


Mechanisms of Vernalization-Induced Flowering in Legumes

Mathematical Biology and Bioinformatics Laboratory, Peter the Great St. Petersburg Polytechnic University, Polytechnicheskaya, 29, 195251 St. Petersburg, Russia
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(17), 9889; https://doi.org/10.3390/ijms23179889
Submission received: 4 August 2022
/
Revised: 25 August 2022
/
Accepted: 26 August 2022
/
Published: 31 August 2022
(This article belongs to the Special Issue A Commemorative Issue in Honor of 200th Anniversary of the Birth of Gregor Johann Mendel: The Genius of Genetics)
Abstract
:Vernalization is the requirement for exposure to low temperatures to trigger flowering. The best knowledge about the mechanisms of vernalization response has been accumulated for Arabidopsis and cereals. In Arabidopsis thaliana, vernalization involves an epigenetic silencing of the MADS-box gene FLOWERING LOCUS C (FLC), which is a flowering repressor. FLC silencing releases the expression of the main flowering inductor FLOWERING LOCUS T (FT), resulting in a floral transition. Remarkably, no FLC homologues have been identified in the vernalization-responsive legumes, and the mechanisms of cold-mediated transition to flowering in these species remain elusive. Nevertheless, legume FT genes have been shown to retain the function of the main vernalization signal integrators. Unlike Arabidopsis, legumes have three subclades of FT genes, which demonstrate distinct patterns of regulation with respect to environmental cues and tissue specificity. This implies complex mechanisms of vernalization signal propagation in the flowering network, that remain largely elusive. Here, for the first time, we summarize the available information on the genetic basis of cold-induced flowering in legumes with a special focus on the role of FT genes.
1. Introduction
In the winter plant varieties, flowering could be initiated only after prolonged cold exposure or vernalization [1,2,3]. The role of vernalization in plant development is to maintain a vegetative state during winter via the repression of major flowering inductors. This ensures that flowering and sexual reproduction take place in the milder conditions of spring [4]. After the end of the cold treatment, the floral activator genes are de-repressed. This leads to their activation by the photoperiod pathway and floral transition [5].
The best knowledge on the cold-induced flowering has been accumulated for Arabidopsis and cereals. In Arabidopsis thaliana, the response to vernalization is mostly mediated through the MADS-box gene FLOWERING LOCUS C (FLC) [6,7]. FLC is a repressor that delays flowering by preventing the expression of floral activators [8]. Vernalization treatment represses FLC and releases flowering promotion. It has been shown that the major targets of FLC in Arabidopsis are FLOWERING LOCUS T (FT) and SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1) [9,10].
The Arabidopsis FT gene functions as an integrator, converging information from the photoperiodic, vernalization and autonomous regulatory pathways [11]. The interplay between activating and repressive inputs from different pathways is controlled through a complex cis-regulatory region [12]. The homologues of the FT gene are functioning in a variety of plant species [13,14,15,16,17,18,19,20,21,22,23].
FT encodes a major florigen which promotes transition from vegetative growth to flowering [9,24]. A protein product of the FT gene accumulates information on environmental signals in the leaves, and then moves to the shoot apex to transfer this information to meristem identity genes, including the key genes APETALA1 (AP1) and LEAFY (LFY) [25,26,27,28,29,30]. These genes function to convert meristem to the reproductive state. FT activates LFY expression through the MADS-domain transcription factor SOC1 [5,31,32]. Moreover, AP1 and LFY act as mutual transcriptional activators [33].
An inductive function of the FT gene product is opposed by the TERMINAL FLOWER 1 (TFL1) gene, which represses flowering and controls inflorescence architecture via the down-regulation of AP1 and LFY [34,35,36,37].
In legumes, vernalization sensitivity, inherent to wild germplasm, has been mostly lost from cultivated genotypes due to the long-standing breeding efforts aimed at creating the early-flowering spring cultivars. Nevertheless, some cultivated genotypes still retain a low level of vernalization response [38,39]. Understanding the genetic bases of cold-induced flowering in legumes enables one to decipher the origin of adaptation to different environments, history of domestication, and evolution of flowering time networks. However, the molecular mechanisms of vernalization response in legumes remain largely obscure.
Remarkably, the FLC gene is missing in many legume species. This comprises all galegoid legumes, including Medicago truncatula, Pisum sativum, Vicia faba, Lens culinaris and Cicer arietinum, and genistoid legumes such as Lupinus angustifolius, Lupinus luteus and Lupinus albus [40,41]. Despite the absence of the FLC gene, these species possess a vernalization-based flowering promotion through the activation of FT genes (Table 1) [42,43]. The published results suggest that FT genes are the main targets of vernalization in legumes (Table 2), although the mechanisms of FT activation are still unclear [44,45]. So far, the most intensive research of cold-induced flowering has been conducted in Medicago trancatula and narrow-leafed lupin Lupinus angustifolius [44,45,46,47]. Here, we summarize the available information for genetic bases of vernalization in eight legume species with a special emphasis on the role of FT genes.
2. The Mechanism of Cold–Induced Flowering in Arabidopsis
Prior to vernalization, flowering promotion in Arabidopsis is repressed by the MADS-box transcription factor FLC, which interacts with the first intron of the FT gene and the promoter region of SOC1 gene [8]. FLC represses FT expression in the leaves, while SOC1 is down-regulated in the shoot apex [8,11].
The direct targets of FLC include SHORT VEGETATIVE PHASE (SVP) [59], TEMPRANILLO 1 (TEM1) [60,61], SQUAMOSA PROMOTER-BINDING-LIKE PROTEIN 15 (SPL15) [62,63], and a few more flowering-related genes (summarized in [64]).
FLC expression is up-regulated by the FRIGIDA (FRI) gene through a FRI-containing supercomplex, which establishes a local chromosomal environment for a high-level production of FLC mRNAs [65,66].
The vernalization treatment leads to the epigenetic silencing of FLC, which involves multiple factors. These factors include (1) the components of autonomous pathway; (2) the long noncoding RNAs (lncRNAs) produced by FLC locus (COOLAIR complex); (3) Polycomb repressive complex 2 (PRC2), including the core components VERNALIZATION2 (VRN2), FERTILIZATION INDEPENDENT ENDOSPERM (FIE), MULTICOPY SUPPRESSOR OF IRA1 (MSI1) and SWINGER (SWN); (4) the plant homeodomain (PHD) proteins VERNALIZATION INSENSITIVE 3 (VIN3), VERNALIZATION 5 (VRN5) and VIN3-LIKE2. The repression of FLC involves switching chromatin to a repressed state. The PHD–PRC2 complex acts to deposit the repressive epigenetic mark H3K27me3 across the FLC locus, which provides maintenance of FLC repression after the return to warm conditions. FLC repression proceeds progressively via the cell-autonomous mechanism, and a stable down-regulation of FLC is achieved after the prolonged cold exposure, which occurs during winter [6,67,68,69,70,71,72,73,74,75,76].
The mechanisms of FLC silencing and epigenetic memory in Arabidopsis have been widely studied and summarized in the recent reviews [3,77,78].
Following the release of repression by FLC, the FT and SOC1 genes become activated by the photoperiodic pathway, leading to flowering promotion [79,80,81,82]. A generalized mechanism of the cold-induced flowering transition in Arabidopsis is shown in Figure 1a.
Extensive research revealed that the flowering-related genes are largely conserved between Arabidopsis and legumes [42,44,45,49,53,54,55,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97]. However, the legumes generally lack any FLC orthologs, except for soybean, a vernalization-independent species, which retained one FLC copy in its paleopolyploid genome [42,98,99].
3. FT Genes in Legumes
Despite the absence of FLC orthologs, the legume FT genes retain the role of major vernalization signal integrators (Table 2).
Legumes have three subclades of FT genes (FTa, FTb and FTc) with five Arabidopsis FT orthologs detected in pea Pisum sativum, and six orthologs in Medicago trancatula and Lens culinaris (Table 1) [44,45,53,55,56,100]. Unlike pea and Medicago, chickpea Cicer arietinum has a single FTb gene, resulting in a family of five genes (Table 1) [54].
Similarly to the above described legume species, faba bean Vicia faba presumably has five or six FT genes, however, their number and identity are yet to be determined [57].
The narrow-leafed lupin Lupinus angustifolius is a member of the genistoid clade, which is the most basal clade of papilionoid legumes [45,101]. In Lupinus angustifolius, the FTb subclade is completely missing, while there are two FTa genes and two FTc genes (Table 1). Phylogenetic analysis revealed that lupin FTa1 and FTa2 genes correspond to the Medicago FTa3 subclade [45]. Lupin FTa and FTc genes presumably originated from single copies by lineage-specific duplication [90].
Interestingly, the members of different FT subclades in pea and Medicago showed distinct patterns of regulation with respect to environmental cues and tissue specificity. For example, FTa and FTb genes are expressed in leaves, while FTc genes function in the shoot apex and may contribute to the integration of FTa and FTb signals [43,44,53]. Moreover, several studies point to the existence of cross-regulation between different members of the FT family [44,53]. This demonstrates the complexity of regulatory networks involving various FT family members in legumes, including those implicated in the vernalization response (Figure 1).
4. A Role of the Legume FT Genes in Vernalization Response
4.1. Medicago
The study by Laurie et al. provided the first clue on the role of FT family genes in the vernalization response in legumes [44]. It was shown that FTa1 mutants in Medicago trancatula are late flowering and have the reduced ability of vernalization response [44,100]. Remarkably, despite vernalization insensitivity, the MtFTa1 mutants retained the photoperiod response [44].
The experiments on MtFTa1 overexpression showed the early flowering phenotype in unvernalized plants, resembling that of wild type plants after vernalization. Interestingly, the expression of the M. trancatula FTc gene was significantly reduced in MtFTa1 mutants, suggesting that the MtFTc gene might be downstream of MtFTa [44].
An insertion of Tnt1, a long terminal repeat (LTR) retrotransposon from tobacco, into FTa1 3rd intron or in 3′ region resulted in the early flowering phenotype (“spring” mutants), similar to that after FTa1 overexpression [46,48]. This insertion eliminated vernalization response, but resulted in a strong photoperiod sensitivity. These data suggest that the MtFTa1 gene may have evolved separate mechanisms of response to vernalization and photoperiod.
Several transcription factors, such as SOC1 and FRUITFULL (FUL) showed elevated expression in spring mutants, which is consistent with their downstream position relative to FT signaling in Arabidopsis [46,48,100]. The Arabidopsis FUL gene encodes a MADS-box protein and acts as a flowering-time and meristem-identity gene, closely related to AP1 and CAULIFLOWER (CAL) [102]. FUL is involved in an up-regulation of the LFY gene. The MtSOC1 genes demonstrated up-regulation by both vernalization and photoperiod, which was reduced in FTa1 mutants, providing evidence that these genes are downstream FTa1 [49].
An essential role of FTa1 in Medicago was confirmed by a recent RNAseq assay showing a wide range of genes differentially regulated by mutation of MtFTa1 [96]. Upon floral transition, the MtFTa1 targets included genes from photoperiod, gibberellin-related and age-related pathways.
In Arabidopsis, the FT protein acts in complex with the basic domain/leucine zipper (bZIP) transcription factor FLOWERING LOCUS D (FD), and the model of FT-FD interaction is conserved in different plant species [89,103,104,105,106,107]. The FT-FD heterodimer is responsible for activation of AP1 [25,106], FUL and SEPALLATA3 (SEP3) [108]. Interestingly, the mutation in the MtFDa gene did not abolish vernalization responsiveness, unlike the mutation in MtFTa. Nevertheless, the double mutation of MtFDa and MtFTa1 completely blocked floral transition pointing to the complementary roles of these two genes [96].
4.2. Narrow-Leafed Lupin
Studies on the narrow-leafed lupin (Lupinus angustifolius) revealed that of all FT genes, only LanFTc1 shows strong vernalization response [45]. The expression of this gene was repressed in the unvernalized conditions, but possessed strong induction following cold treatment.
This is the first case of the FTc subclade gene involvement in vernalization response [45]. Of all FT subclades, FTc genes are the most divergent and differ from most other FT genes by substitution of several conserved residues [109].
It has been shown that several natural mutations (Ku, Jul and Pal) provide vernalization independence in L. angustifolius. All these mutations are located in the promoter region of the LanFTc1 gene [45,47,90,110].
Ku and Jul are naturally occurring dominant mutations, independently discovered in cultivars “Borre” and “Krasnolistny” in 1960s, which determine vernalization insensitivity and early flowering of the cultivated lupin [110,111,112]. Ku and Jul are deletions within the LanFTc1 promoter region, spanning 1423 and 5162 bp respectively. Another 1208 bp deletion, partially overlapping with Ku and Jul, has been revealed in the wild germplasm from Palestine, and termed Pal mutation. Pal is associated with a partial vernalization responsiveness and slightly delayed flowering [110].
The analysis of LanFTc1 expression in the three deletion variants confirmed their importance for flowering time regulation via the vernalization pathway. Remarkably, Ku and Jul deletions appeared to be functionally equivalent, resulting in the similar expression profile of LanFTc1 gene. Therefore, it has been suggested that major regulatory elements responsible for LanFTc1 vernalization sensitivity, confer to a region of Ku deletion, namely, the 1423 bp sequence [110]. The functional activity of 1208 bp Pal deletion presumably refines this major regulatory region.
An effect of Ku, Jul and Pal mutations in the narrow-leafed lupin is very similar to that of “spring” mutations in Medicago, suggesting a common mechanism of vernalization response by de-repression of FT genes [46]. In both cases, current research provides evidence about the genomic regions, containing important binding sites, which enable transcriptional repression of the FT genes in wild type. However, in both Medicago and lupin, it is unknown which elements within these regions are responsible for the FT repression in the absence of vernalization and its de-repression after vernalization [45,110].
The promoter region of the L. angustifolius LanFTc1 gene contains putative binding sites for the homologues of many transcription factors regulating FT expression in Arabidopsis [45,110]. This region has been divided into two zones. The deletion of the first zone provided a high level of LanFTc1 expression, while the deletion of the second zone resulted in the moderately high expression levels. However, the function of these sequences in wild type remains unclear [110]. In Arabidopsis, the FT promoter contains four major blocks, including the ID block with two insertions/deletions (indels) affecting promoter efficiency [12,113]. A position of the ID block within the Arabidopsis FT promoter is similar to a position of the 1423 bp Ku deletion in L. angustifolius LanFTc1 promoter. However, the Ku region showed no sequence conservation with the ID block [90,110].
Besides LanFTc1, recent studies have shown the involvement of a number of novel candidate genes in the vernalization response [47,111]. The expression profiles of these genes were examined for vernalization responsiveness in three accessions, carrying domesticated allele Ku, intermediate allele Pal, and wild allele ku [47].
LanAGL8 (AGAMOUS-LIKE 8) is an L. angustifolius ortholog of the A. thaliana FUL gene [115,116,117]. In L. angustifolius, the expression pattern and vernalization responsiveness of LanAGL8 were very similar to those of LanFTc1. The authors suggested that LanAGL8 could act downstream of LanFTc1. As described above, an expression of FUL in Medicago also pointed to its downstream position relative to MtFTa1 gene [46,48].
LanFD, the L. angustifolius ortholog of the Arabidopsis FD gene, showed vernalization responsiveness, which varied between early and late flowering genotypes. Since the protein product of FD gene in Arabidopsis acts in a complex with the FT protein [103], a variation in FD expression could modulate an effect of the FT signal [47].
A recent study provided transcriptomic evidence on the role of LanCRLK1 and LanUGT85A2 genes, acting in C-repeat binding factor (CBF) cold responsiveness and UDP-glycosyltransferase pathways, in the LanFTc1-mediated vernalization response [114]. Nevertheless, the analysis of LanCRLK1 and LanUGT85A2 expression profiles did not provide convincing evidence for the role of these genes in the vernalization response of L. angustifolius [47].
4.3. Yellow Lupin
Despite the similar domestication history of narrow-leafed lupin and yellow lupin, (Lupinus luteus), the genetics of domestication traits have been studied much more intensively in the narrow-leafed lupin [50,118,119]. The studies on QTL mapping of domestication syndrome and flowering time traits in L. luteus were first published in 2020 [50,120].
4.4. White Lupin
Remarkably, unlike narrow-leafed and yellow lupin, in the white lupin Lupinus albus the vernalization response was revealed as a continuous trait suggesting polygenic regulation [52,121]. Despite the involvement of different number of genes (single genes vs several QTLs), the genomes of white and narrow-leafed lupin showed high collinearity [121].
Genotyping with PCR-based markers highlighted major candidates for the vernalization response and early flowering QTLs [51,52,121]. The QTL analyses revealed significant correlations between time to flowering and polymorphism in markers anchored in the sequences of regulatory genes. These genes are slightly divergent between two studies involving mapping population and germplasm collection [51,52]. and include L. albus orthologs of FTa1, FTc1, SEP3, EARLY FLOWER 1 (ELF1), FLOWERING LOCUS D (FLD), FRIGIDA (FRI) CONSTANS (CO), FY, MOTHER OF FT AND TFL1 (MFT), PHYTO-CHROME INTERACTING FACTOR4 (PIF4), SKI-INTERACTING PROTEIN 1 (SKIP1), and VERNALIZATION INDEPENDENCE 3 (VIP3) [51,52]. Remarkably, both studies highlighted a possible role of the FRI gene, a major component of the vernalization pathway, which is responsible for activation of FLC expression (see above) [65]. In Arabidopsis, an allelic variation of FRI accounts for the majority of natural variation in flowering time [122,123].
Overall, these studies highlight a complex nature of flowering control in white lupin, with candidate genes dispersed among numerous loci. This contrasts with rather simple mechanisms of cold-induced flowering in L. angustifolius and L. luteus.
4.5. Garden Pea
In the garden pea Pisum sativum, the FTa1 gene, corresponding to the GIGAS locus described in earlier studies [124], plays a major role in flowering induction [53]. The gigas mutants demonstrated late flowering in all photoperiodic conditions tested, nevertheless, with an unambiguous effect of vernalization on flowering time [124]. However, such an effect was observed in the specific tissues, suggesting that FTa1/GIGAS may respond to vernalization in a tissue-specific manner [53,124,125]. Acquisition of more molecular data is required to analyze mechanisms of vernalization-induced flowering in pea.
4.6. Chickpea
In chickpea Cicer arietinum, a major QTL providing 55% of the phenotypic variation for vernalization response trait has been identified on the linkage group 3 (LG3) of the chickpea genetic map [126]. LG3 on chromosome 3 was earlier reported to harbor flowering time genes by many authors [127,128,129,130,131]. Thus, LG3 harbors several QTLs for flowering time and vernalization response in chickpea [126].
A recent study detected a strong genetic association between early flowering and a cluster of FT genes on chromosome 3, comprising FTa1, FTa2 and FTc [54]. This cluster has been located to the same genomic region as the QTL for vernalization response [126]. Nevertheless, it is unknown, which of these FT genes might be responsible for the effects on vernalization response.
4.7. Lentil
A recent QTL analysis revealed that the DTF6a locus on lentil (Lens culinaris) chromosome 6 strongly contributes to early flowering of the Indian landrace ILL 2601 [55].
The detailed inspection of the DTF6a region revealed that it corresponds to the syntenic regions on Medicago chromosome 7, pea chromosome 5 and chickpea chromosome 3. This region harbors a conserved cluster of FT genes, including two FTa genes (FTa1 and FTa2) and an FTc gene, located adjacent or nearby [44,53,54,132].
Further analysis showed that DTF6a deletion is associated with elevated expression of FTa1 and FTa2 genes. The sequences of FTa1-FTa2 cluster were compared between the early flowering lentil accession ILL 2601 and the late flowering accession ILL 5588. There were no differences in the coding regions of either gene, but the comparison of non-coding regions identified 136 nucleotide polymorphisms (SNPs) and 25 indels distinguishing two accessions. The most substantial difference was the 7441 bp deletion, comprising most of the FTa1–FTa2 intergenic region in the early ILL 2601 accession [55].
The analysis of vernalization response showed that the late ILL 5588 accession was vernalization-sensitive under long and short photoperiods. On the contrary, ILL 2601 did not show any vernalization-induced acceleration of flowering.
The results suggested that FTa1 (LcFTa1) is the most prominent candidate for the early flowering and vernalization insensitivity of the early ILL 2601 accession [55]. As described above, FTa1 is responsible for early flowering in Medicago and pea [44,53]. A large 7441 bp indel detected in the FTa1–FTa2 intergenic of the early ILL 2601 accession might include regulatory elements required for LcFTa1 repression [55]. In this case a mechanism, which confers early flowering and vernalization independence in lentil, should resemble the above described Ku mutations in the promoter region of the FTc1 gene in L. angustifolius [45] and transposon insertions in the third intron or 3′ untranslated region of the FTa1 gene in M. trancatula [46].
4.8. Faba Bean
In faba bean Vicia faba, the candidate gene mapping and the quantitative trait loci (QTL) analysis revealed the cluster of FT loci close to the most conserved flowering time QTL on chromosome 5 in a region syntenic with a section of Medicago chromosome 7 containing FTa and FTc genes [58]. However, the authors failed to map individual FT genes within this region due to high sequence conservation [58].
In another recent study, the single-molecule, real-time (SMRT) sequencing revealed 50 flowering-related genes that could be associated with vernalization. The vernalization-responsive transcripts showed significant up-regulation in the cold-treated samples [57].
The expression dynamics of one candidate gene, an ortholog of SOC1, was examined by RT-PCR. Phylogenetic analysis demonstrated that VfSOC1 is most similar to MtSOC1c in Medicago (86.5% identity). Under a low temperature treatment, the levels of VfSOC1 in a leaf showed elevation on day 7 and steadily increased thereafter. The ectopic expression of VfSOC1 in Arabidopsis could promote earlier flowering. These results point to the important role of VfSOC1 in vernalization response of faba bean [57].
5. Candidates for the Vernalization-Repressed Repressors in Legumes
The repressors of flowering, down-regulated by cold exposure, have yet to be uncovered in legumes. As a starting point, current research aims to analyze legume orthologs of the Arabidopsis genes involved in the regulation of FLC locus.
FRI is a major gene controlling vernalization response in Arabidopsis. FRI suppresses flowering mostly through the upregulation of FLC [7]. The analysis of A. thaliana populations revealed that the allelic variation of FRI accounts for approximately 70% of flowering time variation [123]. In the white lupin L. albus, FRI has been recently highlighted as a candidate gene underlying the early flowering QTL [51,52].
As described above, PHD proteins VIN3 and VRN5 are the components of PHD–PRC2 complex, required for the stable repression of FLC following vernalization in Arabidopsis [70,74,133]. Despite their similar function, VIN3 and VRN5 genes differ in the expression dynamics and the vernalization responsiveness [133]. There are three copies of each gene in the genome of narrow-leaved lupine L. angustifolius. The expression of LanVIN3 and LanVRN5 genes was different from that of Arabidopsis with significantly higher levels of LanVIN3 compared to LanVRN5 [47]. However, the differences in the expression profiles and vernalization sensitivety of the LanVIN3 and LanVRN5 genes did not match the observed differences in time to flowering. Therefore, the authors suggested that these genes are unlikely to be involved in the vernalization response [47].
The VRN2 protein is a core component of Polycomb repressive complex 2 (PRC2) in Arabidopsis. Phylogenetic analysis revealed the VRN2-like genes in some legume species including Medicago, pea and lupin [67]. Interestingly, in Medicago, MtVRN2 gene functions as a repressor of the floral activator FTa1 [134]. Mtvrn2 mutants demonstrated early, vernalization-independent flowering, as well as an elevated expression of FTa1 and its target genes, including SOC1a, FULb and AP1. Thus, in the absence of the FLC clade of flowering time repressors in Medicago, MtVRN2 apparently plays a role in silencing major flowering activators [134].
Nevertheless, the “memory of the cold”, or prolonged activation of flowering after vernalization, is preserved in M. trancatula. MtFTa1 gene was not expressed in the germinated seedlings after 14 days in the cold. However, its expression started immediately after plants were transferred to warm conditions, and the expression levels increased after 7 and 14 days in the warmth [44]. The mechanisms of such memory of the cold exposure are yet unknown.
6. Conclusions
Vernalization is a widespread mechanism of flowering time regulation in annual plants from temperate regions. However, the vernalization pathways appear to have evolved independently in various plant lineages, leading to differences of gene functions and regulatory pathways across groups [1,109]
Since the functions of VIN3, VRN5 and VRN2 genes in legumes differ from those in Arabidopsis, and the FLC gene is absent, it is obvious that the cold-induced de-repression of flowering activators proceeds via different mechanisms. There could be several scenarios of vernalization-based FT induction in legumes.
For example, in cereals, vernalization involves the down-regulation of the FT repressor, which is not orthologous to FLC in Arabidopsis [135,136]. On the other hand, in the sugar beet Beta vulgaris, vernalization represses an FT gene (BvFT1), which down-regulates another FT gene (BvFT2) [137].
Since the MtVRN2 gene in Medicago performs a function opposite to that in Arabidopsis, namely, it represses FTa1 instead of activation [134], the latter scenario is quite plausible. There may also be cross-regulatory interactions between different FT genes, as it has been suggested that the Medicago MtFTc gene acts downstream of MtFTa1 [44]. This suggests a complex network transmitting the vernalization signal from several FT genes to the meristem identity genes AP1 and LFY (Figure 1b). It should be noted that in addition to being activated by the FT orthologs, the legume genes AP1 and LFY are regulated by several copies of the TFL1 gene [53,91,109,138,139]. Future research should explore this regulatory complexity in terms of the vernalization signal propagation.
Author Contributions
Conceptualization, S.Y.S. and M.G.S.; writing—original draft preparation, S.Y.S.; writing—review and editing, S.Y.S. and M.G.S.; visualization, S.Y.S.; funding acquisition, M.G.S. All authors have read and agreed to the published version of the manuscript.
Funding
The research is funded by the Ministry of Science and Higher Education of the Russian Federation under the strategic academic leadership program “Priority 2030” (Agreement 075-15-2021-1333 dated 30 September 2021).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kim, D.; Doyle, M.; Sung, S.; Amasino, R. Winter and the timing of flowering in plants. Annu. Rev. Cell Dev. Biol. 2009, 25, 277–299. [Google Scholar] [CrossRef] [PubMed]
- Ream, T.; Woods, D.; Amasino, R. The molecular basis of vernalization in different plant groups. Cold Spring Harb. Symp. Quant. Biol. 2012, 77, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Geuten, K.; Giri, S.B.; Varma, A. The molecular mechanism of vernalization in Arabidopsis and cereals: Role of Flowering Locus C and its homologs. Physiol. Plant. 2020, 170, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Putterill, J.; Zhang, L.; Yeoh, C.C.; Balcerowicz, M.; Jaudal, M.; Gasic, E.V. FT genes and regulation of flowering in the legume Medicago truncatula. Funct. Plant Biol. 2013, 40, 1199–1207. [Google Scholar] [CrossRef]
- Yoo, S.K.; Chung, K.S.; Kim, J.; Lee, J.H.; Hong, S.M.; Yoo, S.J.; Yoo, S.Y.; Lee, J.S.; Ahn, J.H. CONSTANS activates SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 through FLOWERING LOCUS T to promote flowering in Arabidopsis. Plant Physiol. 2005, 139, 770–778. [Google Scholar] [CrossRef]
- Michaels, S.D.; Amasino, R.M. FLOWERING LOCUS C encodes a novel MADS domain protein that acts as a repressor of flowering. Plant Cell 1999, 11, 949–956. [Google Scholar] [CrossRef]
- Michaels, S.D.; Amasino, R.M. Loss of FLOWERING LOCUS C activity eliminates the late-flowering phenotype of FRIGIDA and autonomous pathway mutations but not responsiveness to vernalization. Plant Cell 2001, 13, 935–941. [Google Scholar] [CrossRef]
- Searle, I.; He, Y.; Turck, F.; Vincent, C.; Fornara, F.; Kröber, S.; Amasino, R.A.; Coupland, G. The transcription factor FLC confers a flowering response to vernalization by repressing meristem competence and systemic signaling in Arabidopsis. Genes Dev. 2006, 20, 898–912. [Google Scholar] [CrossRef]
- Helliwell, C.A.; Wood, C.C.; Robertson, M.; James Peacock, W.; Dennis, E.S. The Arabidopsis FLC protein interacts directly in vivo with SOC1 and FT chromatin and is part of a high-molecular-weight protein complex. Plant J. Cell Mol. Biol. 2006, 46, 183–192. [Google Scholar] [CrossRef]
- Seo, E.; Lee, H.; Jeon, J.; Park, H.; Kim, J.; Noh, Y.S.; Lee, I. Crosstalk between cold response and flowering in Arabidopsis is mediated through the flowering-time gene SOC1 and its upstream negative regulator FLC. Plant Cell 2009, 21, 3185–3197. [Google Scholar] [CrossRef] [Green Version]
- Zeevaart, J.A. Leaf-produced floral signals. Curr. Opin. Plant Biol. 2008, 11, 541–547. [Google Scholar] [CrossRef]
- Adrian, J.; Farrona, S.; Reimer, J.J.; Albani, M.C.; Coupland, G.; Turck, F. cis-Regulatory Elements and Chromatin State Coordinately Control Temporal and Spatial Expression of FLOWERING LOCUS T in Arabidopsis. Plant Cell 2010, 22, 1425–1440. [Google Scholar] [CrossRef]
- Kojima, S.; Takahashi, Y.; Kobayashi, Y.; Monna, L.; Sasaki, T.; Araki, T.; Yano, M. Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant Cell Physiol. 2002, 43, 1096–1105. [Google Scholar] [CrossRef]
- Hou, C.J.; Yang, C.H. Functional analysis of FT and TFL1 orthologs from orchid (Oncidium Gower Ramsey) that regulate the vegetative to reproductive transition. Plant Cell Physiol. 2009, 50, 1544–1557. [Google Scholar] [CrossRef]
- Xu, F.; Rong, X.; Huang, X.; Cheng, S. Recent advances of flowering locus T gene in higher plants. Int. J. Mol. Sci. 2012, 13, 3773–3781. [Google Scholar] [CrossRef]
- Li, C.; Luo, L.; Fu, Q.; Niu, L.; Xu, Z.F. Isolation and functional characterization of JcFT, a FLOWERING LOCUS T (FT) homologous gene from the biofuel plant Jatropha curcas. BMC Plant Biol. 2014, 14, 125. [Google Scholar] [CrossRef]
- Li, D.M.; L, F.B.; Zhu, G.F.; Sun, Y.B.; Liu, H.L.; Liu, J.W.; Wang, Z. Molecular characterization and functional analysis of a Flowering locus T homolog gene from a Phalaenopsis orchid. Genet. Mol. Res. 2014, 13, 5982–5994. [Google Scholar] [CrossRef]
- Guo, D.; Li, C.; Dong, R.; Li, X.; Xiao, X.; Huang, X. Molecular cloning and functional analysis of the FLOWERING LOCUS T (FT) homolog GhFT1 from Gossypium hirsutum. J. Integr. Plant Biol. 2015, 57, 522–533. [Google Scholar] [CrossRef]
- Lei, H.; Su, S.; Ma, L.; Wen, Y.; Wang, X. Molecular cloning and functional characterization of CoFT1, a homolog of FLOWERING LOCUS T (FT) from Camellia oleifera. Gene 2017, 626, 215–226. [Google Scholar] [CrossRef]
- Chaurasia, A.K.; Patil, H.B.; Krishna, B.; Subramaniam, V.R.; Sane, P.V.; Sane, A.P. Flowering time in banana (Musa spp.), a day neutral plant, is controlled by at least three FLOWERING LOCUS T homologues. Sci. Rep. 2017, 7, 5935. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.; Zhang, T.; Guo, T.; Ding, W.; Long, R.; Yang, Q.; Wang, Z. Isolation and Functional Characterization of MsFTa, a FLOWERING LOCUS T Homolog from Alfalfa (Medicago sativa). Int. J. Mol. Sci. 2019, 20, 1968. [Google Scholar] [CrossRef]
- Wu, J.; Wu, Q.; Bo, Z.; Zhu, X.; Zhang, J.; Li, Q.; Kong, W. Comprehensive Effects of Flowering Locus T-Mediated Stem Growth in Tobacco. Front. Plant Sci. 2022, 13, 922919. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Rim, Y.; Cho, H.; Hyun, T.K. Identification and Functional Characterization of FLOWERING LOCUS T in Platycodon grandiflorus. Plants 2022, 11, 325. [Google Scholar] [CrossRef] [PubMed]
- Pin, P.A.; Nilsson, O. The multifaceted roles of FLOWERING LOCUS T in plant development. Plant Cell Environ. 2012, 35, 1742–1755. [Google Scholar] [CrossRef]
- Wigge, P.; Kim, M.; Jaeger, K.; Busch, W.; Schmid, M.; Lohmann, J.; Weigel, D. Integration of spatial and temporal information during floral induction in Arabidopsis. Science 2005, 309, 1056–1059. [Google Scholar] [CrossRef] [PubMed]
- Kardailsky, I.; Shukla, V.K.; Ahn, J.H.; Dagenais, N.; Christensen, S.K.; Nguyen, J.T.; Chory, J.; Harrison, M.J.; Weigel, D. Activation tagging of the floral inducer FT. Science 1999, 286, 1962–1965. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, K.E.; Wigge, P.A. FT protein acts as a long-range signal in Arabidopsis. Curr. Biol. 2007, 17, 1050–1054. [Google Scholar] [CrossRef]
- Chen, Q.; Payyavula, R.S.; Chen, L.; Zhang, J.; Zhang, C.; Turgeon, R. FLOWERING LOCUS T mRNA is synthesized in specialized companion cells in Arabidopsis and Maryland Mammoth tobacco leaf veins. Proc. Natl. Acad. Sci. USA 2018, 115, 2830–2835. [Google Scholar] [CrossRef]
- Schultz, E.A.; Haughn, G.W. LEAFY, a Homeotic Gene That Regulates Inflorescence Development in Arabidopsis. Plant Cell 1991, 3, 771–781. [Google Scholar] [CrossRef]
- Kaufmann, K.; Wellmer, F.; Muiño, J.M.; Ferrier, T.; Wuest, S.E.; Kumar, V.; Serrano-Mislata, A.; Madueño, F.; Krajewski, P.; Meyerowitz, E.M.; et al. Orchestration of floral initiation by APETALA1. Science 2010, 328, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Oh, M.; Park, H.; Lee, I. SOC1 translocated to the nucleus by interaction with AGL24 directly regulates leafy. Plant J. Cell Mol. Biol. 2008, 55, 832–843. [Google Scholar] [CrossRef]
- Lee, J.; Lee, I. Regulation and function of SOC1, a flowering pathway integrator. J. Exp. Bot. 2010, 61, 2247–2254. [Google Scholar] [CrossRef]
- Liljegren, S.J.; Gustafson-Brown, C.; Pinyopich, A.; Ditta, G.S.; Yanofsky, M.F. Interactions among APETALA1, LEAFY, and TERMINAL FLOWER1 specify meristem fate. Plant Cell 1999, 11, 1007–1018. [Google Scholar] [CrossRef]
- Ratcliffe, O.J.; Amaya, I.; Vincent, C.A.; Rothstein, S.; Carpenter, R.; Coen, E.S.; Bradley, D.J. A common mechanism controls the life cycle and architecture of plants. Development 1998, 125, 1609–1615. [Google Scholar] [CrossRef]
- Ratcliffe, O.J.; Bradley, D.J.; Coen, E.S. Separation of shoot and floral identity in Arabidopsis. Development 1999, 126, 1109–1120. [Google Scholar] [CrossRef]
- Hanano, S.; Goto, K. Arabidopsis TERMINAL FLOWER1 is involved in the regulation of flowering time and inflorescence development through transcriptional repression. Plant Cell 2011, 23, 3172–3184. [Google Scholar] [CrossRef]
- Serrano-Mislata, A.; Fernández-Nohales, P.; Doménech, M.J.; Hanzawa, Y.; Bradley, D.; Madueño, F. Separate elements of the TERMINAL FLOWER 1 cis-regulatory region integrate pathways to control flowering time and shoot meristem identity. Development 2016, 143, 3315–3327. [Google Scholar] [CrossRef]
- Sharma, S.; Upadhyaya, H. Vernalization and photoperiod response in annual wild cicer species and cultivated chickpea. Crop. Sci. 2015, 55, 2393–2400. [Google Scholar] [CrossRef]
- Pinhasi van Oss, R.; Sherman, A.; Zhang, H.; Vandemark, G.; Coyne, C.; Abbo, S. Vernalization response of domesticated × wild chickpea progeny is subject to strong genotype by environment interaction. Plant Breed. 2016, 135, 102–110. [Google Scholar] [CrossRef]
- Liew, L.; Singh, M.; Bhalla, P. Unique and conserved features of floral evocation in legumes. J. Integr. Plant Biol. 2014, 56, 714–728. [Google Scholar] [CrossRef]
- Lee, C.; Yu, D.; Choi, H.K.; Kim, R.W. Reconstruction of a composite comparative map composed of ten legume genomes. Genes Genom. 2017, 39, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Hecht, V.; Foucher, F.; Ferrándiz, C.; Macknight, R.; Navarro, C.; Morin, J.; Vardy, M.E.; Ellis, N.; Beltrán, J.P.; Rameau, C.; et al. Conservation of Arabidopsis Flowering Genes in Model Legumes. Plant Physiol. 2005, 137, 1420–1434. [Google Scholar] [CrossRef] [PubMed]
- Weller, J.L.; Ortega, R. Genetic control of flowering time in legumes. Front. Plant Sci. 2015, 6, 207. [Google Scholar] [CrossRef] [PubMed]
- Laurie, R.E.; Diwadkar, P.; Jaudal, M.; Zhang, L.; Hecht, V.; Wen, J.; Tadege, M.; Mysore, K.S.; Putterill, J.; Weller, J.L.; et al. The Medicago FLOWERING LOCUS T Homolog, MtFTa1, Is a Key Regulator of Flowering Time. Plant Physiol. 2011, 156, 2207–2224. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.N.; Książkiewicz, M.; Rychel, S.; Besharat, N.; Taylor, C.M.; Wyrwa, K.; Jost, R.; Erskine, W.; Cowling, W.A.; Berger, J.D.; et al. The loss of vernalization requirement in narrow-leafed lupin is associated with a deletion in the promoter and de-repressed expression of a Flowering Locus T (FT) homologue. New Phytol. 2017, 213, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Jaudal, M.; Yeoh, C.C.; Zhang, L.; Stockum, C.; Mysore, K.S.; Ratet, P.; Putterill, J. Retroelement insertions at the Medicago FTa1 locus in spring mutants eliminate vernalisation but not long-day requirements for early flowering. Plant J. Cell Mol. Biol. 2013, 76, 580–591. [Google Scholar] [CrossRef] [PubMed]
- Rychel-Bielska, S.; Plewinski, P.; Kozak, B.; Galek, R.; Ksiazkiewicz, M. Photoperiod and vernalization control of flowering-related genes: A case study of the narrow-leafed lupin (Lupinus angustifolius L.). Front. Plant Sci. 2020, 11, 572135. [Google Scholar] [CrossRef]
- Yeoh, C.C.; Balcerowicz, M.; Zhang, L.; Jaudal, M.; Brocard, L.; Ratet, P.; Putterill, J. Fine mapping links the FTa1 flowering time regulator to the dominant spring1 locus in Medicago. PLoS ONE 2013, 8, e53467. [Google Scholar] [CrossRef]
- Fudge, J.B.; Lee, R.H.; Laurie, R.E.; Mysore, K.S.; Wen, J.; Weller, J.L.; Macknight, R.C. Medicago truncatula SOC1 Genes Are Up-regulated by Environmental Cues That Promote Flowering. Front. Plant Sci. 2018, 9, 496. [Google Scholar] [CrossRef]
- Iqbal, M.; Erskine, W.; Berger, J.; Nelson, M. Phenotypic characterisation and linkage mapping of domestication syndrome traits in yellow lupin (Lupinus luteus L.). Theor. Appl. Genet. 2020, 133, 2975–2987. [Google Scholar] [CrossRef]
- Rychel, S.; Ksiazkiewicz, M.; Tomaszewska, M.; Bielski, W.; Wolko, B. FLOWERING LOCUS T, GIGANTEA, SEPALLATA and FRIGIDA homologs are candidate genes involved in white lupin (Lupinus albus L.) early flowering. Mol. Breed. 2019, 39, 43. [Google Scholar] [CrossRef]
- Rychel-Bielska, S.; Surma, A.; Bielski, W.; Kozak, B.; Galek, R.; Książkiewicz, M. Quantitative Control of Early Flowering in White Lupin (Lupinus albus L.). Int. J. Mol. Sci. 2021, 22, 3856. [Google Scholar] [CrossRef]
- Hecht, V.; Laurie, R.E.; Schoor, J.K.V.; Ridge, S.; Knowles, C.L.; Liew, L.C.; Sussmilch, F.C.; Murfet, I.C.; Macknight, R.C.; Weller, J.L. The Pea GIGAS Gene Is a FLOWERING LOCUS T Homolog Necessary for Graft-Transmissible Specification of Flowering but Not for Responsiveness to Photoperiod. Plant Cell 2011, 23, 147–161. [Google Scholar] [CrossRef]
- Ortega, R.; Hecht, V.F.G.; Freeman, J.S.; Rubio, J.; Carrasquilla-Garcia, N.; Mir, R.R.; Penmetsa, R.V.; Cook, D.R.; Millan, T.; Weller, J.L. Altered Expression of an FT Cluster Underlies a Major Locus Controlling Domestication-Related Changes to Chickpea Phenology and Growth Habit. Front. Plant Sci. 2019, 10, 824. [Google Scholar] [CrossRef]
- Rajandran, V.; Ortega, R.; Vander Schoor, J.K.; Butler, J.B.; Freeman, J.S.; Hecht, V.; Erskine, W.; Murfet, I.C.; Bett, K.E.; Weller, J.L. Genetic analysis of early phenology in lentil identifies distinct loci controlling component traits. Journal of experimental botany. J. Exp. Bot. 2022, 73, 3963–3977. [Google Scholar] [CrossRef]
- Yuan, H.; Caron, C.; Ramsay, L.; Fratini, R.; de la Vega, M.; Vandenberg, A.; Weller, J.; Bett, K. Genetic and gene expression analysis of flowering time regulation by light quality in lentil. Ann. Bot. 2021, 4, 469–480. [Google Scholar] [CrossRef]
- Yuan, X.; Wang, Q.; Yan, B.; Zhang, J.; Xue, C.; Chen, J.; Lin, Y.; Zhang, X.; Shen, W.; Chen, X. Single-Molecule Real-Time and Illumina-Based RNA Sequencing Data Identified Vernalization-Responsive Candidate Genes in Faba Bean (Vicia faba L.). Front. Genet. 2021, 12, 656137. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Benitez, D.; Casimiro-Soriguer, I.; Maalouf, F.; Torres, A.M. Linkage mapping and QTL analysis of flowering time in faba bean. Sci. Rep. 2021, 11, 13716. [Google Scholar] [CrossRef]
- Mateos, J.L.; Madrigal, P.; Tsuda, K.; Rawat, V.; Richter, R.; Romera-Branchat, M.; Fornara, F.; Schneeberger, K.; Krajewski, P.; Coupland, G. Combinatorial activities of SHORT VEGETATIVE PHASE and FLOWERING LOCUS C define distinct modes of flowering regulation in Arabidopsis. Genome Biol. 2015, 16, 31. [Google Scholar] [CrossRef]
- Castillejo, C.; Pelaz, S. The balance between CONSTANS and TEMPRANILLO activities determines FT expression to trigger flowering. Curr. Biol. 2008, 18, 1338–1343. [Google Scholar] [CrossRef] [Green Version]
- Osnato, M.; Castillejo, C.; Matías-Hernández, L.; Pelaz, S. TEMPRANILLO genes link photoperiod and gibberellin pathways to control flowering in Arabidopsis. Nat. Commun. 2012, 3, 808. [Google Scholar] [CrossRef]
- Schwarz, S.; Grande, A.V.; Bujdoso, N.; Saedler, H.; Huijser, P. The microRNA regulated SBP-box genes SPL9 and SPL15 control shoot maturation in Arabidopsis. Plant Mol. Biol. 2008, 67, 183–195. [Google Scholar] [CrossRef]
- Hyun, Y.; Richter, R.; Vincent, C.; Martinez-Gallegos, R.; Porri, A.; Coupland, G. Multi-layered Regulation of SPL15 and Cooperation with SOC1 Integrate Endogenous Flowering Pathways at the Arabidopsis Shoot Meristem. Dev. Cell 2016, 37, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Madrid, E.; Chandler, J.W.; Coupland, G. Gene regulatory networks controlled by FLOWERING LOCUS C that confer variation in seasonal flowering and life history. J. Exp. Bot. 2021, 72, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, R.J.; Amasino, R.M. Vernalization: A model for investigating epigenetics and eukaryotic gene regulation in plants. Biochim. Biophys. Acta 2007, 1769, 269–275. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, D.; He, Y. FRIGIDA establishes a local chromosomal environment for FLOWERING LOCUS C mRNA production. Nat. Plants 2018, 4, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, C.C.; Rouse, D.T.; Finnegan, E.J.; Peacock, W.J.; Dennis, E.S. The molecular basis of vernalization: The central role of FLOWERING LOCUS C (FLC). Proc. Natl Acad. Sci. USA 2000, 97, 3753–3758. [Google Scholar] [CrossRef] [PubMed]
- Gendall, A.R.; Levy, Y.Y.; Wilson, A.; Dean, C. The VERNALIZATION 2 gene mediates the epigenetic regulation of vernalization in Arabidopsis. Cell 2001, 107, 525–535. [Google Scholar] [CrossRef]
- Bastow, R.; Mylne, J.S.; Lister, C.; Lippman, Z.; Martienssen, R.A.; Dean, C. Vernalization requires epigenetic silencing of FLC by histone methylation. Nature 2004, 427, 164–167. [Google Scholar] [CrossRef]
- Sung, S.; Amasino, R.M. Vernalization and epigenetics: How plants remember winter. Curr. Opin. Plant Biol. 2004, 7, 4–10. [Google Scholar] [CrossRef]
- Sung, S.; Amasino, R.M. Remembering winter: Toward a molecular understanding of vernalization. Annu. Rev. Plant Biol. 2005, 56, 491–508. [Google Scholar] [CrossRef]
- Sung, S.; Schmitz, R.J.; Amasino, R.M. A PHD finger protein involved in both the vernalization and photoperiod pathways in Arabidopsis. Genes Dev. 2006, 20, 3244–3248. [Google Scholar] [CrossRef]
- Wood, C.; Robertson, M.; Tanner, G.; Peacock, W.; Dennis, E.; Helliwell, C. The Arabidopsis thaliana vernalization response requires a polycomb-like protein complex that also includes VERNALIZATION INSENSITIVE 3. Proc. Natl Acad. Sci. USA 2006, 103, 14631–14636. [Google Scholar] [CrossRef]
- Greb, T.; Mylne, J.; Crevillen, P.; Geraldo, N.; An, H.; Gendall, A.; Dean, C. The PHD finger protein VRN5 functions in the epigenetic silencing of Arabidopsis FLC. Curr. Biol. 2007, 17, 73–78. [Google Scholar] [CrossRef]
- De Lucia, F.; Crevillen, P.; Jones, A.; Greb, T.; Dean, C. A PHD-polycomb repressive complex 2 triggers the epigenetic silencing of FLC during vernalization. Proc. Natl Acad. Sci. USA 2008, 105, 16831–16836. [Google Scholar] [CrossRef]
- Berry, S.; Dean, C. Environmental perception and epigenetic memory: Mechanistic insight through FLC. Plant J. Cell Mol. Biol. 2015, 83, 133–148. [Google Scholar] [CrossRef]
- Whittaker, C.; Dean, C. The FLC Locus: A Platform for Discoveries in Epigenetics and Adaptation. Annual review of cell and developmental biology. Annu. Rev. Cell Dev. Biol. 2017, 33, 555–575. [Google Scholar] [CrossRef]
- Costa, S.; Dean, C. Storing memories: The distinct phases of Polycomb-mediated silencing of Arabidopsis FLC. Biochem. Soc. Trans. 2019, 47, 1187–1196. [Google Scholar] [CrossRef]
- Golembeski, G.S.; Imaizumi, T. Photoperiodic Regulation of Florigen Function in Arabidopsis thaliana. Arab. Book 2015, 13, e0178. [Google Scholar] [CrossRef]
- Golembeski, G.S.; Kinmonth-Schultz, A.H.; Song, Y.H.; Imaizumi, T. Photoperiodic flowering regulation in Arabidopsis thaliana. Adv. Bot. Res. 2014, 72, 1–28. [Google Scholar] [CrossRef] [Green Version]
- An, H.; Roussot, C.; Suárez-López, P.; Corbesier, L.; Vincent, C.; Piñeiro, M.; Hepworth, S.; Mouradov, A.; Justin, S.; Turnbull, C.; et al. CONSTANS acts in the phloem to regulate a systemic signal that induces photoperiodic flowering of Arabidopsis. Development 2004, 131, 3615–3626. [Google Scholar] [CrossRef]
- Song, Y.H.; Lee, I.; Lee, S.Y.; Imaizumi, T.; Hong, J.C. CONSTANS and ASYMMETRIC LEAVES 1 complex is involved in the induction of FLOWERING LOCUS T in photoperiodic flowering in Arabidopsis. Plant J. Cell Mol. Biol. 2012, 69, 332–342. [Google Scholar] [CrossRef]
- Platten, J.D.; Foo, E.; Foucher, F.; Hecht, V.; Reid, J.B.; Weller, J.L. The cryptochrome gene family in pea includes two differentially expressed CRY2 genes. Plant Mol. Biol. 2005, 59, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Pierre, J.B.; Bogard, M.; Herrmann, D.; Huyghe, C.; Julier, B. A CONSTANS-like gene candidate that could explain most of the genetic variation for flowering date in Medicago truncatula. Mol. Breed. 2011, 28, 25–35. [Google Scholar] [CrossRef]
- Berbel, A.; Ferrándiz, C.; Hecht, V.; Dalmais, M.; Lund, O.S.; Sussmilch, F.C.; Taylor, S.A.; Bendahmane, A.; Ellis, T.H.; Beltrán, J.P.; et al. VEGETATIVE1 is essential for development of the compound inflorescence in pea. Nat. Commun. 2012, 3, 797. [Google Scholar] [CrossRef]
- Kim, M.; Kang, Y.; Lee, T.; Lee, S. Divergence of flowering-related genes in three legume species. Plant Genome 2013, 6, 4. [Google Scholar] [CrossRef]
- Chao, Y.; Yang, Q.; Kang, J.; Zhang, T.; Sun, Y. Expression of the alfalfa FRIGIDA-like gene, MsFRI-L delays flowering time in transgenic Arabidopsis thaliana. Mol. Biol. Rep. 2013, 40, 2083–2090. [Google Scholar] [CrossRef] [PubMed]
- Jaudal, M.; Monash, J.; Zhang, L.; Wen, J.; Mysore, K.S.; Macknight, R.; Putterill, J. Overexpression of Medicago SVP genes causes floral defects and delayed flowering in Arabidopsis but only affects floral development in Medicago. J. Exp. Bot. 2014, 65, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Sussmilch, F.C.; Berbel, A.; Hecht, V.; Schoor, J.K.V.; Ferrándiz, C.; Madueño, F.; Weller, J.L. Pea VEGETATIVE2 Is an FD Homolog That Is Essential for Flowering and Compound Inflorescence Development. Plant Cell 2015, 27, 1046–1060. [Google Scholar] [CrossRef]
- Ksiazkiewicz, M.; Rychel, S.; Nelson, M.; Wyrwa, K.; Naganowska, B.; Wolko, B. Expansion of the phosphatidyleth-anolamine binding protein family in legumes: A case study of Lupinus angustifolius L. FLOWERING LOCUS T homologs, LanFTc1 and LanFTc2. BMC Genom. 2016, 17, 820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridge, S.; Deokar, A.; Lee, R.; Daba, K.; Macknight, R.C.; Weller, J.L.; Tar’an, B. The Chickpea Early Flowering 1 (Efl1) Locus Is an Ortholog of Arabidopsis ELF3. Plant Physiol. 2017, 175, pp.00082.2017. [Google Scholar] [CrossRef]
- Jaudal, M.; Zhang, L.; Che, C.; Li, G.; Tang, Y.; Wen, J.; Mysore, K.S.; Putterill, J. A SOC1-like gene MtSOC1a promotes flowering and primary stem elongation in Medicago. J. Exp. Bot. 2018, 69, 4867–4880. [Google Scholar] [CrossRef]
- Cheng, X.; Li, G.; Tang, Y.; Wen, J. Dissection of genetic regulation of compound inflorescence development in Medicago truncatula. Development 2018, 145, dev.158766. [Google Scholar] [CrossRef] [PubMed]
- Jaudal, M.; Wen, J.; Mysore, K.S.; Putterill, J. Medicago PHYA promotes flowering, primary stem elongation and expression of flowering time genes in long days. BMC Plant Biol. 2020, 20, 329. [Google Scholar] [CrossRef]
- Thomson, G.; Zhang, L.; Wen, J.; Mysore, K.S.; Putterill, J. The Candidate Photoperiod Gene MtFE Promotes Growth and Flowering in Medicago truncatula. Front. Plant Sci. 2021, 12, 634091. [Google Scholar] [CrossRef]
- Cheng, X.; Li, G.; Krom, N.; Tang, Y.; Wen, J. Genetic regulation of flowering time and inflorescence architecture by MtFDa and MtFTa1 in Medicago truncatula. Plant Physiol. 2021, 185, 161–178. [Google Scholar] [CrossRef]
- Williams, O.; Vander Schoor, J.K.; Butler, J.B.; Ridge, S.; Sussmilch, F.C.; Hecht, V.; Weller, J.L. The genetic architecture of flowering time changes in pea from wild to crop. J. Exp. Bot. 2022, 73, 3978–3990. [Google Scholar] [CrossRef]
- Watanabe, S.; Harada, K.; Abe, J. Genetic and molecular bases of photoperiod responses of flowering in soybean. Breed. Sci. 2012, 61, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Lyu, J.; Cai, Z.; Li, Y.; Suo, H.; Yi, R.; Zhang, S.; Nian, H. The Floral Repressor GmFLC-like Is Involved in Regulating Flowering Time Mediated by Low Temperature in Soybean. Int. J. Mol. Sci. 2020, 21, 1322. [Google Scholar] [CrossRef]
- Weller, J.L.; Macknight, R.C. Functional Genomics and Flowering Time in Medicago truncatula: An Overview. Methods Mol. Biol. 2018, 1822, 261–271. [Google Scholar] [CrossRef]
- Kroc, M.; Koczyk, G.; Święcicki, W.; Kilian, A.; Nelson, M.N. New evidence of ancestral polyploidy in the Genistoid legume Lupinus angustifolius L. (narrow-leafed lupin). Theor. Appl. Genet. 2014, 127, 1237–1249. [Google Scholar] [CrossRef]
- Ferrándiz, C.; Gu, Q.; Martienssen, R.; Yanofsky, M.F. Redundant regulation of meristem identity and plant architecture by FRUITFULL, APETALA1 and CAULIFLOWER. Development 2000, 127, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Kobayashi, Y.; Yamamoto, S.; Daimon, Y.; Yamaguchi, A.; Ikeda, Y.; Ichinoki, H.; Notaguchi, M.; Goto, K.; Araki, T. FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science 2005, 309, 1052–1056. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, H.; Nakamura, H.; Taoka, K.; Shimamoto, K. Functional diversification of FD transcription factors in rice, components of florigen activation complexes. Plant Cell Physiol. 2013, 54, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhai, H.; Wang, Y.; Tian, X.; Zhang, Y.; Wu, H.; Lü, S.; Yang, G.; Li, Y.; Wang, L.; et al. Functional conservation and diversification of the soybean maturity gene E1 and its homologs in legumes. Sci. Rep. 2016, 6, srep29548. [Google Scholar] [CrossRef]
- Collani, S.; Neumann, M.; Yant, L.; Schmid, M. FT Modulates Genome-Wide DNA-Binding of the bZIP Transcription Factor FD. Plant Physiol. 2019, 180, 367–380. [Google Scholar] [CrossRef]
- Romera-Branchat, M.; Severing, E.; Pocard, C.; Ohr, H.; Vincent, C.; Née, G.; Martinez-Gallegos, R.; Jang, S.; Andrés, F.; Madrigal, P.; et al. Functional Divergence of the Arabidopsis Florigen-Interacting bZIP Transcription Factors FD and FDP. Cell Rep. 2020, 32, 107966. [Google Scholar] [CrossRef]
- Teper-Bamnolker, P.; Samach, A. The flowering integrator FT regulates SEPALLATA3 and FRUITFULL accumulation in Arabidopsis leaves. Plant Cell 2005, 17, 2661–2675. [Google Scholar] [CrossRef]
- Benlloch, R.; Berbel, A.; Ali, L.; Gohari, G.; Millán, T.; Madueño, F. Genetic control of inflorescence architecture in legumes. Front. Plant Sci. 2015, 6, 543. [Google Scholar] [CrossRef]
- Taylor, C.; Kamphuis, L.; Zhang, W.; Garg, G.; Berger, J.; Mousavi-Derazmahalleh, M.; Bayer, P.; Edwards, D.; Singh, K.; Cowling, W.; et al. INDEL variation in the regulatory region of the major flowering time gene LanFTc1 is associated with vernalization response and flowering time in narrow-leafed lupin (Lupinus angustifolius L.). Plant Cell Environ. 2019, 42, 174–187. [Google Scholar] [CrossRef] [Green Version]
- Mikołajczyk, J. Genetic studies in Lupinus angustifolius. Part. III. Inheritance of the alkaloid content, seed hardness and length of the growing season in blue lupin. Front. Plant Sci. 1966, 7, 181–196. [Google Scholar]
- Gladstones, J.S.; Hill, G.D. Selection for economic characters in Lupinus angustifolius and L. digitatus. 2. Time of flowering. Aust. J. Exp. Agric. Anim. Husb. 1969, 9, 213–220. [Google Scholar] [CrossRef]
- Liu, L.; Adrian, J.; Pankin, A.; Hu, J.; Dong, X.; von Korff, M.; Turck, F. Induced and natural variation of promoter length modulates the photoperiodic response of FLOWERING LOCUS T. Nat. Commun. 2014, 5, 4558. [Google Scholar] [CrossRef]
- Plewiński, P.; Książkiewicz, M.; Rychel-Bielska, S.; Rudy, E.; Wolko, B.F. Candidate Domestication-Related Genes Revealed by Expression Quantitative Trait Loci Mapping of Narrow-Leafed Lupin (Lupinus angustifolius L.). Int. J. Mol. Sci. 2019, 20, 5670. [Google Scholar] [CrossRef]
- Mandel, M.A.; Yanofsky, M.F. The Arabidopsis AGL8 MADS box gene is expressed in inflorescence meristems and is negatively regulated by APETALA1. Plant Cell 1995, 7, 1763–1771. [Google Scholar] [CrossRef]
- Hempel, F.D.; Weigel, D.; Mandel, M.A.; Ditta, G.; Zambryski, P.C.; Feldman, L.J.; Yanofsky, M.F. Floral determination and expression of floral regulatory genes in Arabidopsis. Development 1997, 124, 3845–3853. [Google Scholar] [CrossRef]
- Gu, Q.; Ferrandiz, C.; Yanofsky, M.F.; Martienssen, R. The FRUITFULL MADS-box gene mediates cell differentiation during Arabidopsis fruit development. Development 1998, 125, 1509–1517. [Google Scholar] [CrossRef]
- Cowling, W.; Huyghe, C.; Swiecicki, W.; Gladstones, J.; Atkins, C.; Hamblin, J. Lupin Breeding; CAB International: London, UK, 1998; pp. 93–120. [Google Scholar]
- Nelson, M.; Berger, J.; Erskine, W. Flowering time control in annual legumes: Prospects in a changing global climate. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2012, 5, 1–14. [Google Scholar] [CrossRef]
- Lichtin, N.; Salvo-Garrido, H.; Till, B.; Caligari, P.; Rupayan, A.; Westermeyer, F.; Olivos, M. Genetic and comparative mapping of Lupinus luteus L. highlight syntenic regions with major orthologous genes controlling anthracnose resistance and flowering time. Sci. Rep. 2020, 10, 19174. [Google Scholar] [CrossRef]
- Książkiewicz, M.; Nazzicari, N.; Yang, H.; Nelson, M.N.; Renshaw, D.; Rychel, S.; Ferrari, B.; Carelli, M.; Tomaszewska, M.; Stawiński, S.; et al. A high-density consensus linkage map of white lupin highlights synteny with narrow-leafed lupin and provides markers tagging key agronomic traits. Sci. Rep. 2017, 7, 15335. [Google Scholar] [CrossRef]
- Johanson, U.; West, J.; Lister, C.; Michaels, S.; Amasino, R.; Dean, C. Molecular analysis of FRIGIDA, a major determinant of natural variation in Arabidopsis flowering time. Science 2005, 290, 344–347. [Google Scholar] [CrossRef]
- Shindo, C.; Aranzana, M.J.; Lister, C.; Baxter, C.; Nicholls, C.; Nordborg, M.; Dean, C. Role of FRIGIDA and FLOWERING LOCUS C in determining variation in flowering time of Arabidopsis. Plant Physiol. 2005, 138, 1163–1173. [Google Scholar] [CrossRef] [PubMed]
- Beveridge, C.; Murfet, I. The gigas mutant in pea is deficient in the floral stimulus. Physiol. Plant 1996, 96, 637–645. [Google Scholar] [CrossRef]
- Reid, J.; Murfet, I. Flowering in Pisum: The sites and possible mechanisms of the vernalization response. J. Exp. Bot. 1975, 26, 860–867. [Google Scholar] [CrossRef]
- Samineni, S.; Kamatam, S.; Thudi, M.; Varshney, R.; Gaur, P. Vernalization response in chickpea is controlled by a major QTL. Euphytica 2016, 207, 453–461. [Google Scholar] [CrossRef]
- Cobos, M.; Winter, P.; Kharrat, M.; Cubero, J.; Gil, J.; Milian, T.; Rubio, J. Genetic analysis of agronomic traits in a wide cross of chickpea. Field Crops Res. 2009, 111, 130–136. [Google Scholar] [CrossRef]
- Aryamanesh, N.; Nelson, M.; Yan, G.; Clarke, H.; Siddique, K. Mapping a major gene for growth habit and QTLs for ascochyta blight resistance and flowering time in a population between chickpea and Cicer reticulatum. Euphytica 2010, 173, 307–319. [Google Scholar] [CrossRef]
- Hossain, S.; Ford, R.; McNeil, D.; Pittock, C.; Panozzo, J. Development of a selection tool for seed shape and QTL analysis of seed shape with other morphological traits for selective breeding in chickpea (Cicer arietinum L.). Aust. J. Crop Sci. 2010, 4, 278–288. [Google Scholar]
- Mallikarjuna, B.P.; Samineni, S.; Thudi, M.; Sajja, S.B.; Khan, A.W.; Patil, A.; Viswanatha, K.P.; Varshney, R.K.; Gaur, P.M. Molecular Mapping of Flowering Time Major Genes and QTLs in Chickpea (Cicer arietinum L.). Front. Plant Sci. 2017, 8, 1140. [Google Scholar] [CrossRef]
- Srivastava, R.; Upadhyaya, H.; Kumar, R.; Daware, A.; Basu, U.; Shimray, P.; Tripathi, S.; Bharadwaj, C.; Tyagi, A.; Parida, S. A multiple QTL-Seq strategy delineates potential genomic loci governing flowering time in chickpea. Front. Plant Sci. 2017, 8, 1105. [Google Scholar] [CrossRef]
- Weller, J.L.; Vander Schoor, J.K.; Perez-Wright, E.C.; Hecht, V.; Gonzalez, A.M.; Capel, C.; Yuste-Lisbona, F.J.; Lozano, R.; Santalla, M. Parallel origins of photoperiod adaptation following dual domestications of common bean. J. Exp. Bot. 2019, 70, 1209–1219. [Google Scholar] [CrossRef]
- Kim, D.H.; Sung, S. Coordination of the vernalization response through a VIN3 and FLC gene family regulatory network in Arabidopsis. Plant Cell 2013, 25, 454–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaudal, M.; Zhang, L.; Che, C.; Hurley, D.G.; Thomson, G.; Wen, J.; Mysore, K.S.; Putterill, J. MtVRN2 is a Polycomb VRN2-like gene which represses the transition to flowering in the model legume Medicago truncatula. Plant J. Cell Mol. Biol. 2016, 86, 145–160. [Google Scholar] [CrossRef]
- Yan, L.; Loukoianov, A.; Blechl, A.; Tranquilli, G.; Ramakrishna, W.; SanMiguel, P.; Bennetzen, J.; Echenique, V.; Dubcovsky, J. The wheat VRN2 gene is a flowering repressor downregulated by vernalization. Science 2004, 303, 1640–1644. [Google Scholar] [CrossRef] [PubMed]
- Hemming, M.N.; Peacock, W.J.; Dennis, E.S.; Trevaskis, B. Low-temperature and daylength cues are integrated to regulate FLOWERING LOCUS T in barley. Plant Physiol. 2008, 147, 355–366. [Google Scholar] [CrossRef]
- Pin, P.; Benlloch, R.; Bonnet, D.; Wremerth-Weich, E.; Kraft, T.; Gielen, J.; Nilsson, O. An antagonistic pair of FT homologs mediates the control of flowering time in sugar beet. Science 2010, 330, 1397–1400. [Google Scholar] [CrossRef]
- Foucher, F.; Morin, J.; Courtiade, J.; Cadioux, S.; Ellis, N.; Banfield, M.J.; Rameau, C. DETERMINATE and LATE FLOWERING are two TERMINAL FLOWER1/CENTRORADIALIS homologs that control two distinct phases of flowering initiation and development in pea. Plant Cell 2003, 15, 2742–2754. [Google Scholar] [CrossRef]
- Gursky, V.V.; Kozlov, K.N.; Nuzhdin, S.V.; Samsonova, M.G. Dynamical Modeling of the Core Gene Network Controlling Flowering Suggests Cumulative Activation From the FLOWERING LOCUS T Gene Homologs in Chickpea. Front. Genet. 2018, 9, 547. [Google Scholar] [CrossRef] [Green Version]
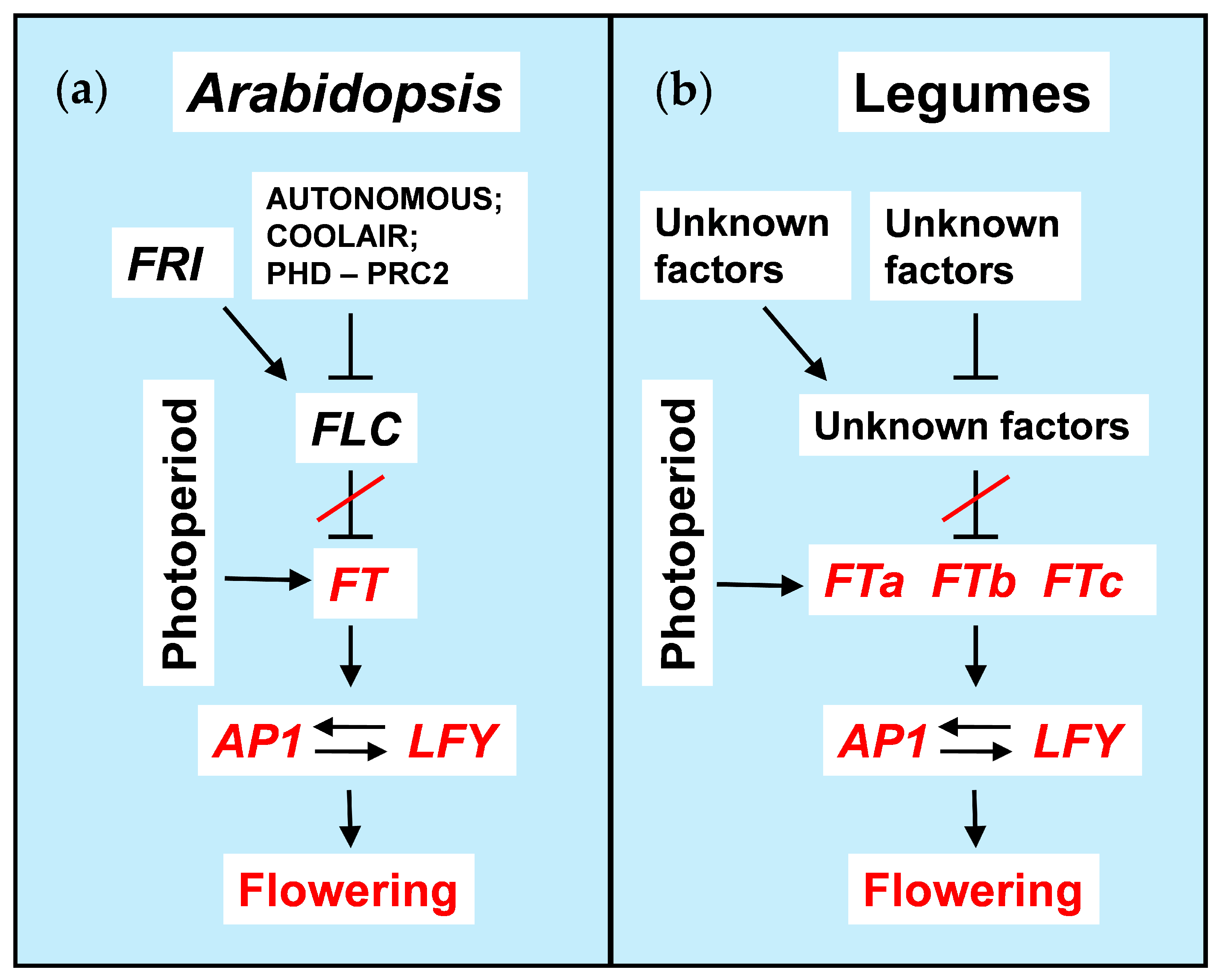
Figure 1.
Flowering promotion by vernalization. (a) A generalized regulatory mechanism of cold-induced flowering promotion in Arabidopsis, (b) a putative regulatory network in legumes. Arrows and T-bars show positive and negative regulatory interactions respectively. Activation state of the core components is shown with the red color. ‘Autonomous’ and ‘Photoperiod’ denote the corresponding pathways in A. thaliana. FTa, FTb and FTc are three subclades of FT orthologs in legumes.
Figure 1.
Flowering promotion by vernalization. (a) A generalized regulatory mechanism of cold-induced flowering promotion in Arabidopsis, (b) a putative regulatory network in legumes. Arrows and T-bars show positive and negative regulatory interactions respectively. Activation state of the core components is shown with the red color. ‘Autonomous’ and ‘Photoperiod’ denote the corresponding pathways in A. thaliana. FTa, FTb and FTc are three subclades of FT orthologs in legumes.


{kind=link}
Table 1.
The orthologs of Arabidopsis FLOWERING LOCUS T (FT) gene in legumes.
| Legume Species | FT Genes |
|---|---|
| Medicago trancatula | FTa1, FTa2, FTa3, FTb1, FTb2, FTc |
| Pisum sativum | FTa1, FTa2, FTb1, FTb2, FTc |
| Lens culinaris | FTa1, FTa2, FTa3, FTb1, FTb2, FTc |
| Cicer arietinum | FTa1, FTa2, FTa3, FTb, FTc |
| Lupinus angustifolius | FTa1, FTa2, FTc1, FTc2 |
Table 2.
Major vernalization targets (FT and SOC1 genes) in different legume species.
| Legume Species | Targets of Vernalization | References |
|---|---|---|
| Medicago trancatula | FTa1, SOC1 | [44,48] |
| [46,49] | ||
| Lupinus angustifolius | FTc1 | [45] |
| Lupinus luteus | FTc1 | [50] |
| Lupinus albus | polygenic regulation, | [51] |
| including FTa1 and FTc1 | [52] | |
| Pisum sativum | FTa1 (?) | [53] |
| Cicer arietinum | FTa1, FTa2, FTc (?) | [54] |
| Lens culinaris | FTa1, FTa2 | [55,56] |
| Vicia faba | FT, SOC1 | [57] |
| [58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Surkova, S.Y.; Samsonova, M.G. Mechanisms of Vernalization-Induced Flowering in Legumes. Int. J. Mol. Sci. 2022, 23, 9889. https://doi.org/10.3390/ijms23179889
AMA Style
Surkova SY, Samsonova MG. Mechanisms of Vernalization-Induced Flowering in Legumes. International Journal of Molecular Sciences. 2022; 23(17):9889. https://doi.org/10.3390/ijms23179889
Chicago/Turabian StyleSurkova, Svetlana Yu., and Maria G. Samsonova. 2022. "Mechanisms of Vernalization-Induced Flowering in Legumes" International Journal of Molecular Sciences 23, no. 17: 9889. https://doi.org/10.3390/ijms23179889
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.