
Novel Potency Assay for MSC Secretome-Based Treatment of Idiopathic Male Infertility Employed Leydig Cells and Revealed Vascular Endothelial Growth Factor as a Promising Potency Marker

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
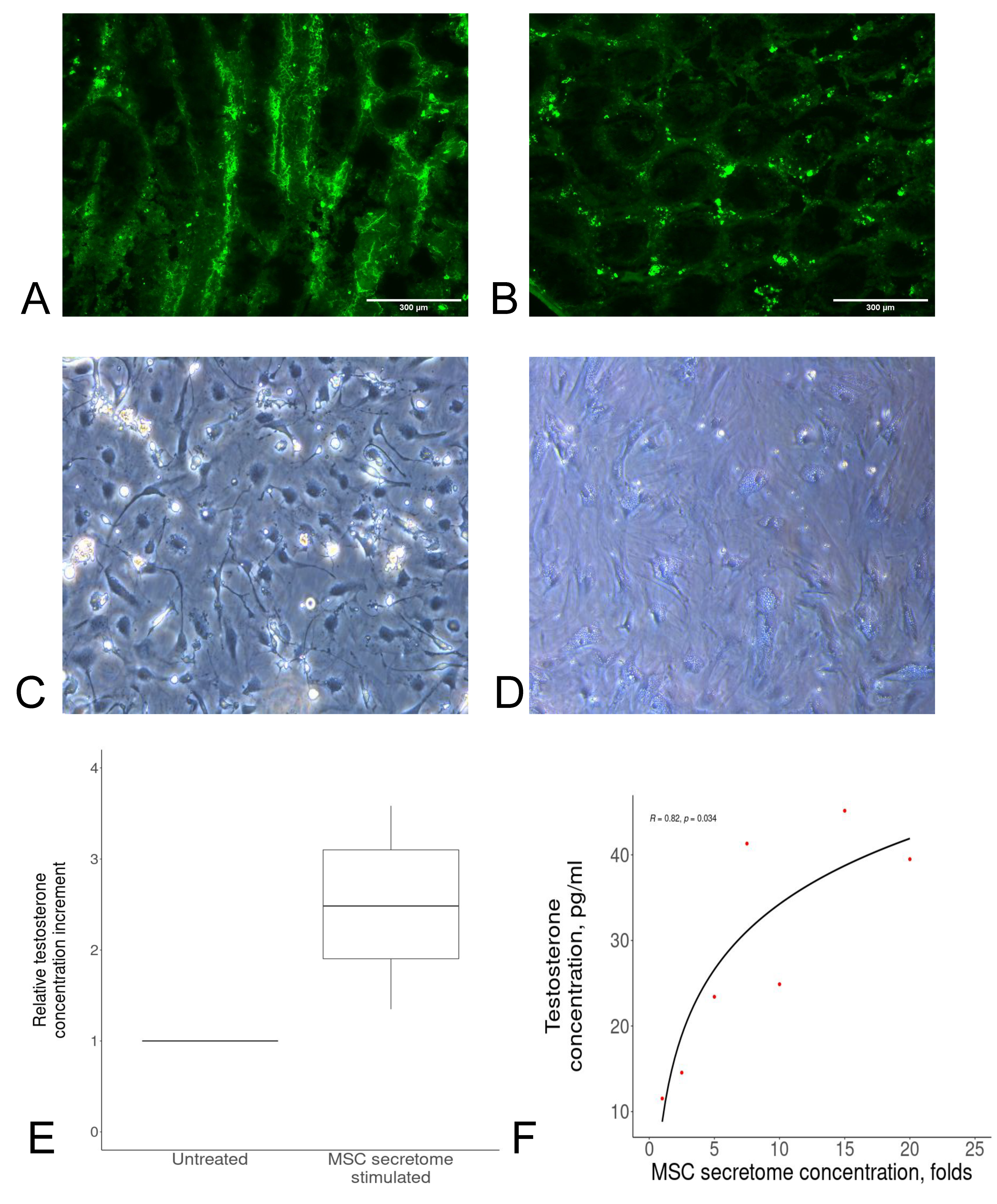
2.1. MSC Secretome Stimulates Testosterone Secretion by Leydig Cells in a Dose-Dependent Manner
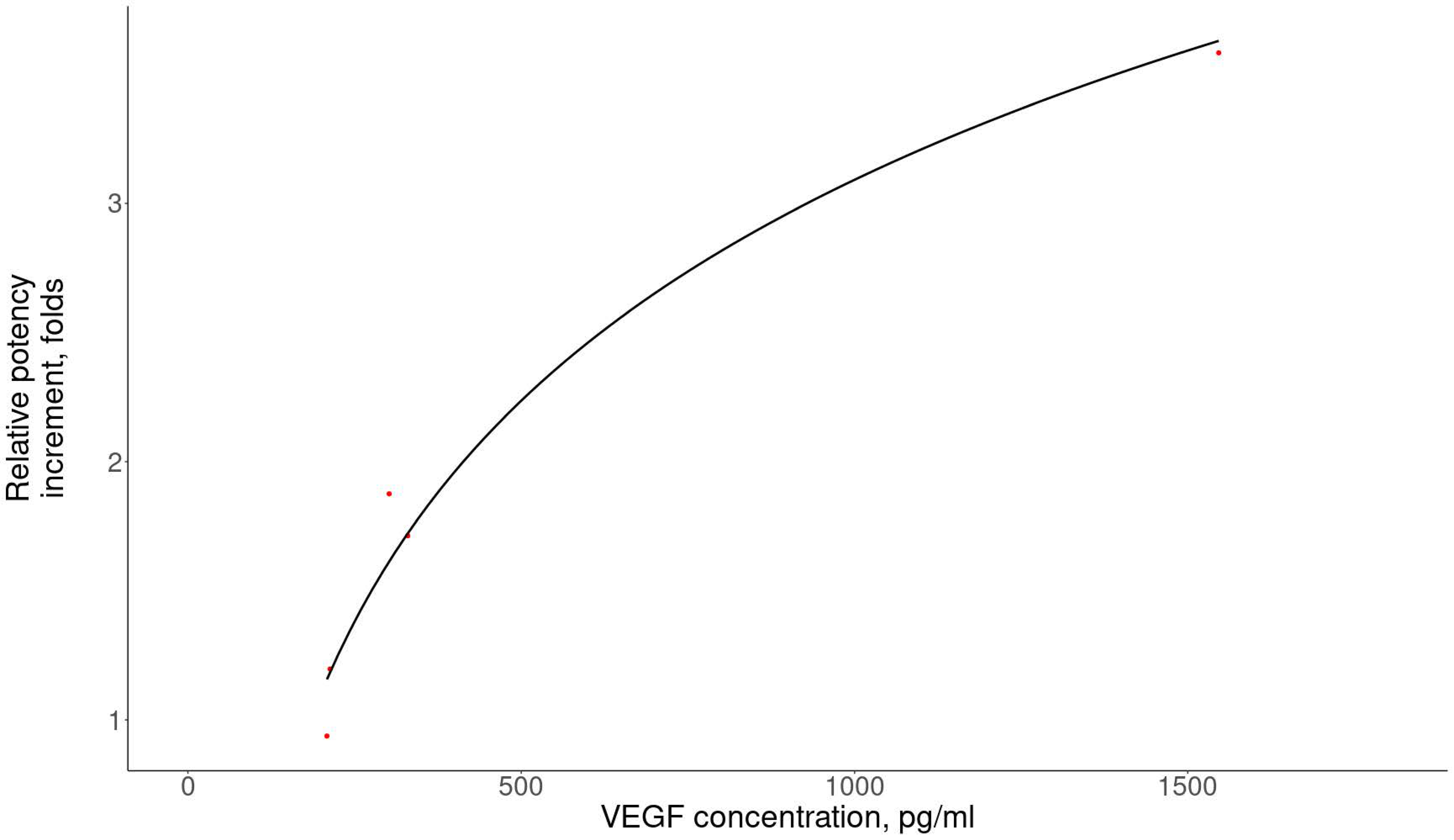
2.2. Testosterone Secretion by Leydig Cells Is Directly Related to VEGF Content in MSC Secretome
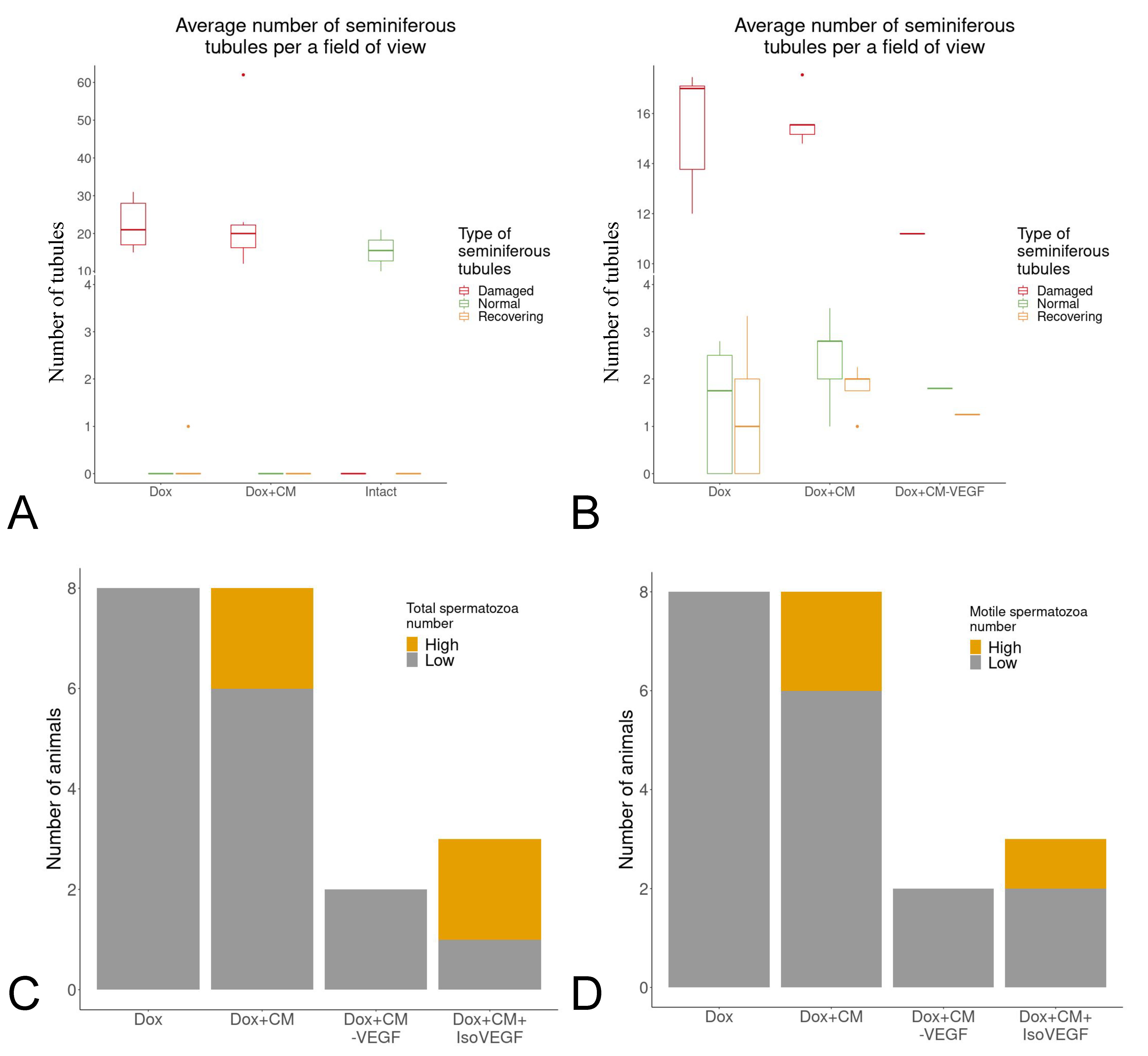
2.3. Removal of VEGF Abolishes Effects of MSC Secretome Injection in a Murine Model of Doxorubicin-Induced Male Infertility
3. Discussion
4. Materials and Methods
4.1. Isolation of MSCs
4.2. Manufacturing of MSC Secretome
4.3. Isolation of Leydig Cells
4.4. Modeling the Potency Assay using Leydig Cells
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Doxorubicin-Induced Male Infertility Murine Model
4.6.1. Chemicals
4.6.2. Animals
4.7. Histological Analysis
4.8. Morphometric Analysis of the Contents of the Epididymis
4.9. Phase-Contrast Microscopy
4.10. Biodistribution Assay
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agarwal, A.; Baskaran, S.; Parekh, N.; Cho, C.L.; Henkel, R.; Vij, S.; Arafa, M.; Panner Selvam, M.K.; Shah, R. Male Infertility. Lancet 2021, 397, 319–333. [Google Scholar] [CrossRef]
- Shahid, M.N.; Khan, T.M.; Neoh, C.F.; Lean, Q.Y.; Bukhsh, A.; Karuppannan, M. Effectiveness of Pharmacological Intervention Among Men with Infertility: A Systematic Review and Network Meta-Analysis. Front. Pharmacol. 2021, 12, 638628. [Google Scholar] [CrossRef] [PubMed]
- Cannarella, R.; Condorelli, R.A.; Mongioì, L.M.; Barbagallo, F.; Calogero, A.E.; La Vignera, S. Effects of the Selective Estrogen Receptor Modulators for the Treatment of Male Infertility: A Systematic Review and Meta-Analysis. Expert Opin. Pharmacother. 2019, 20, 1517–1525. [Google Scholar] [CrossRef] [PubMed]
- Steiner, A.Z.; Hansen, K.R.; Barnhart, K.T.; Cedars, M.I.; Legro, R.S.; Diamond, M.P.; Krawetz, S.A.; Usadi, R.; Baker, V.L.; Coward, R.M.; et al. The Effect of Antioxidants on Male Factor Infertility: The Males, Antioxidants, and Infertility (MOXI) Randomized Clinical Trial. Fertil. Steril. 2020, 113, 552–560.e3. [Google Scholar] [CrossRef] [PubMed]
- Sagaradze, G.; Monakova, A.; Basalova, N.; Popov, V.; Balabanyan, V.; Efimenko, A. Regenerative Medicine for Male Infertility: A Focus on Stem Cell Niche Injury Models. Biomed. J. 2022, 45, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhankina, R.; Baghban, N.; Askarov, M.; Saipiyeva, D.; Ibragimov, A.; Kadirova, B.; Khoradmehr, A.; Nabipour, I.; Shirazi, R.; Zhanbyrbekuly, U.; et al. Mesenchymal Stromal/stem Cells and Their Exosomes for Restoration of Spermatogenesis in Non-Obstructive Azoospermia: A Systemic Review. Stem Cell Res. Ther. 2021, 12, 229. [Google Scholar] [CrossRef]
- Gauthier-Fisher, A.; Kauffman, A.; Librach, C.L. Potential Use of Stem Cells for Fertility Preservation. Andrology 2020, 8, 862–878. [Google Scholar] [CrossRef] [Green Version]
- Sagaradze, G.D.; Basalova, N.A.; Kirpatovsky, V.I.; Ohobotov, D.A.; Grigorieva, O.A.; Balabanyan, V.Y.; Kamalov, A.A.; Efimenko, A.Y. Application of Rat Cryptorchidism Model for the Evaluation of Mesenchymal Stromal Cell Secretome Regenerative Potential. Biomed. Pharmacother. 2019, 109, 1428–1436. [Google Scholar] [CrossRef]
- Mohammed, S.S.; Mansour, M.F.; Salem, N.A. Therapeutic Effect of Stem Cells on Male Infertility in a Rat Model: Histological, Molecular, Biochemical, and Functional Study. Stem Cells Int. 2021, 2021, 8450721. [Google Scholar] [CrossRef]
- Bateman, M.E.; Strong, A.L.; Gimble, J.M.; Bunnell, B.A. Concise Review: Using Fat to Fight Disease: A Systematic Review of Nonhomologous Adipose-Derived Stromal/Stem Cell Therapies. Stem Cells 2018, 36, 1311–1328. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, W.-Z.; Lin, Y.-H.; Su, L.-J.; Wu, M.-S.; Jeng, H.-Y.; Chang, H.-C.; Huang, Y.-H.; Ling, T.-Y. Mesenchymal Stem/stromal Cell-Based Therapy: Mechanism, Systemic Safety and Biodistribution for Precision Clinical Applications. J. Biomed. Sci. 2021, 28, 28. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, S.; Shi, Y.; Galipeau, J.; Krampera, M.; Leblanc, K.; Martin, I.; Nolta, J.; Phinney, D.G.; Sensebe, L. Mesenchymal Stem versus Stromal Cells: International Society for Cell & Gene Therapy (ISCT®) Mesenchymal Stromal Cell Committee Position Statement on Nomenclature. Cytotherapy 2019, 21, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Wuchter, P.; Bieback, K.; Schrezenmeier, H.; Bornhäuser, M.; Müller, L.P.; Bönig, H.; Wagner, W.; Meisel, R.; Pavel, P.; Tonn, T.; et al. Standardization of Good Manufacturing Practice-Compliant Production of Bone Marrow-Derived Human Mesenchymal Stromal Cells for Immunotherapeutic Applications. Cytotherapy 2015, 17, 128–139. [Google Scholar] [CrossRef]
- Sagaradze, G.D.; Basalova, N.A.; Efimenko, A.Y.; Tkachuk, V.A. Mesenchymal Stromal Cells as Critical Contributors to Tissue Regeneration. Front. Cell Dev. Biol. 2020, 8, 576176. [Google Scholar] [CrossRef]
- Merimi, M.; El-Majzoub, R.; Lagneaux, L.; Moussa Agha, D.; Bouhtit, F.; Meuleman, N.; Fahmi, H.; Lewalle, P.; Fayyad-Kazan, M.; Najar, M. The Therapeutic Potential of Mesenchymal Stromal Cells for Regenerative Medicine: Current Knowledge and Future Understandings. Front. Cell Dev. Biol. 2021, 9, 661532. [Google Scholar] [CrossRef] [PubMed]
- Bari, E.; Perteghella, S.; Catenacci, L.; Sorlini, M.; Croce, S.; Mantelli, M.; Avanzini, M.A.; Sorrenti, M.; Torre, M.L. Freeze-Dried and GMP-Compliant Pharmaceuticals Containing Exosomes for Acellular Mesenchymal Stromal Cell Immunomodulant Therapy. Nanomedicine 2019, 14, 753–765. [Google Scholar] [CrossRef]
- Gimona, M.; Brizzi, M.F.; Choo, A.B.H.; Dominici, M.; Davidson, S.M.; Grillari, J.; Hermann, D.M.; Hill, A.F.; de Kleijn, D.; Lai, R.C.; et al. Critical Considerations for the Development of Potency Tests for Therapeutic Applications of Mesenchymal Stromal Cell-Derived Small Extracellular Vesicles. Cytotherapy 2021, 23, 373–380. [Google Scholar] [CrossRef]
- de Wolf, C.; van de Bovenkamp, M.; Hoefnagel, M. Regulatory Perspective on in Vitro Potency Assays for Human Mesenchymal Stromal Cells Used in Immunotherapy. Cytotherapy 2017, 19, 784–797. [Google Scholar] [CrossRef]
- Thej, C.; Ramadasse, B.; Walvekar, A.; Majumdar, A.S.; Balasubramanian, S. Development of a Surrogate Potency Assay to Determine the Angiogenic Activity of Stempeucel®, a Pooled, Ex-Vivo Expanded, Allogeneic Human Bone Marrow Mesenchymal Stromal Cell Product. Stem Cell Res. Ther. 2017, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- European Medicines Agency. ICH Topic Q 6 B Specifications: Test Procedures and Acceptance Criteria for Biotechnological/Biological Products; European Medicines Agency: Amsterdam, The Netherlands, 1999. [CrossRef]
- Hwang, G.S.; Wang, S.W.; Tseng, W.M.; Yu, C.H.; Wang, P.S. Effect of Hypoxia on the Release of Vascular Endothelial Growth Factor and Testosterone in Mouse TM3 Leydig Cells. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E1763–E1769. [Google Scholar] [CrossRef]
- Sagaradze, G.; Basalova, N.; Kirpatovsky, V.; Ohobotov, D.; Nimiritsky, P.; Grigorieva, O.; Popov, V.; Kamalov, A.; Tkachuk, V.; Efimenko, A. A Magic Kick for Regeneration: Role of Mesenchymal Stromal Cell Secretome in Spermatogonial Stem Cell Niche Recovery. Stem Cell Res. Ther. 2019, 10, 342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhushan, S.; Aslani, F.; Zhang, Z.; Sebastian, T.; Elsässer, H.-P.; Klug, J. Isolation of Sertoli Cells and Peritubular Cells from Rat Testes. J. Vis. Exp. 2016, 108, e53389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punab, M.; Poolamets, O.; Paju, P.; Vihljajev, V.; Pomm, K.; Ladva, R.; Korrovits, P.; Laan, M. Causes of Male Infertility: A 9-Year Prospective Monocentre Study on 1737 Patients with Reduced Total Sperm Counts. Hum. Reprod. 2017, 32, 18–31. [Google Scholar] [CrossRef]
- Li, K.-P.; Yang, X.-S.; Wu, T. The Effect of Antioxidants on Sperm Quality Parameters and Pregnancy Rates for Idiopathic Male Infertility: A Network Meta-Analysis of Randomized Controlled Trials. Front. Endocrinol. 2022, 151, 810242. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, M.; Dobashi, M.; Yamasaki, T.; Kanzaki, M.; Okada, H.; Arakawa, S.; Kamidono, S. Significance of Serum Inhibin B Concentration for Evaluating Improvement in Spermatogenesis after Varicocelectomy. Hum. Reprod. 2001, 16, 1945–1949. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.A.; Wallace, E.M.; Groome, N.P.; Bellis, A.J.; Wu, F.C. Physiological Relationships between Inhibin B, Follicle Stimulating Hormone Secretion and Spermatogenesis in Normal Men and Response to Gonadotrophin Suppression by Exogenous Testosterone. Hum. Reprod. 1997, 12, 746–751. [Google Scholar] [CrossRef]
- Wu, N.; Murono, E.P. A Sertoli Cell-Secreted Paracrine Factor(s) Stimulates Proliferation and Inhibits Steroidogenesis of Rat Leydig Cells. Mol. Cell. Endocrinol. 1994, 106, 99–109. [Google Scholar] [CrossRef]
- Kregel, K.C.; Zhang, H.J. An Integrated View of Oxidative Stress in Aging: Basic Mechanisms, Functional Effects, and Pathological Considerations. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R18–R36. [Google Scholar] [CrossRef]
- Caires, K.C.; de Avila, J.; McLean, D.J. Vascular Endothelial Growth Factor Regulates Germ Cell Survival during Establishment of Spermatogenesis in the Bovine Testis. Reproduction 2009, 138, 667–677. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, H.; Kats, R.; Danilovich, N.; Javeshghani, D.; Sairam, M.R. Intercellular Communication between Sertoli Cells and Leydig Cells in the Absence of Follicle-Stimulating Hormone-Receptor Signaling. Biol. Reprod. 2001, 65, 1201–1207. [Google Scholar] [CrossRef] [Green Version]
- Bhang, D.H.; Kim, B.-J.; Kim, B.G.; Schadler, K.; Baek, K.-H.; Kim, Y.H.; Hsiao, W.; Ding, B.-S.; Rafii, S.; Weiss, M.J.; et al. Testicular Endothelial Cells Are a Critical Population in the Germline Stem Cell Niche. Nat. Commun. 2018, 9, 4379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Windschüttl, S.; Kampfer, C.; Mayer, C.; Flenkenthaler, F.; Fröhlich, T.; Schwarzer, J.U.; Köhn, F.M.; Urbanski, H.; Arnold, G.J.; Mayerhofer, A. Human Testicular Peritubular Cells Secrete Pigment Epithelium-Derived Factor (PEDF), Which May Be Responsible for the Avascularity of the Seminiferous Tubules. Sci. Rep. 2015, 5, 12820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porubska, B.; Vasek, D.; Somova, V.; Hajkova, M.; Hlaviznova, M.; Tlapakova, T.; Holan, V.; Krulova, M. Sertoli Cells Possess Immunomodulatory Properties and the Ability of Mitochondrial Transfer Similar to Mesenchymal Stromal Cells. Stem Cell Rev. Rep. 2021, 17, 1905–1916. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.; Zhang, C.; Li, T.; Zhang, J.; Zhang, N.; Tao, Z.; Zhu, W.; Sun, X. Are Sertoli Cells a Kind of Mesenchymal Stem Cells? Am. J. Transl. Res. 2017, 9, 1067–1074. [Google Scholar]
- Galipeau, J.; Krampera, M.; Barrett, J.; Dazzi, F.; Deans, R.J.; DeBruijn, J.; Dominici, M.; Fibbe, W.E.; Gee, A.P.; Gimble, J.M.; et al. International Society for Cellular Therapy Perspective on Immune Functional Assays for Mesenchymal Stromal Cells as Potency Release Criterion for Advanced Phase Clinical Trials. Cytotherapy 2016, 18, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, L.; Tatum, M.E. Temporal Appearance of Seasonal Changes in Numbers of Sertoli Cells, Leydig Cells, and Germ Cells in Stallions. Biol. Reprod. 1989, 40, 994–999. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.; Lee, J.; Kwon, Y.; Park, K.-S.; Jeong, J.-H.; Choi, S.-J.; Bang, S.I.; Chang, J.W.; Lee, C. Comparative Proteomic Analysis of the Mesenchymal Stem Cells Secretome from Adipose, Bone Marrow, Placenta and Wharton’s Jelly. Int. J. Mol. Sci. 2021, 22, 845. [Google Scholar] [CrossRef]
- Andrzejewska, A.; Catar, R.; Schoon, J.; Qazi, T.H.; Sass, F.A.; Jacobi, D.; Blankenstein, A.; Reinke, S.; Krüger, D.; Streitz, M.; et al. Multi-Parameter Analysis of Biobanked Human Bone Marrow Stromal Cells Shows Little Influence for Donor Age and Mild Comorbidities on Phenotypic and Functional Properties. Front. Immunol. 2019, 10, 2474. [Google Scholar] [CrossRef]
- Choudhery, M.S.; Badowski, M.; Muise, A.; Pierce, J.; Harris, D.T. Donor Age Negatively Impacts Adipose Tissue-Derived Mesenchymal Stem Cell Expansion and Differentiation. J. Transl. Med. 2014, 12, 8. [Google Scholar] [CrossRef] [Green Version]
- Kokai, L.E.; Traktuev, D.O.; Zhang, L.; Merfeld-Clauss, S.; DiBernardo, G.; Lu, H.; Marra, K.G.; Donnenberg, A.; Donnenberg, V.; Meyer, E.M.; et al. Adipose Stem Cell Function Maintained with Age: An Intra-Subject Study of Long-Term Cryopreserved Cells. Aesthet. Surg. J. 2017, 37, 454–463. [Google Scholar] [CrossRef] [Green Version]
- Mohamed-Ahmed, S.; Fristad, I.; Lie, S.A.; Suliman, S.; Mustafa, K.; Vindenes, H.; Idris, S.B. Adipose-Derived and Bone Marrow Mesenchymal Stem Cells: A Donor-Matched Comparison. Stem Cell Res. Ther. 2018, 9, 168. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-H.K.; Ogando, C.R.; Wang See, C.; Chang, T.-Y.; Barabino, G.A. Changes in Phenotype and Differentiation Potential of Human Mesenchymal Stem Cells Aging in Vitro. Stem Cell Res. Ther. 2018, 9, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagaradze, G.; Grigorieva, O.; Nimiritsky, P.; Basalova, N.; Kalinina, N.; Akopyan, Z.; Efimenko, A. Conditioned Medium from Human Mesenchymal Stromal Cells: Towards the Clinical Translation. Int. J. Mol. Sci. 2019, 20, 1656. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Liu, Y.; Sun, Y.; Wang, B.; Xiong, Y.; Lin, W.; Wei, Q.; Wang, H.; He, W.; Wang, B.; et al. Tissue Source Determines the Differentiation Potentials of Mesenchymal Stem Cells: A Comparative Study of Human Mesenchymal Stem Cells from Bone Marrow and Adipose Tissue. Stem Cell Res. Ther. 2017, 8, 275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Available online: https://imagej.net/contribute/citing (accessed on 1 August 2022).



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monakova, A.; Sagaradze, G.; Basalova, N.; Popov, V.; Balabanyan, V.; Efimenko, A. Novel Potency Assay for MSC Secretome-Based Treatment of Idiopathic Male Infertility Employed Leydig Cells and Revealed Vascular Endothelial Growth Factor as a Promising Potency Marker. Int. J. Mol. Sci. 2022, 23, 9414. https://doi.org/10.3390/ijms23169414
Monakova A, Sagaradze G, Basalova N, Popov V, Balabanyan V, Efimenko A. Novel Potency Assay for MSC Secretome-Based Treatment of Idiopathic Male Infertility Employed Leydig Cells and Revealed Vascular Endothelial Growth Factor as a Promising Potency Marker. International Journal of Molecular Sciences. 2022; 23(16):9414. https://doi.org/10.3390/ijms23169414
Chicago/Turabian StyleMonakova, Anna, Georgy Sagaradze, Nataliya Basalova, Vladimir Popov, Vadim Balabanyan, and Anastasia Efimenko. 2022. "Novel Potency Assay for MSC Secretome-Based Treatment of Idiopathic Male Infertility Employed Leydig Cells and Revealed Vascular Endothelial Growth Factor as a Promising Potency Marker" International Journal of Molecular Sciences 23, no. 16: 9414. https://doi.org/10.3390/ijms23169414