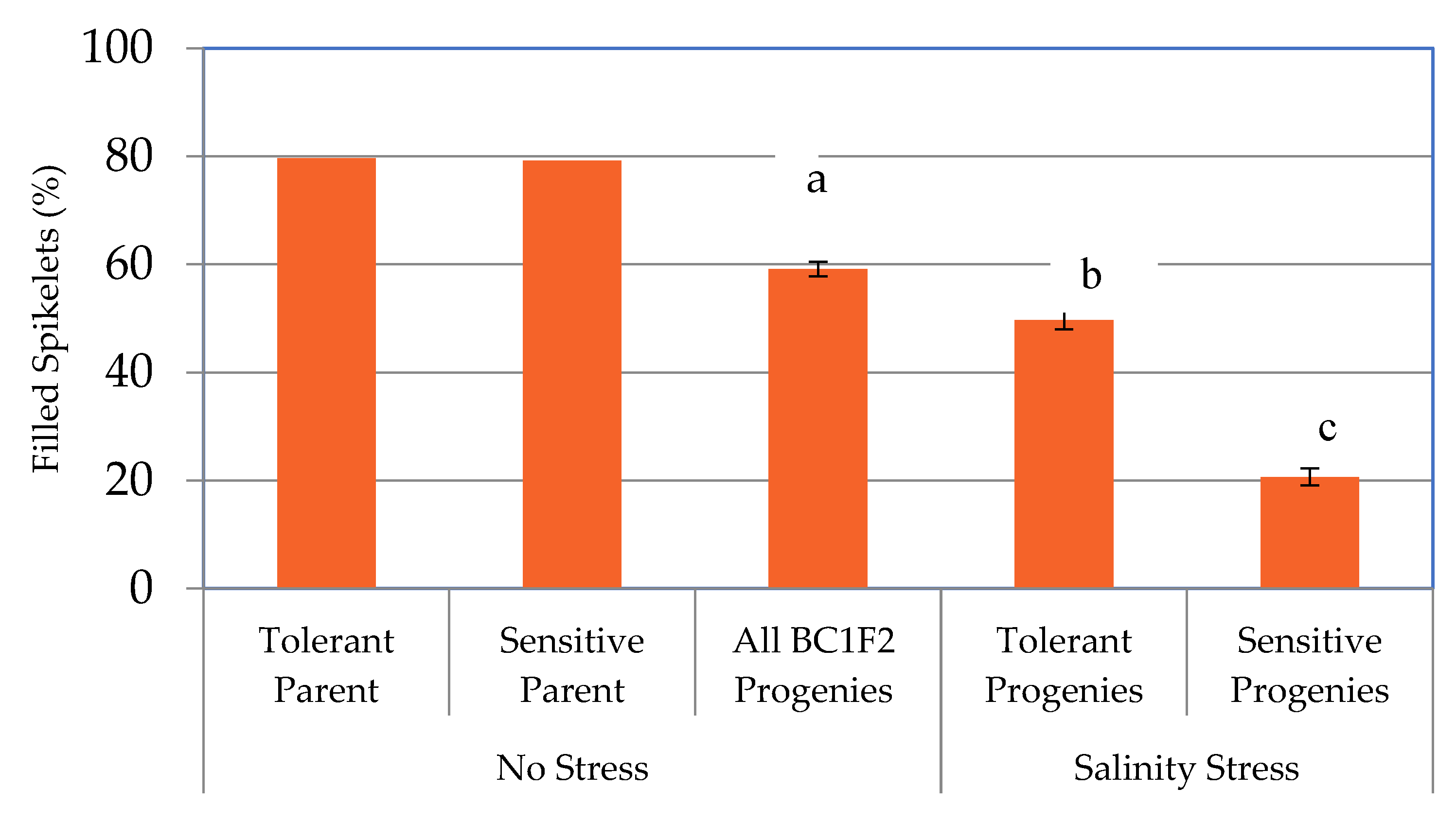
2.1. Responses of the Parental Lines and BC1F2 Progenies to Salt Stress
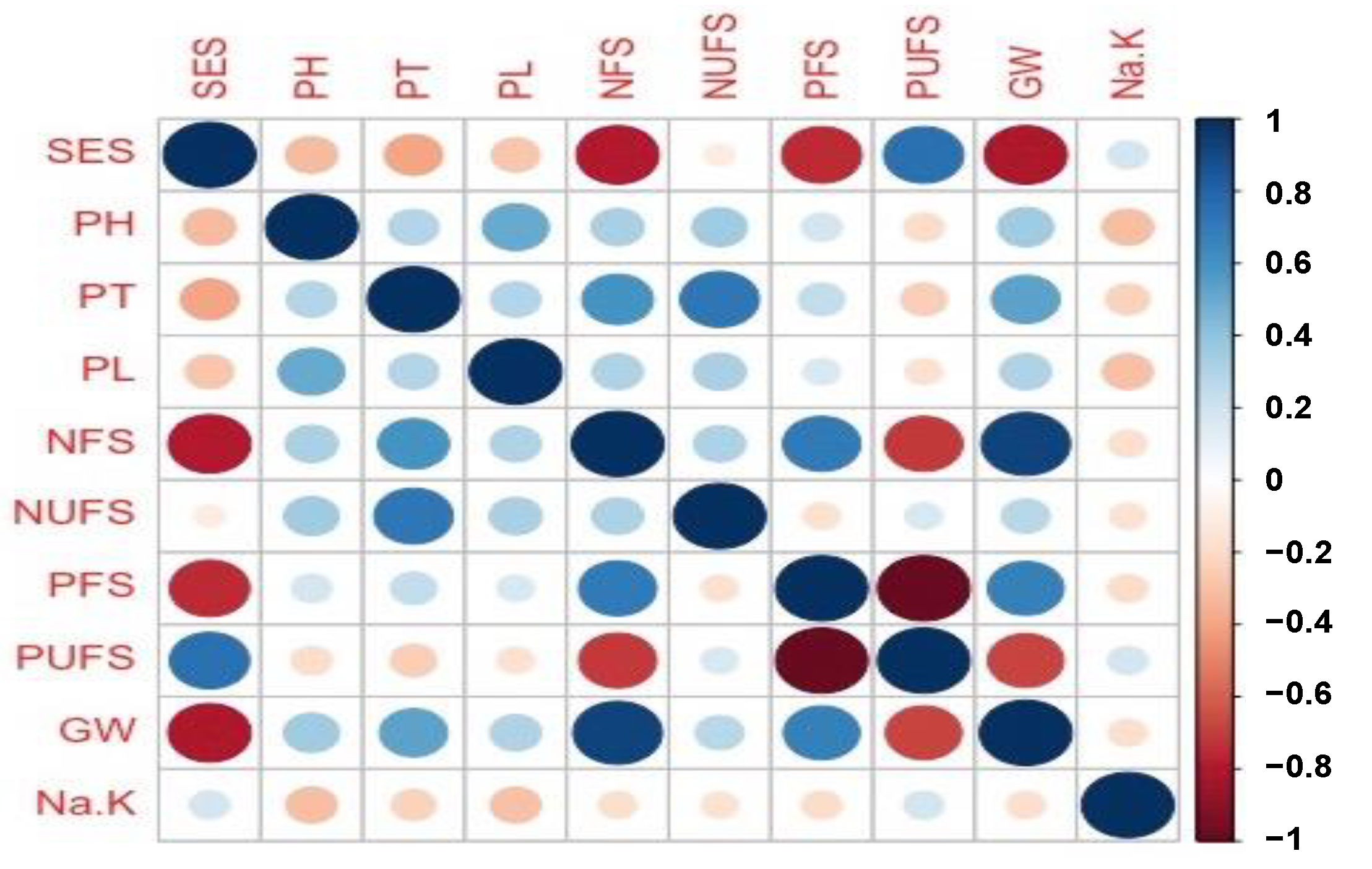
2.2. Correlation Analysis of Yield and Agronomic Components
2.3. Trait Distributions
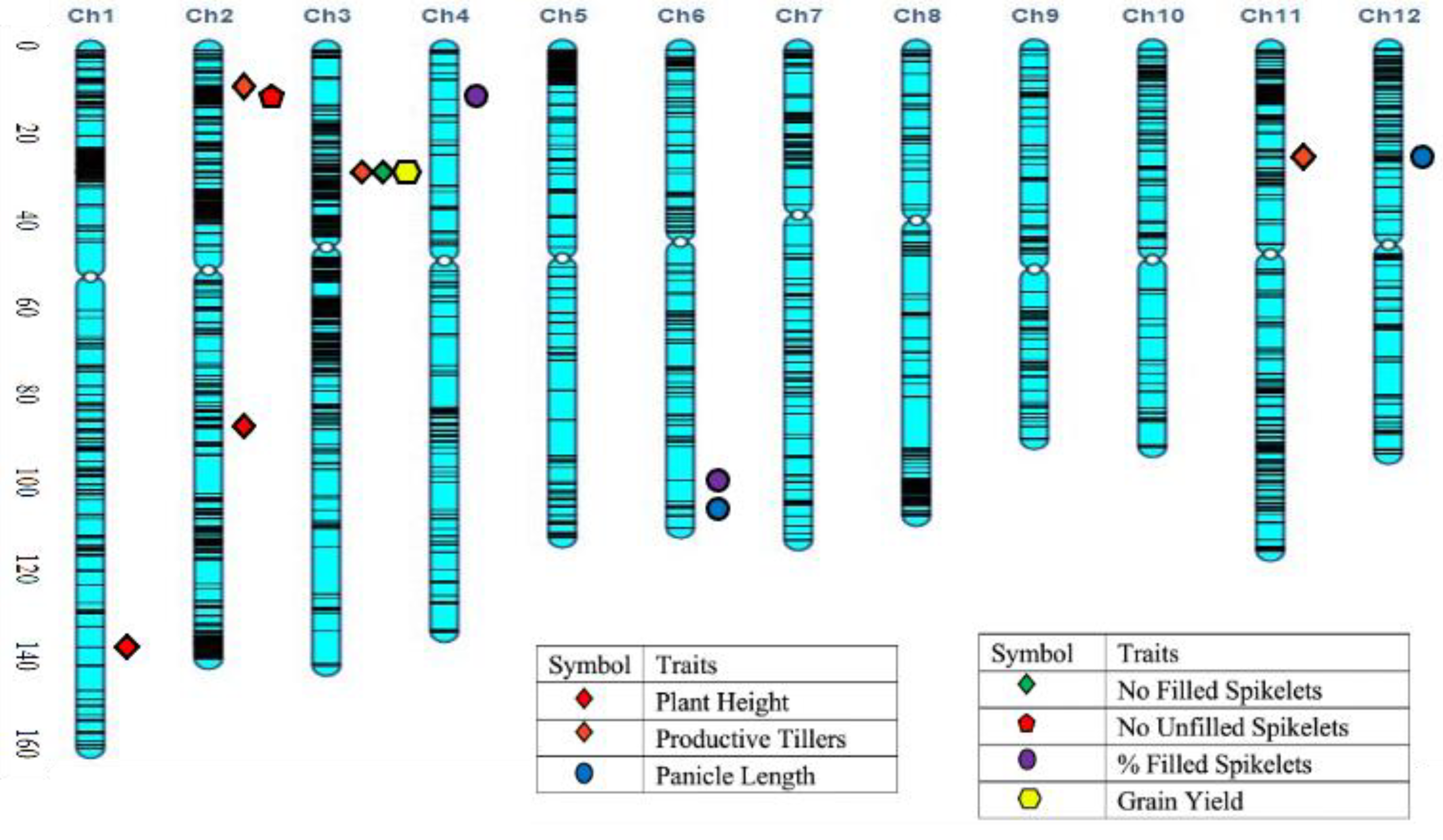
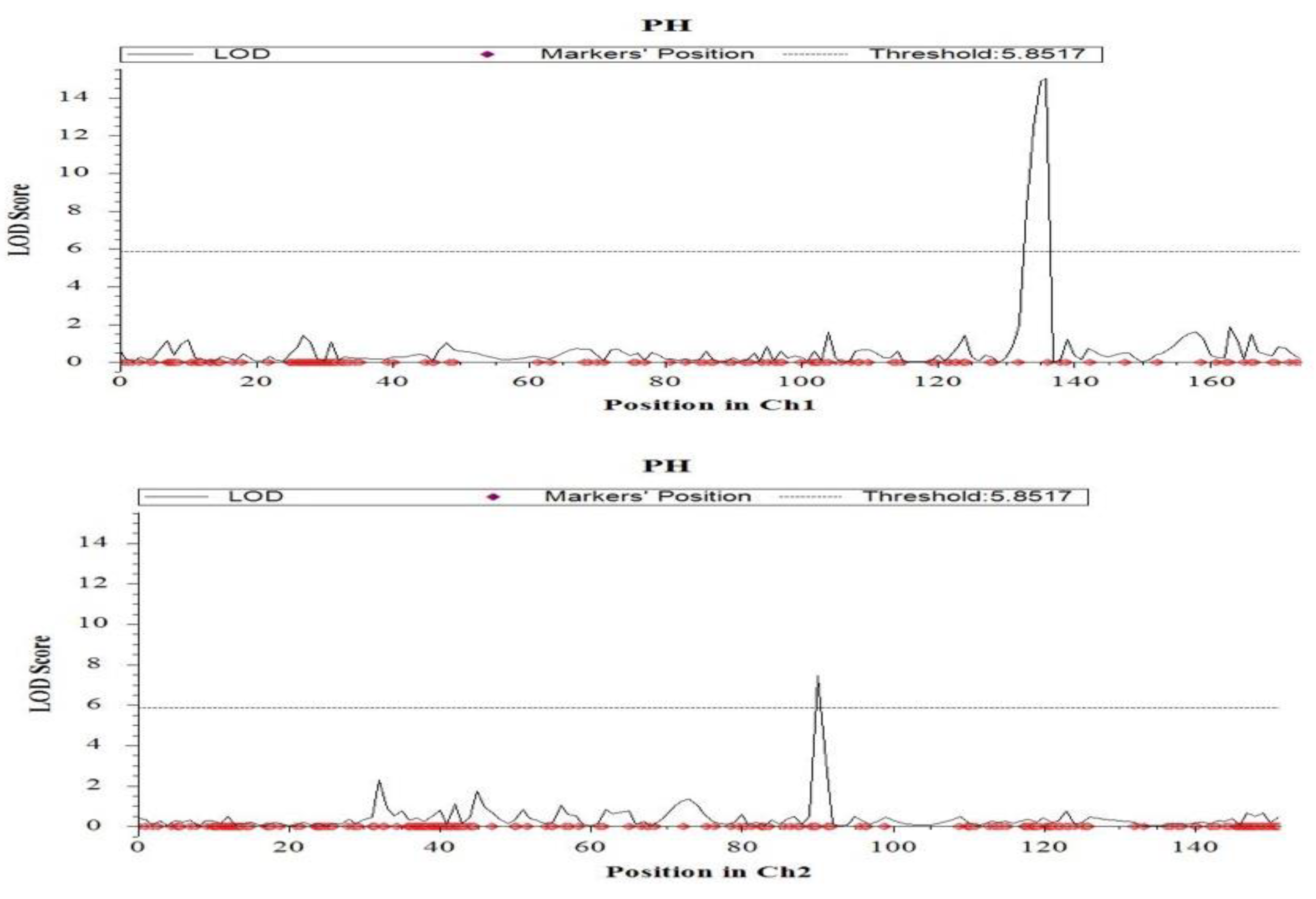
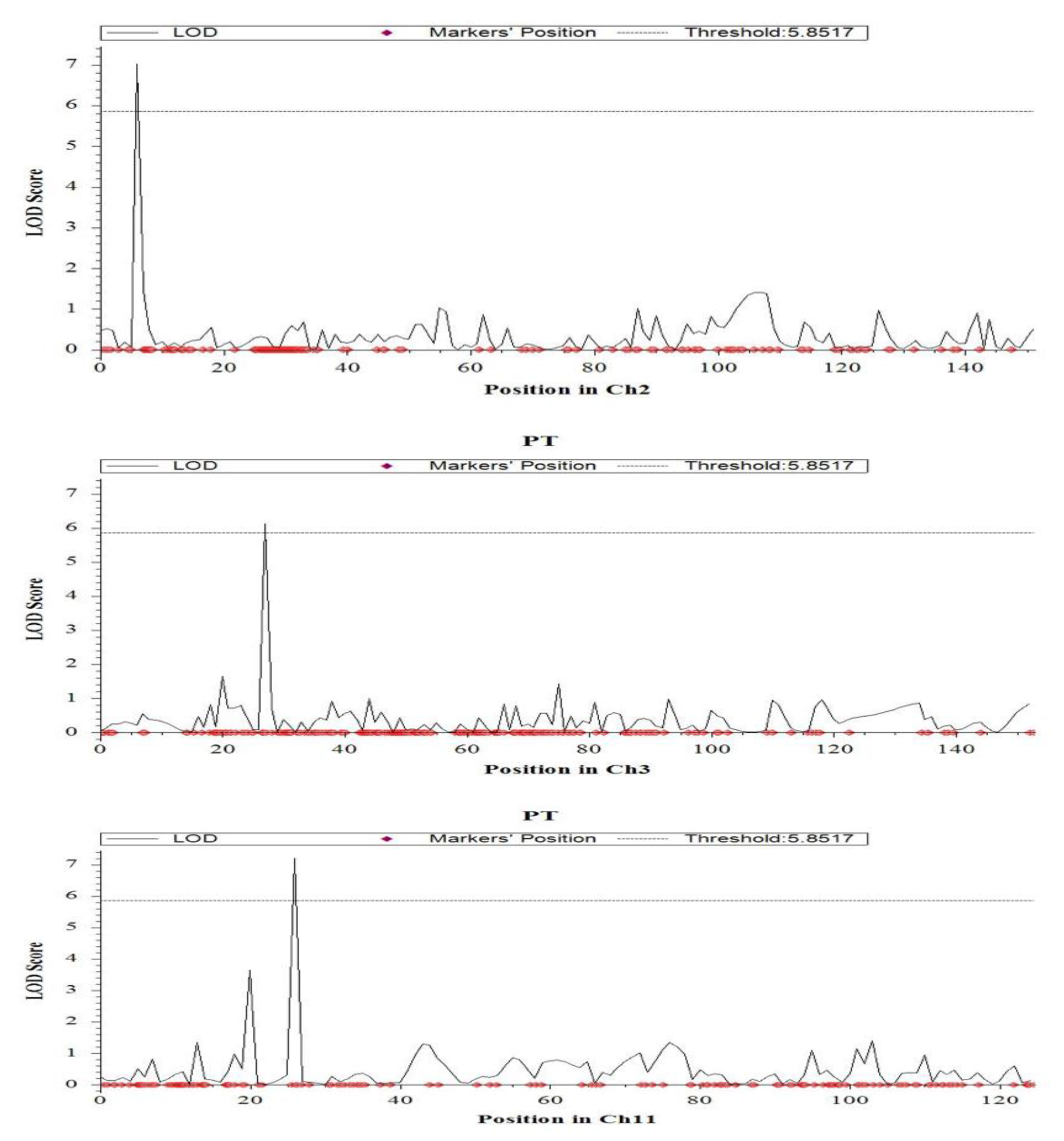
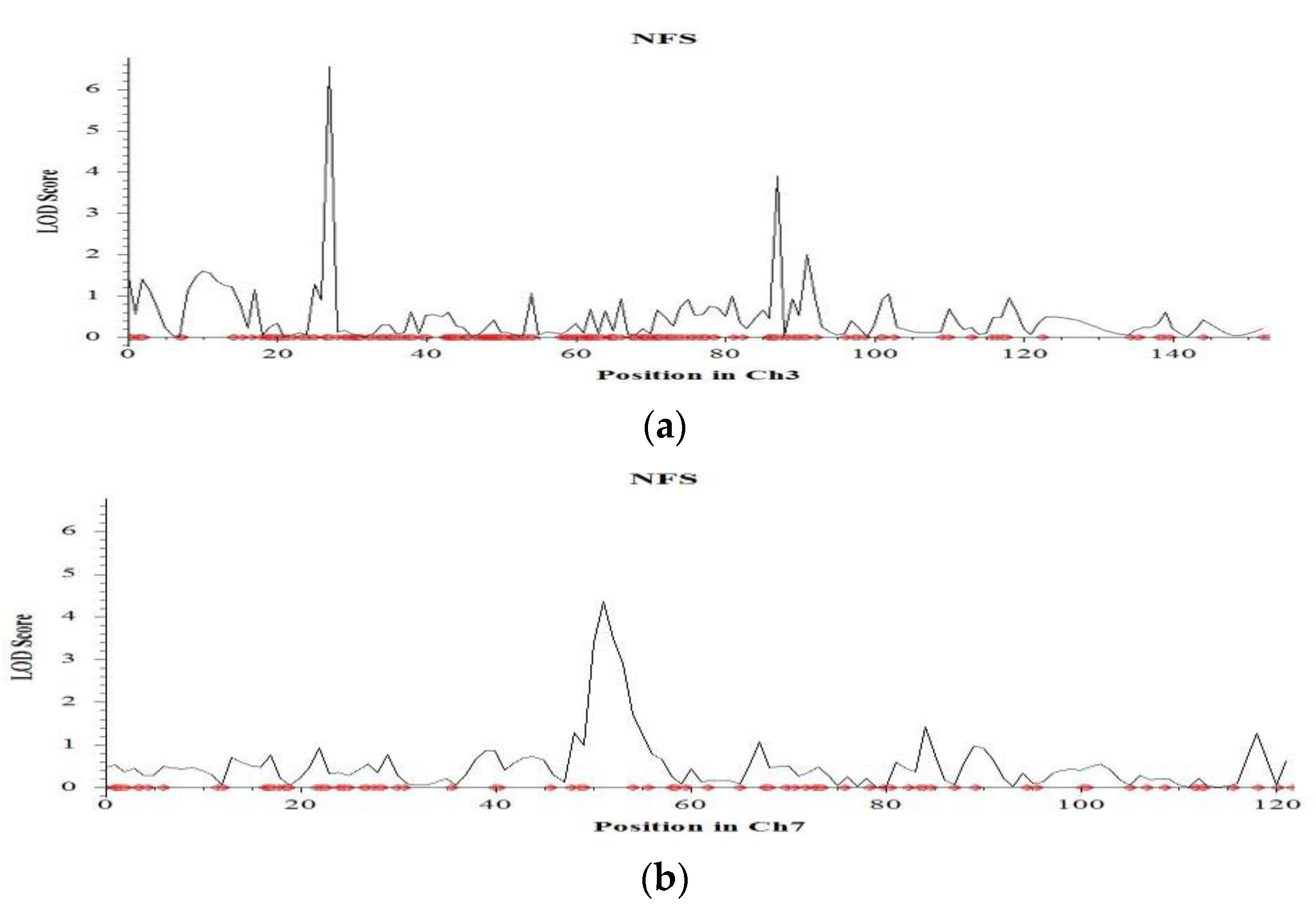
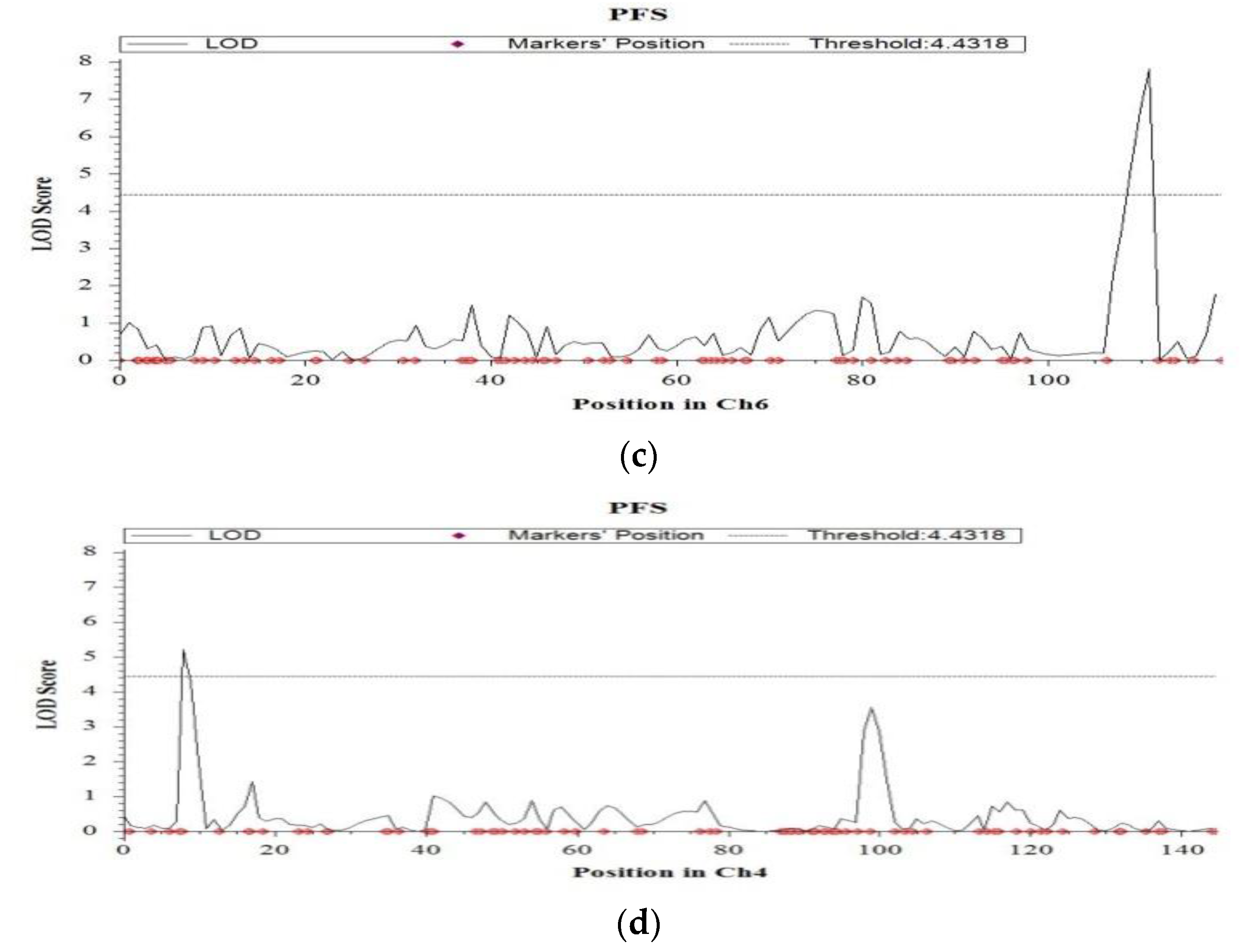
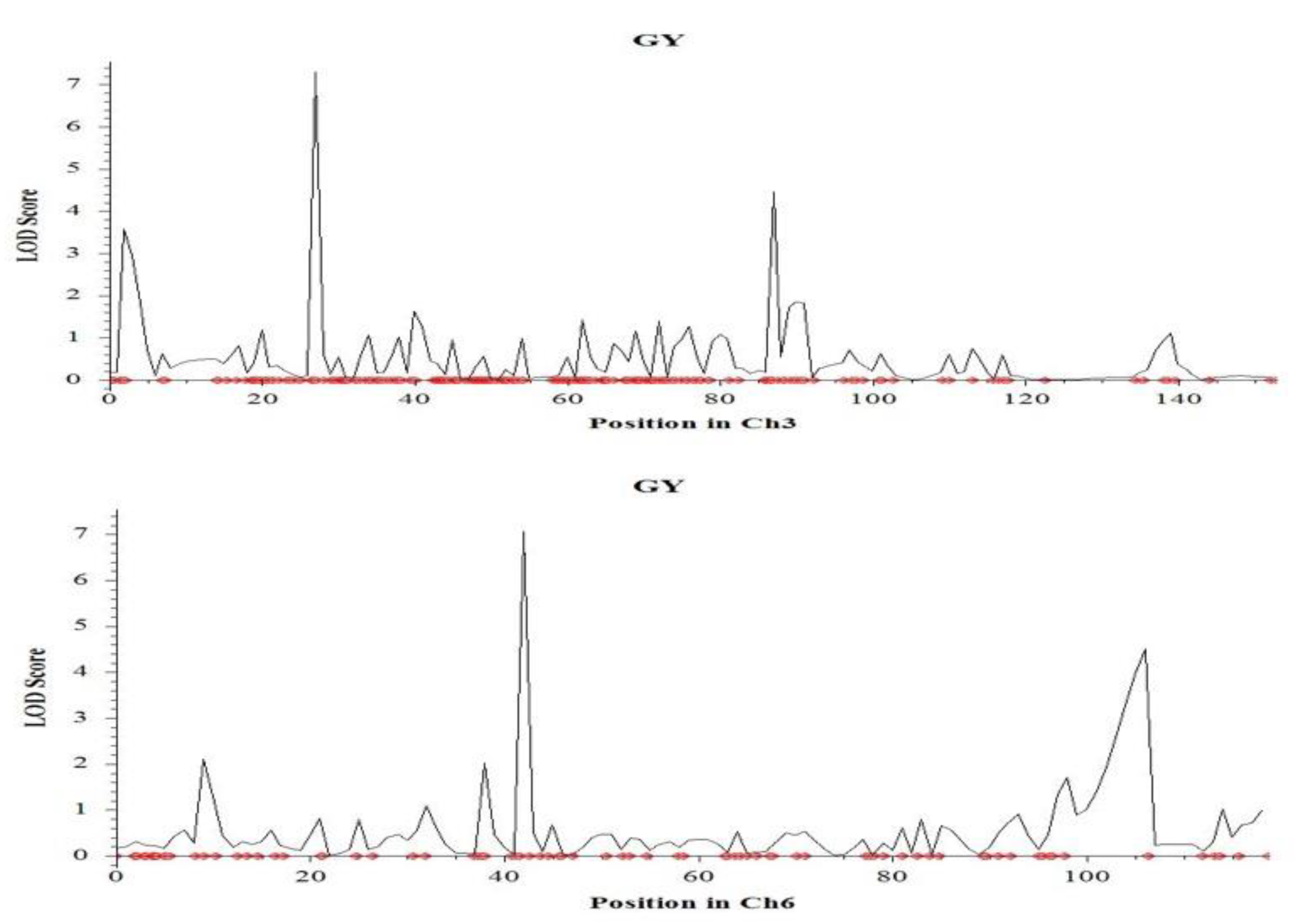
2.4. QTL Mapping for Salinity Tolerance at Reproductive Stage
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Experimental Design and Set-Up
4.3. Phenotypic Evaluation
4.3.1. Seedling Preparation and Management
4.3.2. Salinity Tolerance Assessment at Reproductive Stage
- Na or K = Concentration of sodium and potassium ion (mmol/g dwt).
- C = Concentration of sample aliquot based on atomic absorption spectrophotometer reading as determined relative to standard curve.
- d = dilution factor.
- V = extraction volume (mL).
- dwt = oven dry weight of the plant leaf (g).
4.4. Salinity Scoring and Agronomic Parameters
4.5. Genotyping of Parents and BC1F2 Progenies
4.5.1. DNA Extraction and Molecular Characterization
4.5.2. Linkage Map Construction
4.5.3. QTL Mapping for Salinity Tolerance
4.6. Candidate Gene Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flowers, T.J. Improving crop salt tolerance. J. Exp. Bot. 2004, 55, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Asif, M.A.; Schilling, R.K.; Tilbrook, J.; Brien, C.; Dowling, K.; Rabie, H.; Short, L.; Trittermann, C.; Garcia, A.; Barrett-Lennard, E.G.; et al. Mapping of novel salt tolerance QTL in an Excalibur × Kukri doubled haploid wheat population. Theor. Appl. Genet. 2018, 131, 2179–2196. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.R.; Bagali, P.G.; Hittalmani, S.; Shashidhar, H.E. Molecular mapping of quantitative trait loci associated with seedling tolerance to salt stress in rice (Oryza sativa L.). Curr. Sci. 2000, 78, 162–164. [Google Scholar]
- IPCC. Special Report on Climate Change and Land; IPCC: Geneva, Switzerland, 2019. [Google Scholar] [CrossRef]
- Ismail, A.M.; Horie, T. Genomics, Physiology, and Molecular Breeding Approaches for Improving Salt Tolerance. Annu. Rev. Plant Biol. 2017, 68, 405–434. [Google Scholar] [CrossRef]
- Singh, R.K.; Gregorio, G.B.; Jain, R.K. QTL mapping for salinity tolerance in rice. Physiol. Mol. Biol. Plants 2007, 13, 87. [Google Scholar]
- Singh, R.K.; Flowers, T.J. The physiology and molecular biology of the effects of salinity on rice. In Handbook of Plant and Crop Stress, 3rd ed.; Pessarakli, M., Ed.; Taylor and Francis: Boca Raton, FL, USA, 2010; pp. 901–942. [Google Scholar]
- Ahmadizadeh, M.; Vispo, N.A.; Calapit-Palao, C.D.O.; Pangaan, I.D.; Viña, C.D.; Singh, R.K. Reproductive stage salinity tolerance in rice: A complex trait to phenotype. Indian J. Plant Physiol. 2016, 21, 528–536. [Google Scholar] [CrossRef]
- Singh, R.K.; Kota, S.; Flowers, T.J. Salt tolerance in rice: Seedling and reproductive stage QTL mapping come of age. Theor. Appl. Genet. 2021, 134, 3495–3533. [Google Scholar] [CrossRef] [PubMed]
- Gimhani, D.; Gregorio, G.B.; Kottearachchi, N.S.; Samarasinghe, W.L.G. SNP-based discovery of salinity-tolerant QTLs in a bi-parental population of rice (Oryza sativa). Mol. Genet. Genom. 2016, 291, 2081–2099. [Google Scholar] [CrossRef]
- Negrão, S.; Schmöckel, S.M.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2016, 119, mcw191. [Google Scholar] [CrossRef]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef]
- Tuberosa, R.; Salvi, S. QTLs and Genes for Tolerance to Abiotic Stress in Cereals. In Cereal Genomics; Springer: Berlin/Heidelberg, Germany, 2006; pp. 253–315. [Google Scholar] [CrossRef]
- Thomson, M.J.; Ismail, A.M.; McCouch, S.R.; Mackill, D.J. Marker assisted breeding. In Abiotic Stress Adaptation in Plants: Physiological, Molecular and Genomic Foundation; Springer: Dordrecht, The Netherlands, 2010. [Google Scholar] [CrossRef]
- Thomson, M.J.; Zhao, K.; Wright, M.; McNally, K.L.; Rey, J.; Tung, C.-W.; Reynolds, A.; Scheffler, B.; Eizenga, G.; McClung, A.; et al. High-throughput single nucleotide polymorphism genotyping for breeding applications in rice using the BeadXpress platform. Mol. Breed. 2011, 29, 875–886. [Google Scholar] [CrossRef]
- Thomson, M.J. High-Throughput SNP Genotyping to Accelerate Crop Improvement. Plant Breed. Biotechnol. 2014, 2, 195–212. [Google Scholar] [CrossRef]
- Hossain, H.; Rahman, M.A.; Alam, M.S.; Singh, R.K. Mapping of Quantitative Trait Loci Associated with Reproductive-Stage Salt Tolerance in Rice. J. Agron. Crop Sci. 2015, 201, 17–31. [Google Scholar] [CrossRef]
- Fan, X.; Jiang, H.; Meng, L.; Chen, J. Gene Mapping, Cloning and Association Analysis for Salt Tolerance in Rice. Int. J. Mol. Sci. 2021, 22, 11674. [Google Scholar] [CrossRef]
- Mai, N.S.; Hanh, D.D.; Nakashima, M.; Kumamoto, K.; Thuy, N.T.T.; Kobata, T.; Saitoh, K.; Hirai, Y. Identification and Validation of QTLs for Yield and Yield Components under Long-Term Salt Stress Using IR64 CSSLs in the Genetic Background of Koshihikari and Their Backcross Progenies. Agriculture 2021, 11, 777. [Google Scholar] [CrossRef]
- Thomson, M.J.; Singh, N.; Dwiyanti, M.S.; Wang, D.R.; Wright, M.H.; Perez, F.A.; Declerck, G.; Chin, J.H.; Malitic-Layaoen, G.A.; Juanillas, V.M.; et al. Large-scale deployment of a rice 6 K SNP array for genetics and breeding applications. Rice 2017, 10, 40. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Bimpong, I.K.; Bizimana, J.B.; Pascual, E.D.; Arceta, M.; Swamy, B.P.M.; Diaw, F.; Singh, R.K. Mapping QTLs using a novel source of salinity tolerance from Hasawi and their interaction with environments in rice. Rice 2017, 10, 47. [Google Scholar] [CrossRef]
- Bizimana, J.B.; Luzi-Kihupi, A.; Murori, R.W.; Singh, R.K. Identification of quantitative trait loci for salinity tolerance in rice (Oryza sativa L.) using IR29/Hasawi mapping population. J. Genet. 2017, 96, 571–582. [Google Scholar] [CrossRef]
- Bimpong, I.K.; Manneh, B.; Diop, B.; Ghislain, K.; Sow, A.; Amoah, N.K.A.; Gregorio, G.; Singh, R.K.; Ortiz, R.; Wopereis, M. New quantitative trait loci for enhancing adaptation to salinity in rice from Hasawi, a Saudi landrace into three African cultivars at the reproductive stage. Euphytica 2014, 200, 45–60. [Google Scholar] [CrossRef]
- Mao, B.-B.; Cai, W.-J.; Zhang, Z.-H.; Hu, Z.-L.; Li, P.; Zhu, L.-H.; Zhu, Y.-G. Characterization of QTLs for harvest index and source-sink characters in a DH population of rice (Oryza sativa L.). Yi Chuan Xue Bao 2003, 30, 1118–1126. [Google Scholar]
- Sasaki, A.; Ashikari, M.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Swapan, D.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.S.; et al. A mutant gibberellin-synthesis gene in rice. Nature 2002, 416, 701–702. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Borromeo, T.H.; Diaz, M.G.Q.; Amas, J.; Rahman, M.A.; Thomson, M.; Gregorio, G.B. Dissecting QTLs for Reproductive Stage Salinity Tolerance in Rice from BRRI dhan 47. Plant Breed. Biotechnol. 2019, 7, 302–312. [Google Scholar] [CrossRef]
- Mohammadi, R.; Mendioro, M.S.; Diaz, G.Q.; Gregorio, G.B.; Singh, R.K. Mapping quantitative trait loci associated with yield and yield components under reproductive stage salinity stress in rice (Oryza sativa L.). J. Genet. 2013, 92, 433–443. [Google Scholar] [CrossRef]
- Krishnamurthy, S.L.; Lokeshkumar, B.M.; Rathor, S.; Warraich, A.S.; Yadav, S.; Gautam, R.K.; Singh, R.K.; Sharma, P.C. Development of Salt-Tolerant Rice Varieties to Enhancing Productivity in Salt-Affected Environments. Environ. Sci. Proc. 2022, 16, 30. [Google Scholar] [CrossRef]
- Mei, H.W.; Xu, J.L.; Li, Z.K.; Yu, X.Q.; Guo, L.B.; Wang, Y.P.; Ying, C.S.; Luo, L.J. QTLs influencing panicle size detected in two reciprocal introgressive line (IL) populations in rice (Oryza sativa L.). Theor. Appl. Genet. 2006, 112, 648–656. [Google Scholar] [CrossRef]
- Xiao, J.; Li, J.; Yuan, L.; Tanksley, S.D. Identification of QTLs affecting traits of agronomic importance in a recombinant inbred population derived from a subspecific rice cross. Theor. Appl. Genet. 1996, 92, 230–244. [Google Scholar] [CrossRef] [PubMed]
- Ueda, K.; Yoshimura, F.; Miyao, A.; Hirochika, H.; Nonomura, K.-I.; Wabiko, H. COLLAPSED ABNORMAL POLLEN1 Gene Encoding the Arabinokinase-Like Protein Is Involved in Pollen Development in Rice. Plant Physiol. 2013, 162, 858–871. [Google Scholar] [CrossRef]
- Cho, Y.-G.; Kang, H.-J.; Lee, J.-S.; Lee, Y.-T.; Lim, S.-J.; Gauch, H.; Eun, M.-Y.; McCouch, S.R. Identification of Quantitative Trait Loci in Rice for Yield, Yield Components, and Agronomic Traits across Years and Locations. Crop Sci. 2007, 47, 2403–2417. [Google Scholar] [CrossRef]
- Ren, Z.-H.; Gao, J.-P.; Li, L.-G.; Cai, X.-L.; Huang, W.; Chao, D.-Y.; Zhu, M.-Z.; Wang, Z.-Y.; Luan, S.; Lin, H.-X. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, P.; Hu, F.; Zhou, J.; Li, J.; Deng, X.; Ren, G.; Li, F.; Tao, D. A new sterile gene from Oryza glaberrima on chromosome 3. Rice Genet. Newsl. 2005, 22, 26–29. [Google Scholar]
- Zhenbo, T.; Lishuang, S.; Zuolian, Y.; Chaofu, L.; Ying, C.; Kaida, Z.; Lihuang, Z. Identification of QTLs for ratooning ability and grain yield traits of rice and analysis of their genetic effects. Zuo Wu Xue Bao 1997, 23, 289–295. [Google Scholar]
- IRRI. Standardization Evaluation System for Rice; International Rice Research Institute: Makati, Philippines, 2013; p. 34. [Google Scholar]
- Doyle, J. DNA Protocols for Plants. In Molecular Techniques in Taxonomy; Hewitt, G.M., Johnston, A.W.B., Young, J.P.W., Eds.; Springer Berlin Heidelberg: NATO ASI Series; Springer: Berlin/Heidelberg, Germany, 1991; Volume 57, pp. 283–293. [Google Scholar] [CrossRef]
- Golicz, A.A.; Bayer, P.E.; Edwards, D. Skim-Based Genotyping by Sequencing. Methods Mol. Biol. 2015, 1245, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chebotarov, D.; Kudrna, D.; Llaca, V.; Lee, S.; Rajasekar, S.; Mohammed, N.; Al-Bader, N.; Sobel-Sorenson, C.; Parakkal, P.; et al. A platinum standard pan-genome resource that represents the population structure of Asian rice. Sci. Data 2020, 7, 113. [Google Scholar] [CrossRef]
- Van Os, H.; Stam, P.; Visser, R.G.F.; van Eck, H.J. SMOOTH: A statistical method for successful removal of genotyping errors from high-density genetic linkage data. Theor. Appl. Genet. 2005, 112, 187–194. [Google Scholar] [CrossRef]
- Churchill, G.A.; Doerge, R.W. Empirical threshold values for quantitative trait mapping. Genetics 1994, 138, 963–971. [Google Scholar] [CrossRef]
- McCouch, S.R.; Cho, Y.G.; Yano, M.; Paul, E.; Blinstrub, M.; Kinoshita, T. Report on QTL nomenclature. Rice Genet. Newsl. 1997, 14, 11–13. [Google Scholar]
- McCouch, S.R.; CGSNL (Committee on Gene Symbolization, Nomenclature and Linkage, Rice Genetics Cooperative). Gene nomenclature system for rice. Rice 2008, 1, 72–84. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Chr. | cM | QTL 1 | Flanking Markers | LOD | PVE (%) | Additive Effect | Allele Effect |
|---|---|---|---|---|---|---|---|---|
| Plant height | 1 | 138.7 | qPH1.1 | CM020682.1_33842918_1; CM020682.1_34965920_1 | 14.9 | 27.0 | −22.46 | Hasawi |
| 2 | 91.2 | qPH2.1 | CM020683.1_22803914_2; CM020683.1_23139808_2 | 7.5 | 9.6 | −12.36 | Hasawi | |
| 4 | 34.4 | qPH4.1 | CM020685.1_6844517_4; CM020685.1_8787440_4 | 3.1 | 3.9 | −9.00 | Hasawi | |
| Productive tillers | 2 | 7.2 | qPT2.1 | CM020683.1_1744356_2; CM020683.1_2059119_2 | 7.0 | 11.2 | −0.01 | Hasawi |
| 3 | 27.1 | qPT3.1 | CM020684.1_6731611_3; CM020684.1_6810365_3 | 6.1 | 11.9 | −0.34 | Hasawi | |
| 6 | 118.9 | qPT6.1 | CM020687.1_29137956_6; CM020687.1_29925158_6 | 4.8 | 11.7 | 3.79 | BRRI dhan28 | |
| 8 | 31.2 | qPT8.1 | CM020689.1_7802407_8′ CM020689.1_8320104_8 | 4.2 | 5.6 | 4.12 | BRRI dhan28 | |
| 11 | 26.4 | qPT11.1 | CM020692.1_6481151_11; CM020692.1_6638823_11 | 7.2 | 13.4 | 3.23 | BRRI dhan28 | |
| 11 | 20.4 | qPT11.2 | CM020692.1_4890980_11; CM020692.1_5380140_11 | 3.6 | 8.6 | 0.37 | BRRI dhan28 | |
| No. filled spikelets | 3 | 27.1 | qNFS3.1 | CM020684.1_6731611_3; CM020684.1_6810365_3 | 6.6 | 17.9 | −6.26 | Hasawi |
| 3 | 87.1 | qNFS3.2 | CM020684.1_21729110_3; CM020684.1_21917452_3 | 3.9 | 8.2 | −241.51 | Hasawi | |
| 6 | 97.9 | qNFS6.1 | CM020687.1_24336222_6; CM020687.1_24647518_6 | 3.6 | 11.5 | 12.88 | BRRI dhan28 | |
| 7 | 52.8 | qNFS7.1 | CM020688.1_12463596_7; CM020688.1_13718255_7 | 3.5 | 17.4 | −129.05 | Hasawi | |
| 8 | 4.2 | qNFS8.1 | CM020689.1_949460_8; CM020689.1_1275124_8 | 3.8 | 16.6 | 63.01 | BRRI dhan28 | |
| 11 | 76.4 | qNFS11.1 | CM020692.1_18857719_11; CM020692.1_19789359_11 | 4.0 | 12.3 | −509.21 | Hasawi | |
| 12 | 82.2 | qNFS12.1 | CM020693.1_19382691_12; CM020693.1_21580067_12 | 3.0 | 15.0 | −435.44 | Hasawi | |
| No. unfilled spikelets | 2 | 10.2 | qNUFS2.1 | CM020683.1_2221957_2; CM020683.1_2585844_2 | 5.3 | 11.4 | −229.78 | Hasawi |
| Percent filled spikelets | 4 | 9.4 | qPFS4.1 | CM020685.1_2019282_4; CM020685.1_3262402_4 | 4.3 | 14.3 | 11.78 | BRRI dhan28 |
| 4 | 99.4 | qPFS4.2 | CM020685.1_24786787_4; CM020685.1_25615450_4 | 3.6 | 6.5 | 4.60 | BRRI dhan28 | |
| 6 | 111.9 | qPFS6.1 | CM020687.1_26815088_6; CM020687.1_28217292_6 | 7.8 | 14.9 | 7.55 | BRRI dhan28 | |
| Grain yield | 1 | 123.7 | qGY1.1 | CM020682.1_30842423_1; CM020682.1_31267467_1 | 6.2 | 7.8 | −0.44 | Hasawi |
| 3 | 27.1 | qGY3.1 | CM020684.1_6731611_3; CM020684.1_6810365_3 | 7.3 | 11.6 | −0.10 | Hasawi | |
| 3 | 87.1 | qGY3.2 | CM020684.1_21729110_3; CM020684.1_21917452_3 | 4.5 | 5.8 | −4.33 | Hasawi | |
| Grain yield | 3 | 2.1 | qGY3.3 | CM020684.1_510306_3; CM020684.1_528354_3 | 3.6 | 4.3 | 2.62 | BRRI dhan28 |
| 4 | 98.4 | qGY4.1 | CM020685.1_24368860_4; CM020685.1_24786787_4 | 4.3 | 10.0 | −1.16 | Hasawi | |
| 6 | 42.9 | qGY6.1 | CM020687.1_10644021_6; CM020687.1_10866598_6 | 7.1 | 19.2 | 0.32 | BRRI dhan28 | |
| 6 | 106.9 | qGY6.2 | CM020687.1_24647518_6; CM020687.1_26815006_6 | 4.5 | 5.0 | 2.09 | BRRI dhan28 | |
| 9 | 75.1 | qGY9.1 | CM020690.1_18345383_9; CM020690.1_19048905_9 | 4.1 | 4.7 | 2.20 | BRRI dhan28 | |
| Na-K ratio | 1 | 163.7 | qNaK1.1 | CM020682.1_40556648_1; CM020682.1_41115782_1 | 12.6 | 1.9 | −3.16 | Hasawi |
| 1 | 171.7 | qNaK1.2 | CM020682.1_42531739_1; CM020682.1_43167854_1 | 11.4 | 1.8 | 0.28 | BRRI dhan28 | |
| 3 | 105.1 | qNaK3.1 | CM020684.1_25664106_3; CM020684.1_27320499_3 | 11.0 | 1.9 | 0.34 | BRRI dhan28 | |
| 3 | 119.1 | qNaK3.2 | CM020684.1_29467527_3; CM020684.1_30649884_3 | 28.8 | 3.0 | 0.23 | BRRI dhan28 | |
| 4 | 105.4 | qNaK4.1 | CM020685.1_26212611_4; CM020685.1_26658501_4 | 10.9 | 1.9 | 0.00 | BRRI dhan28 | |
| 4 | 141.4 | qNaK4.2 | CM020685.1_34446513_4; CM020685.1_36125683_4 | 12.9 | 1.8 | 0.13 | BRRI dhan28 | |
| 5 | 79.7 | qNaK5.1 | CM020686.1_18974331_5; CM020686.1_20934712_5 | 13.6 | 1.8 | −3.13 | Hasawi | |
| 6 | 118.9 | qNaK6.1 | CM020687.1_29137956_6; CM020687.1_29925158_6 | 11.0 | 1.9 | −0.27 | Hasawi | |
| 7 | 110.8 | qNaK7.1 | CM020688.1_27366179_7; CM020688.1_28119385_7 | 10.7 | 1.9 | −0.18 | Hasawi | |
| 9 | 3.1 | qNaK9.1 | CM020690.1_371439_9; CM020690.1_1222229_9 | 12.1 | 1.7 | −3.32 | Hasawi | |
| 11 | 56.4 | qNaK11.1 | CM020692.1_13310544_11; CM020692.1_14431043_11 | 11.6 | 1.9 | −3.03 | Hasawi | |
| 12 | 7.2 | qNaK12.1 | CM020693.1_1735537_12; CM020693.1_1862734_12 | 9.2 | 1.7 | −3.39 | Hasawi |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mondal, S.; Septiningsih, E.M.; Singh, R.K.; Thomson, M.J. Mapping QTLs for Reproductive Stage Salinity Tolerance in Rice Using a Cross between Hasawi and BRRI dhan28. Int. J. Mol. Sci. 2022, 23, 11376. https://doi.org/10.3390/ijms231911376
Mondal S, Septiningsih EM, Singh RK, Thomson MJ. Mapping QTLs for Reproductive Stage Salinity Tolerance in Rice Using a Cross between Hasawi and BRRI dhan28. International Journal of Molecular Sciences. 2022; 23(19):11376. https://doi.org/10.3390/ijms231911376
Chicago/Turabian StyleMondal, Sejuti, Endang M. Septiningsih, Rakesh K. Singh, and Michael J. Thomson. 2022. "Mapping QTLs for Reproductive Stage Salinity Tolerance in Rice Using a Cross between Hasawi and BRRI dhan28" International Journal of Molecular Sciences 23, no. 19: 11376. https://doi.org/10.3390/ijms231911376