
Peripheral Serotonin Deficiency Affects Anxiety-like Behavior and the Molecular Response to an Acute Challenge in Rats

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
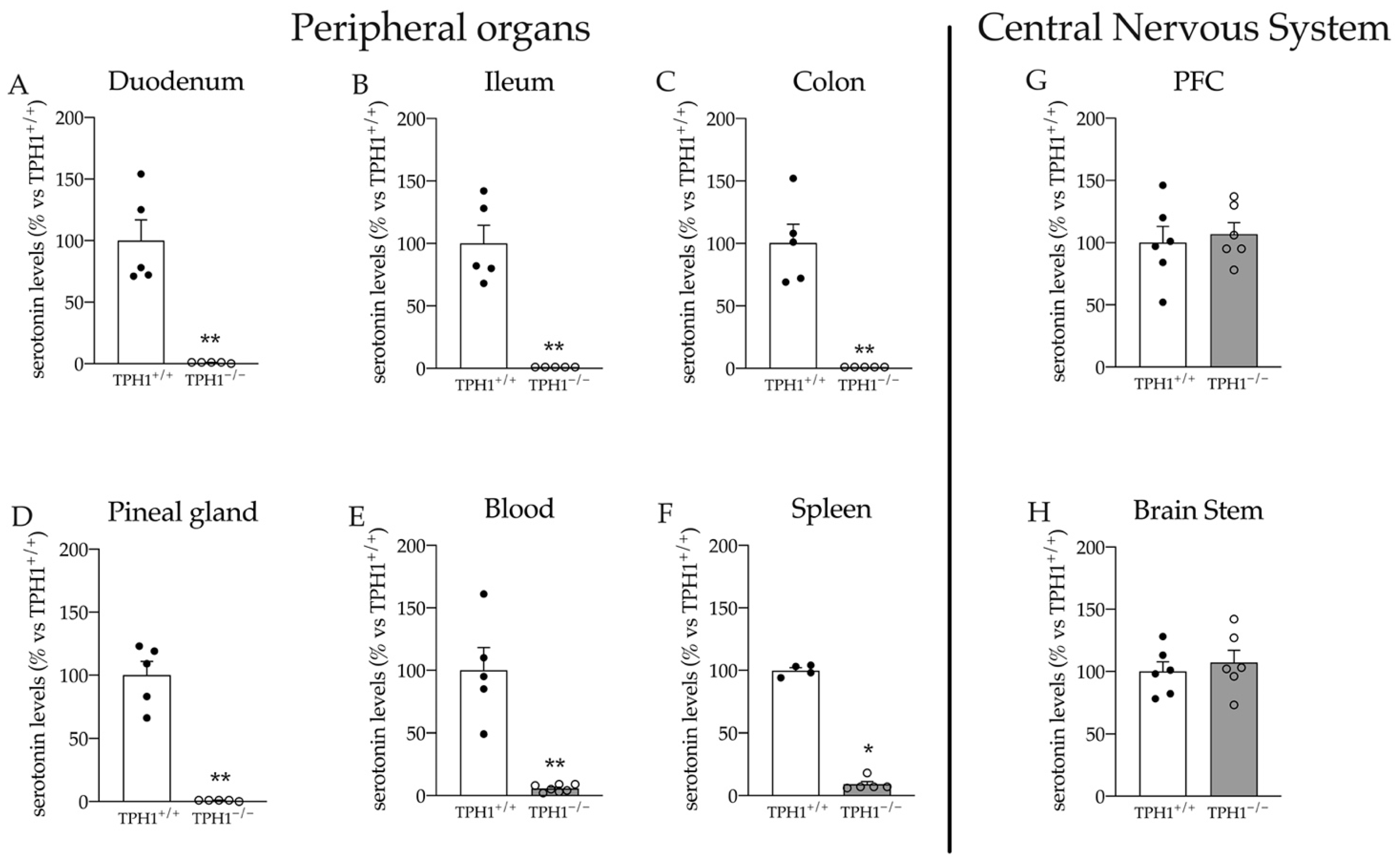
2.1. Serotonin Levels Are Dramatically Reduced in Peripheral Organs of TPH1−/− Rats
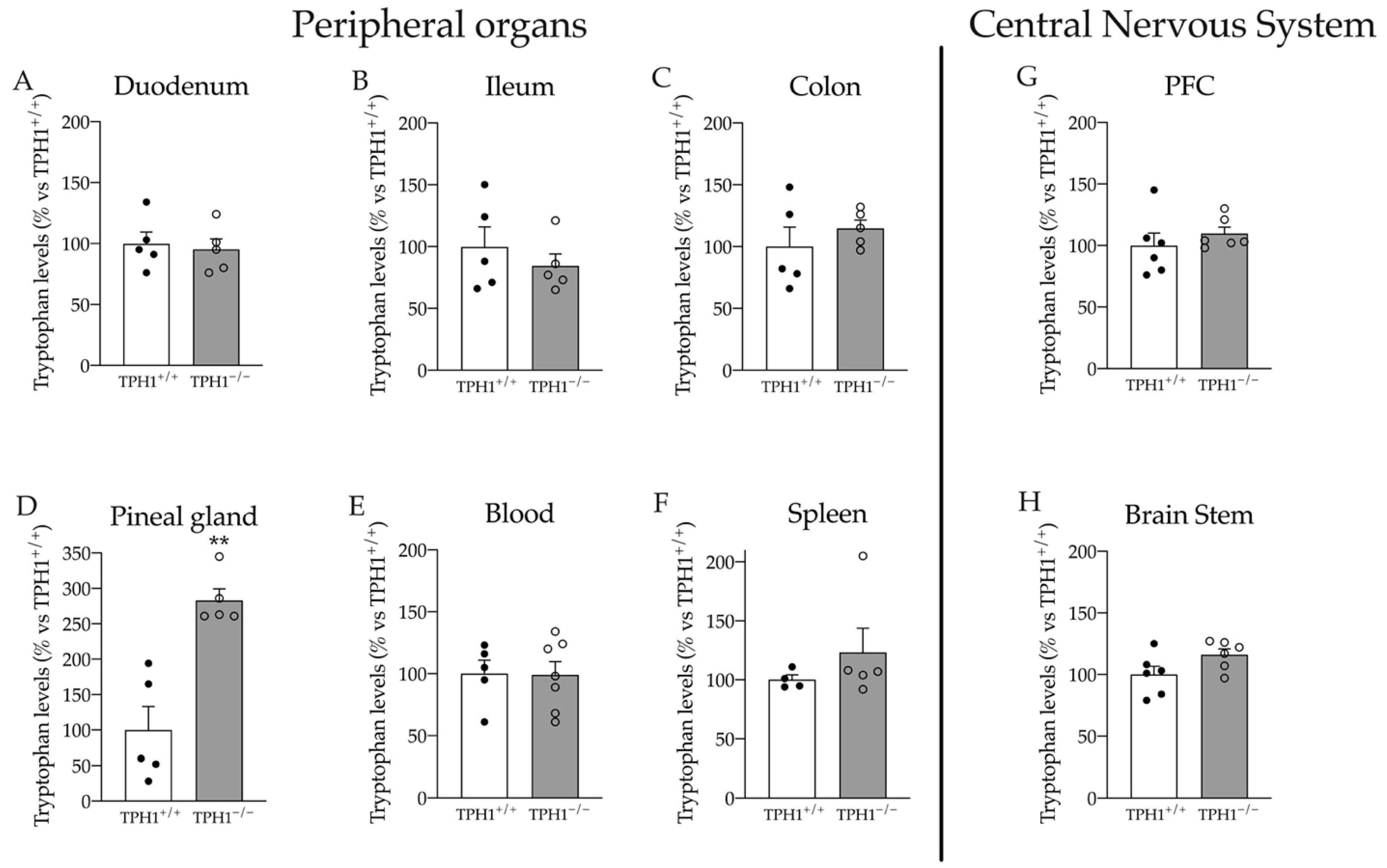
2.2. Tryptophan Levels Are Modulated in the Pineal Gland of TPH1−/− Rats
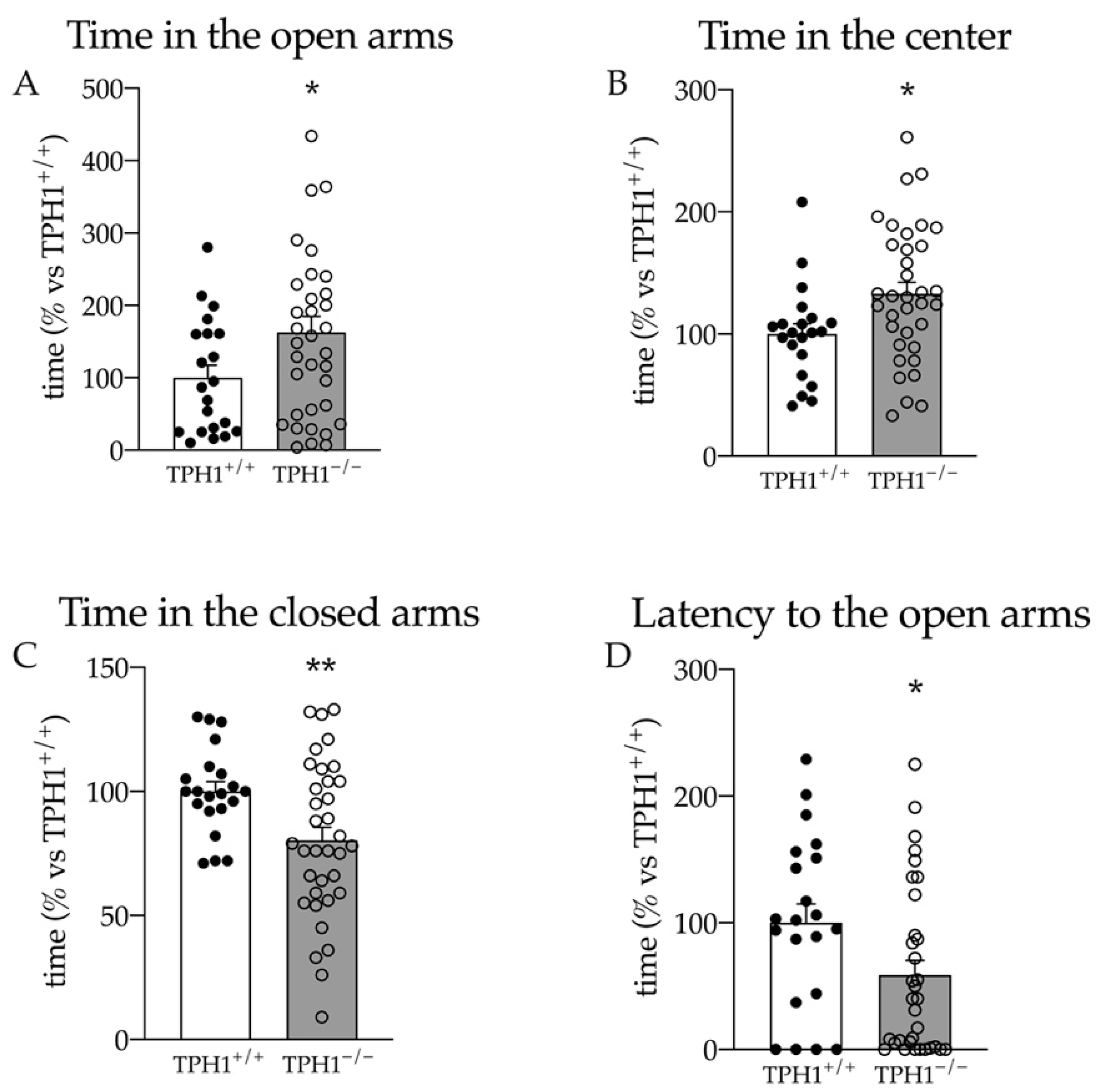
2.3. TPH1−/− Rats Show Reduced Anxiety in the Elevated Plus Maze Test
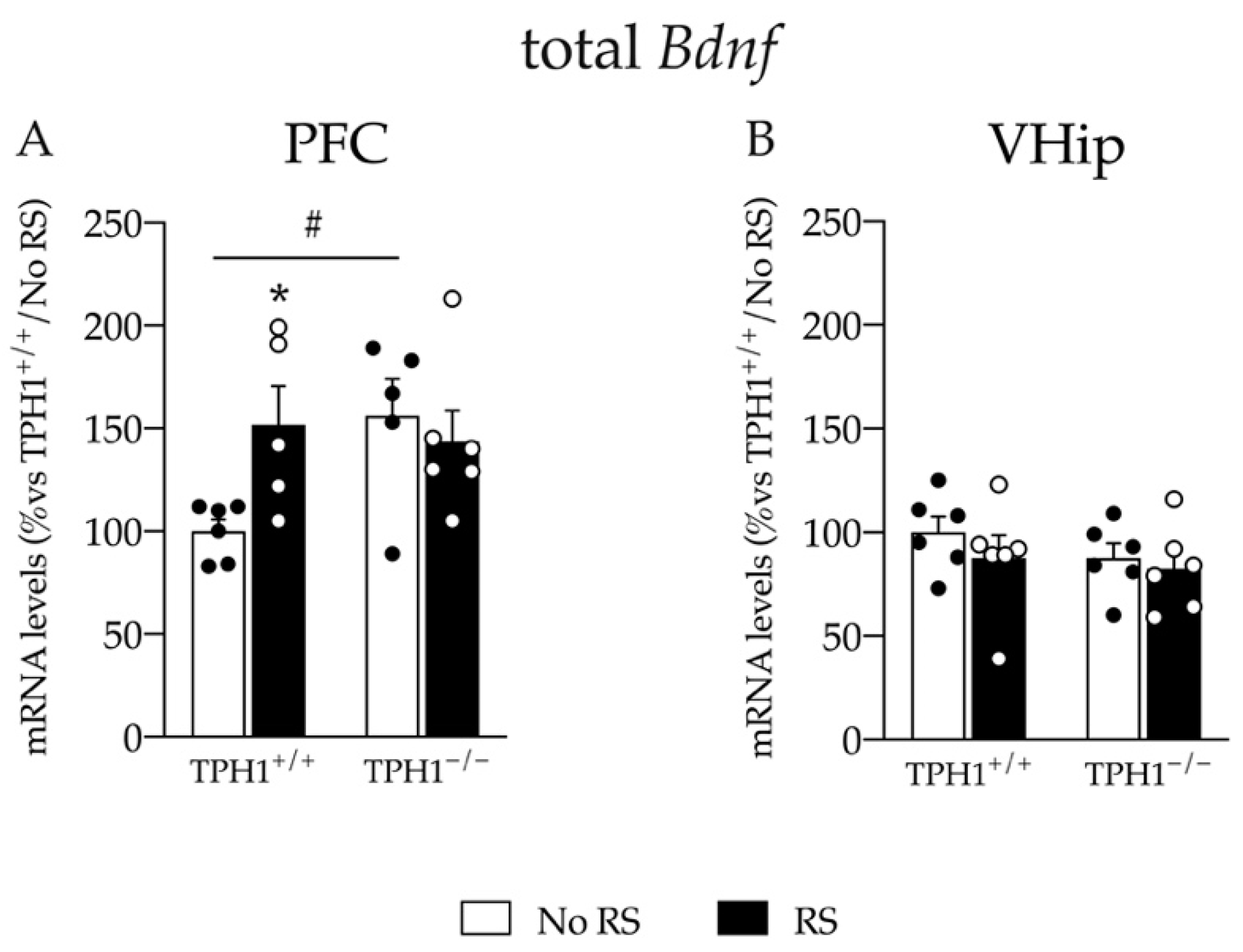
2.4. Baseline Bdnf mRNA Levels Are Increased and Induction of Its Expression Is Impaired after RS in TPH1−/− Rats Specifically in the Prefrontal Cortex
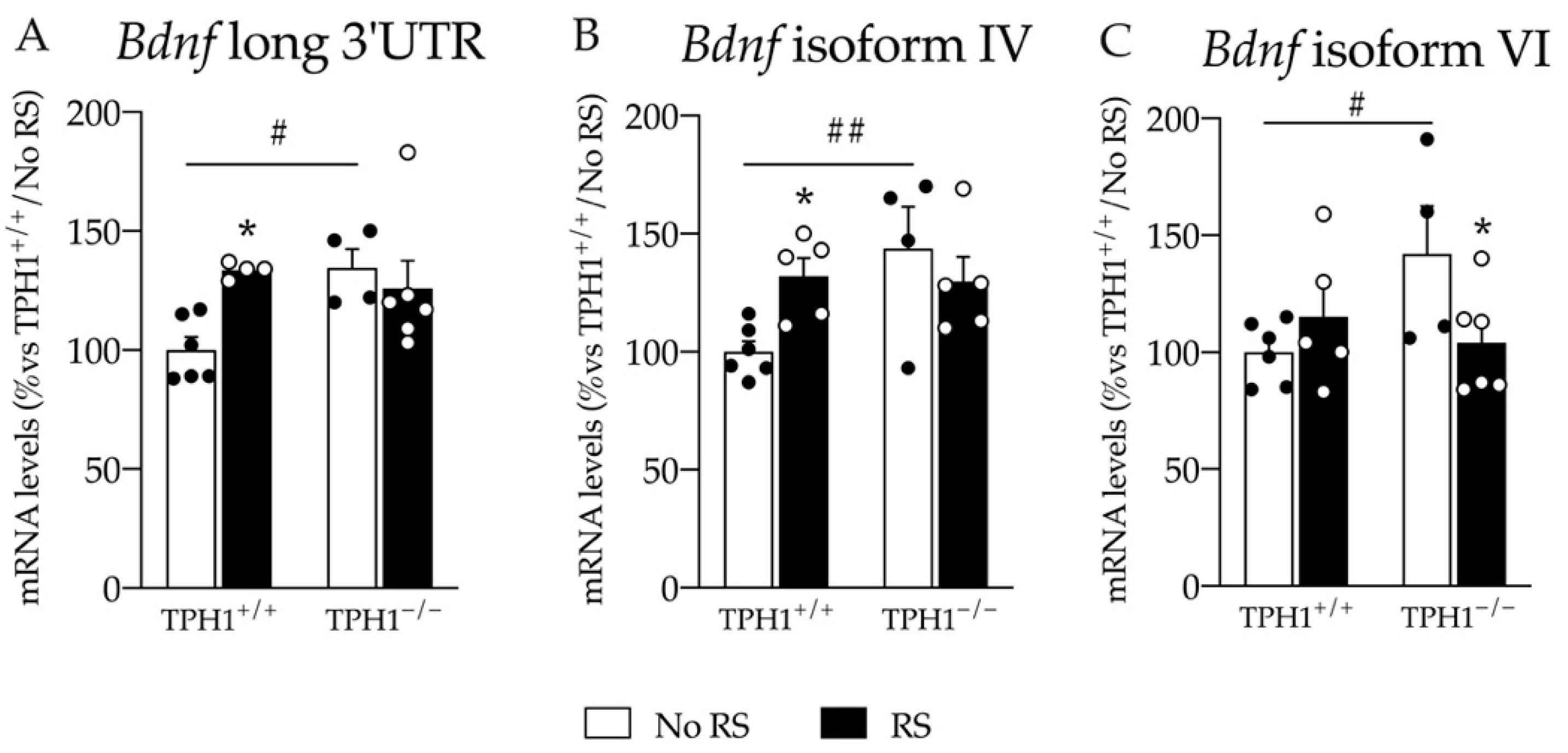
2.5. The Changes in Total Bdnf Expression in the PFC Are Sustained by Bdnf Long 3′UTR, Bdnf Isoform IV and VI
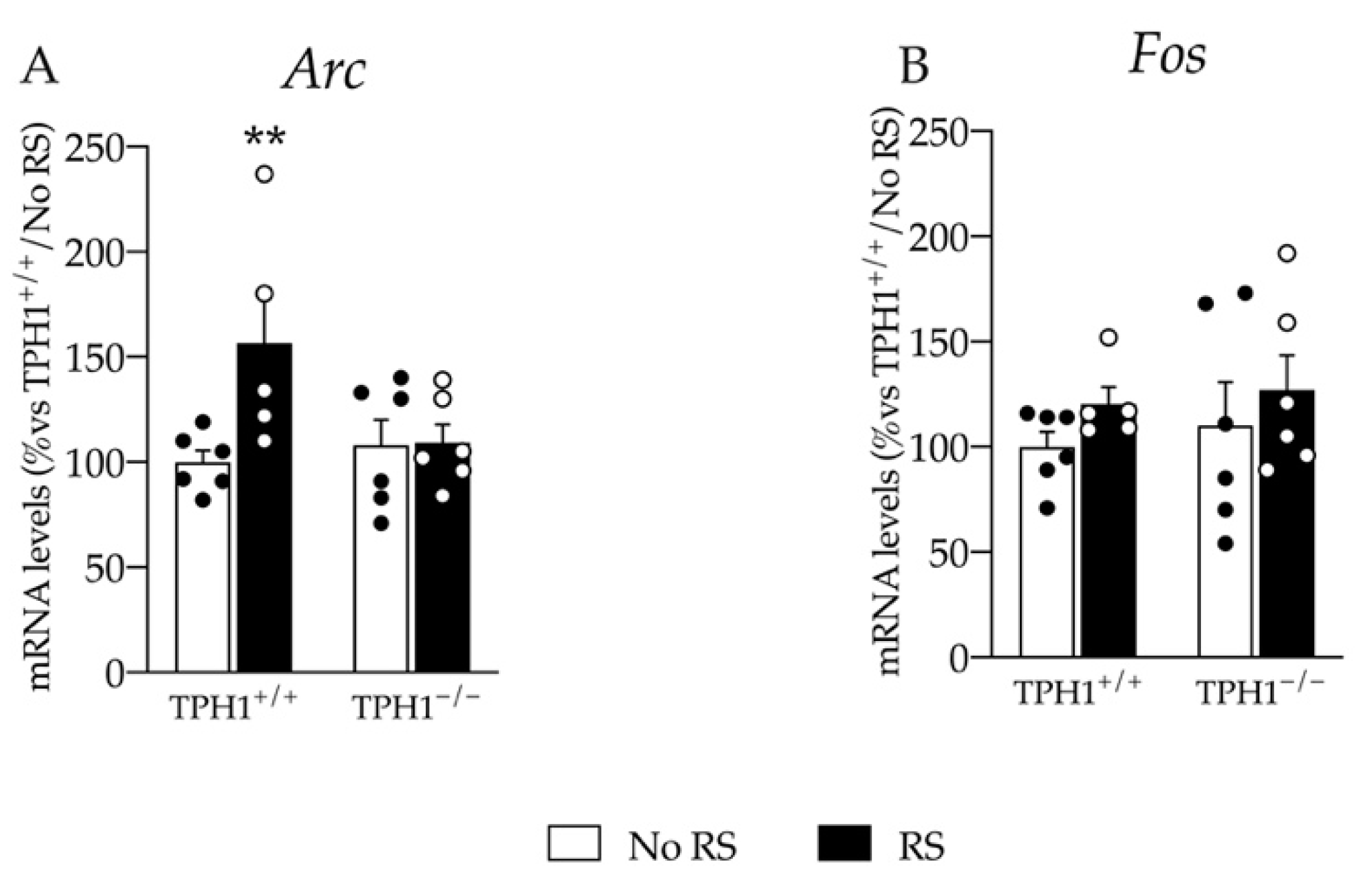
2.6. The Enhancement in the Immediate Early Gene Arc mRNA Levels Due to RS Exposure Is Blunted in TPH1−/− Rats
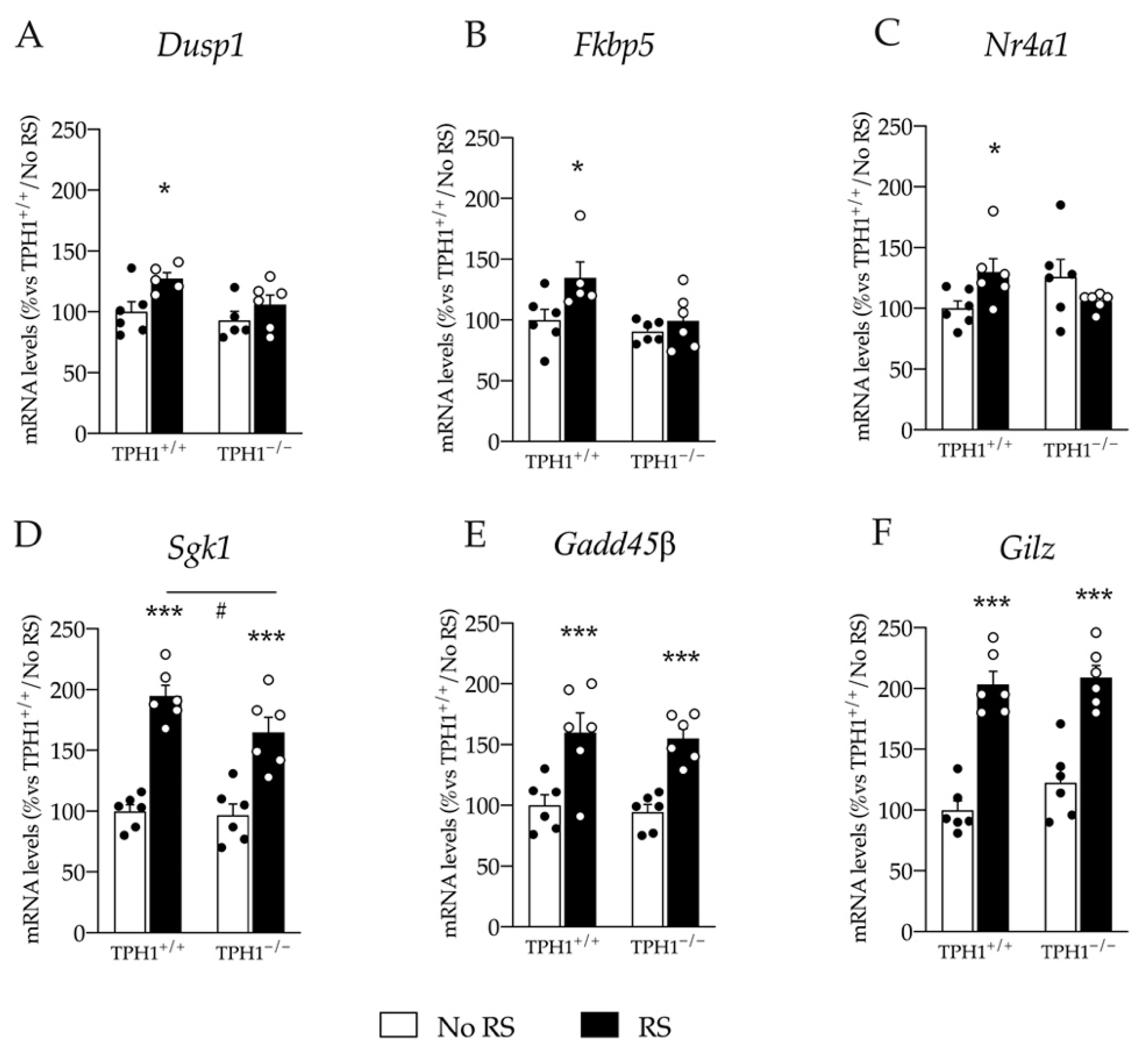
2.7. Upregulation of Glucocorticoid Responsive Genes Expression Induced by RS Exposure Is Blunted in TPH1−/− Rats
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. HPLC Analyses
4.3. mRNA Extraction and Gene Expression Analyses
4.4. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jenkins, T.; Nguyen, J.; Polglaze, K.; Bertrand, P. Influence of Tryptophan and Serotonin on Mood and Cognition with a Possible Role of the Gut-Brain Axis. Nutrients 2016, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, O.; Smith, J.R.; Purdy, R.E. Serotonin and vasoconstrictor synergism. Life Sci. 1998, 62, 1723–1732. [Google Scholar] [CrossRef]
- Paulmann, N.; Grohmann, M.; Voigt, J.-P.; Bert, B.; Vowinckel, J.; Bader, M.; Skelin, M.; Jevsek, M.; Fink, H.; Rupnik, M.; et al. Intracellular serotonin modulates insulin secretion from pancreatic beta-cells by protein serotonylation. PLoS Biol. 2009, 7, e1000229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Toyofuku, Y.; Lynn, F.C.; Chak, E.; Uchida, T.; Mizukami, H.; Fujitani, Y.; Kawamori, R.; Miyatsuka, T.; Kosaka, Y.; et al. Serotonin regulates pancreatic beta cell mass during pregnancy. Nat. Med. 2010, 16, 804–808. [Google Scholar] [CrossRef]
- El-Merahbi, R.; Löffler, M.; Mayer, A.; Sumara, G. The roles of peripheral serotonin in metabolic homeostasis. FEBS Lett. 2015, 589, 1728–1734. [Google Scholar] [CrossRef] [Green Version]
- Walther, D.J.; Bader, M. A unique central tryptophan hydroxylase isoform. Biochem. Pharmacol. 2003, 66, 1673–1680. [Google Scholar] [CrossRef]
- Walther, D.J.; Peter, J.U.; Bashammakh, S.; Hörtnagl, H.; Voits, M.; Fink, H.; Bader, M. Synthesis of serotonin by a second tryptophan hydroxylase isoform. Science 2003, 299, 76. [Google Scholar] [CrossRef]
- Zhang, X.; Beaulieu, J.-M.; Sotnikova, T.D.; Gainetdinov, R.R.; Caron, M.G. Tryptophan hydroxylase-2 controls brain serotonin synthesis. Science 2004, 305, 217. [Google Scholar] [CrossRef]
- Zill, P.; Baghai, T.C.; Zwanzger, P.; Schüle, C.; Eser, D.; Rupprecht, R.; Möller, H.-J.; Bondy, B.; Ackenheil, M. SNP and haplotype analysis of a novel tryptophan hydroxylase isoform (TPH2) gene provide evidence for association with major depression. Mol. Psychiatry 2004, 9, 1030–1036. [Google Scholar] [CrossRef] [Green Version]
- McKinney, J.; Knappskog, P.M.; Haavik, J. Different properties of the central and peripheral forms of human tryptophan hydroxylase. J. Neurochem. 2005, 92, 311–320. [Google Scholar] [CrossRef]
- Viikki, M.; Kampman, O.; Illi, A.; Setälä-Soikkeli, E.; Anttila, S.; Huuhka, M.; Nuolivirta, T.; Poutanen, O.; Mononen, N.; Lehtimäki, T.; et al. TPH1 218A/C polymorphism is associated with major depressive disorder and its treatment response. Neurosci. Lett. 2010, 468, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Wigner, P.; Czarny, P.; Synowiec, E.; Bijak, M.; Białek, K.; Talarowska, M.; Galecki, P.; Szemraj, J.; Sliwinski, T. Association between single nucleotide polymorphisms of TPH1 and TPH2 genes, and depressive disorders. J. Cell. Mol. Med. 2018, 22, 1778–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Liu, F.; Yang, C.; Liang, X.; Shang, Q.; He, W.; Wang, Z. Association between the TPH1 A218C polymorphism and risk of mood disorders and alcohol dependence: Evidence from the current studies. J. Affect. Disord. 2012, 138, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Maurer-Spurej, E.; Pittendreigh, C.; Solomons, K. The influence of selective serotonin reuptake inhibitors on human platelet serotonin. Thromb. Haemost. 2004, 91, 119–128. [Google Scholar] [CrossRef]
- Quintana, J. Platelet serotonin and plasma tryptophan decreases in endogenous depression. Clinical, therapeutic, and biological correlations. J. Affect. Disord. 1992, 24, 55–62. [Google Scholar] [CrossRef]
- Berger, M.; Gray, J.A.; Roth, B.L. The Expanded Biology of Serotonin. Annu. Rev. Med. 2009, 60, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Bader, M. Phenotype of Mice Lacking Tryptophan Hydroxylase 1. In Serotonin; Elsevier: Amsterdam, The Netherlands, 2019; pp. 167–179. [Google Scholar]
- Suidan, G.L.; Duerschmied, D.; Dillon, G.M.; Vanderhorst, V.; Hampton, T.G.; Wong, S.L.; Voorhees, J.R.; Wagner, D.D. Lack of Tryptophan Hydroxylase-1 in Mice Results in Gait Abnormalities. PLoS ONE 2013, 8, e59032. [Google Scholar] [CrossRef]
- Savelieva, K.V.; Zhao, S.; Pogorelov, V.M.; Rajan, I.; Yang, Q.; Cullinan, E.; Lanthorn, T.H. Genetic Disruption of Both Tryptophan Hydroxylase Genes Dramatically Reduces Serotonin and Affects Behavior in Models Sensitive to Antidepressants. PLoS ONE 2008, 3, e3301. [Google Scholar] [CrossRef] [Green Version]
- Homberg, J.R.; Wöhr, M.; Alenina, N. Comeback of the Rat in Biomedical Research. ACS Chem. Neurosci. 2017, 8, 900–903. [Google Scholar] [CrossRef] [Green Version]
- Karpov, B.; Joffe, G.; Aaltonen, K.; Suvisaari, J.; Baryshnikov, I.; Näätänen, P.; Koivisto, M.; Melartin, T.; Oksanen, J.; Suominen, K.; et al. Anxiety symptoms in a major mood and schizophrenia spectrum disorders. Eur. Psychiatry 2016, 37, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Duncan, L.E.; Pollastri, A.R.; Smoller, J.W. Mind the gap: Why many geneticists and psychological scientists have discrepant views about gene–environment interaction (G×E) research. Am. Psychol. 2014, 69, 249–268. [Google Scholar] [CrossRef] [PubMed]
- Belsky, J.; Jonassaint, C.; Pluess, M.; Stanton, M.; Brummett, B.; Williams, R. Vulnerability genes or plasticity genes? Mol. Psychiatry 2009, 14, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Sbrini, G.; Brivio, P.; Bosch, K.; Homberg, J.R.; Calabrese, F. Enrichment environment positively influences depression- and anxiety-like behavior in serotonin transporter knockout rats through the modulation of neuroplasticitcity, spine and GABAergic markers. Genes 2020, 11, 1248. [Google Scholar] [CrossRef] [PubMed]
- Sbrini, G.; Brivio, P.; Peeva, P.M.; Todiras, M.; Bader, M.; Alenina, N.; Calabrese, F. The Absence of Serotonin in the Brain Alters Acute Stress Responsiveness by Interfering With the Genomic Function of the Glucocorticoid Receptors. Front. Cell. Neurosci. 2020, 14, 128. [Google Scholar] [CrossRef] [PubMed]
- Brivio, P.; Sbrini, G.; Peeva, P.; Todiras, M.; Bader, M.; Alenina, N.; Calabrese, F. TPH2 Deficiency Influences Neuroplastic Mechanisms and Alters the Response to an Acute Stress in a Sex Specific Manner. Front. Mol. Neurosci. 2018, 11, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łoś, K.; Waszkiewicz, N. Biological Markers in Anxiety Disorders. J. Clin. Med. 2021, 10, 1744. [Google Scholar] [CrossRef] [PubMed]
- Chiba, S.; Numakawa, T.; Ninomiya, M.; Richards, M.C.; Wakabayashi, C.; Kunugi, H. Chronic restraint stress causes anxiety- and depression-like behaviors, downregulates glucocorticoid receptor expression, and attenuates glutamate release induced by brain-derived neurotrophic factor in the prefrontal cortex. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 39, 112–119. [Google Scholar] [CrossRef]
- Brivio, P.; Sbrini, G.; Tarantini, L.; Parravicini, C.; Gruca, P.; Lason, M.; Litwa, E.; Favero, C.; Riva, M.A.; Eberini, I.; et al. Stress Modifies the Expression of Glucocorticoid-Responsive Genes by Acting at Epigenetic Levels in the Rat Prefrontal Cortex: Modulatory Activity of Lurasidone. Int. J. Mol. Sci. 2021, 22, 6197. [Google Scholar] [CrossRef]
- Brivio, P.; Sbrini, G.; Riva, M.A.; Calabrese, F. Acute Stress Induces Cognitive Improvement in the Novel Object Recognition Task by Transiently Modulating Bdnf in the Prefrontal Cortex of Male Rats. Cell. Mol. Neurobiol. 2020, 40, 1037–1047. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.; Gilbert, F. Activation of 5-HT(1A) receptor subtype in the paraventricular nuclei of the hypothalamus induces CRH and ACTH release in the rat. Neuroendocrinology 1992, 56, 797–802. [Google Scholar] [CrossRef]
- Herman, J.P.; Figueiredo, H.; Mueller, N.K.; Ulrich-Lai, Y.; Ostrander, M.M.; Choi, D.C.; Cullinan, W.E. Central mechanisms of stress integration: Hierarchical circuitry controlling hypothalamo-pituitary-adrenocortical responsiveness. Front. Neuroendocrinol. 2003, 24, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Van De Kar, L.D.; Javed, A.; Zhang, Y.; Serres, F.; Raap, D.K.; Gray, T.S. 5-HT2A receptors stimulate ACTH, corticosterone, oxytocin, renin, and prolactin release and activate hypothalamic CRF and oxytocin-expressing cells. J. Neurosci. 2001, 21, 3572–3579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duman, R.S.; Monteggia, L.M. A Neurotrophic Model for Stress-Related Mood Disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Namkung, J.; Hwang, I.; Kim, H.; Lim, A.; Park, H.J.; Lee, H.W.; Han, K.-H.; Park, S.; Jeong, J.-S.; et al. Serotonin signals through a gut-liver axis to regulate hepatic steatosis. Nat. Commun. 2018, 9, 4824. [Google Scholar] [CrossRef] [PubMed]
- Sumara, G.; Sumara, O.; Kim, J.K.; Karsenty, G. Gut-Derived Serotonin Is a Multifunctional Determinant to Fasting Adaptation. Cell Metab. 2012, 16, 588–600. [Google Scholar] [CrossRef] [Green Version]
- Mordhorst, A.; Dhandapani, P.; Matthes, S.; Mosienko, V.; Rothe, M.; Todiras, M.; Self, J.; Schunck, W.; Schütz, A.; Bader, M.; et al. Phenylalanine hydroxylase contributes to serotonin synthesis in mice. FASEB J. 2021, 35, e21648. [Google Scholar] [CrossRef]
- Cote, F.; Thevenot, E.; Fligny, C.; Fromes, Y.; Darmon, M.; Ripoche, M.-A.; Bayard, E.; Hanoun, N.; Saurini, F.; Lechat, P.; et al. Disruption of the nonneuronal tph1 gene demonstrates the importance of peripheral serotonin in cardiac function. Proc. Natl. Acad. Sci. USA 2003, 100, 13525–13530. [Google Scholar] [CrossRef] [Green Version]
- Crane, J.D.; Palanivel, R.; Mottillo, E.P.; Bujak, A.L.; Wang, H.; Ford, R.J.; Collins, A.; Blümer, R.M.; Fullerton, M.D.; Yabut, J.M.; et al. Inhibiting peripheral serotonin synthesis reduces obesity and metabolic dysfunction by promoting brown adipose tissue thermogenesis. Nat. Med. 2015, 21, 166–172. [Google Scholar] [CrossRef] [Green Version]
- Haq, S.; Grondin, J.A.; Khan, W.I. Tryptophan-derived serotonin-kynurenine balance in immune activation and intestinal inflammation. FASEB J. 2021, 35, e21888. [Google Scholar] [CrossRef]
- Fujiwara, M.; Shibata, M.; Watanabe, Y.; Nukiwa, T.; Hirata, F.; Mizuno, N.; Hayaishi, O. Indoleamine 2,3-dioxygenase. Formation of L-kynurenine from L-tryptophan in cultured rabbit fineal gland. J. Biol. Chem. 1978, 253, 6081–6085. [Google Scholar] [CrossRef]
- Goenjian, A.K.; Bailey, J.N.; Walling, D.P.; Steinberg, A.M.; Schmidt, D.; Dandekar, U.; Noble, E.P. Association of TPH1, TPH2, and 5HTTLPR with PTSD and depressive symptoms. J. Affect. Disord. 2012, 140, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Ons, S.; Marti, O.; Armario, A. Stress-induced activation of the immediate early gene Arc (activity-regulated cytoskeleton-associated protein) is restricted to telencephalic areas in the rat brain: Relationship to c-fos mRNA. J. Neurochem. 2004, 89, 1111–1118. [Google Scholar] [CrossRef] [PubMed]
- Molteni, R.; Calabrese, F.; Maj, P.F.; Olivier, J.D.A.; Racagni, G.; Ellenbroek, B.A.; Riva, M.A. Altered expression and modulation of activity-regulated cytoskeletal associated protein (Arc) in serotonin transporter knockout rats. Eur. Neuropsychopharmacol. 2009, 19, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Schoneveld, O.J.L.M.; Gaemers, I.C.; Lamers, W.H. Mechanisms of glucocorticoid signalling. Biochim. Et Biophys. Acta (BBA)-Gene Struct. Expr. 2004, 1680, 114–128. [Google Scholar] [CrossRef]
- Panettieri, R.A.; Schaafsma, D.; Amrani, Y.; Koziol-White, C.; Ostrom, R.; Tliba, O. Non-genomic Effects of Glucocorticoids: An Updated View. Trends Pharmacol. Sci. 2019, 40, 38–49. [Google Scholar] [CrossRef]
- Hegde, S.S.; Eglen, R.M. Peripheral 5-HT 4 receptors. FASEB J. 1996, 10, 1398–1407. [Google Scholar] [CrossRef]
- Jørgensen, H.; Knigge, U.; Kjaer, A.; Møller, M.; Warberg, J. Serotonergic stimulation of corticotropin-releasing hormone and pro-opiomelanocortin gene expression. J. Neuroendocrinol. 2002, 14, 788–795. [Google Scholar] [CrossRef]
- Chen, G.-L.; Miller, G.M. Advances in tryptophan hydroxylase-2 gene expression regulation: New insights into serotonin-stress interaction and clinical implications. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2012, 159, 152–171. [Google Scholar] [CrossRef] [Green Version]
- Lang, F.; Stournaras, C.; Alesutan, I. Regulation of transport across cell membranes by the serum- and glucocorticoid-inducible kinase SGK1. Mol. Membr. Biol. 2014, 31, 29–36. [Google Scholar] [CrossRef]
- Zannas, A.S.; Wiechmann, T.; Gassen, N.C.; Binder, E.B. Gene–Stress–Epigenetic Regulation of FKBP5: Clinical and Translational Implications. Neuropsychopharmacology 2016, 41, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D.; Nico, B.; Crivellato, E.; Artico, M. Development of the blood-brain barrier: A historical point of view. Anat. Rec. Part B New Anat. 2006, 289, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Bonnin, A.; Goeden, N.; Chen, K.; Wilson, M.L.; King, J.; Shih, J.C.; Blakely, R.D.; Deneris, E.S.; Levitt, P. A transient placental source of serotonin for the fetal forebrain. Nature 2011, 472, 347–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanthakumaran, S.; Sridharan, S.; Somagutta, M.R.; Arnold, A.A.; May, V.; Pagad, S.; Malik, B.H. The Gut-Brain Axis and Its Role in Depression. Cureus 2020, 12, e10280. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Sequence | Reverse Sequence | Probe Sequence |
|---|---|---|---|
| Total Bdnf | AAGTCTGCATTACATTCCTCGA | GTTTTCTGAAAGAGGGACAGTTTAT | TGTGGTTTGTTGCCGTTGCCAAG |
| Arc | GGTGGGTGGCTCTGAAGAAT | ACTCCACCCAGTTCTTCACC | GATCCAGAACCACATGAATGGG |
| Fos | TCCTTACGGACTCCCCAC | CTCCGTTTCTCTTCCTCTTCAG | TGCTCTACTTTGCCCCTTCTGCC |
| Dusp1 | TGTGCCTGACAGTGCAGAAT | ATCTTTCCGGGAAGCATGGT | ATCCTGTCCTTCCTGTACCT |
| Fkbp5 | GAACCCAATGCTGAGCTTATG | ATGTACTTGCCTCCCTTGAAG | TGTCCATCTCCCAGGATTCTTTGGC |
| Sgk1 | GACTACATTAATGGCGGAGAGC | AGGGAGTGCAGATAACCCAAG | TGCTCGCTTCTACGCAGC |
| Gilz | CGGTCTATCAACTGCACAATTTC | CTTCACTAGATCCATGGCCTG | AACGGAAACCACATCCCCTCCAA |
| 36b4 | TCAGTGCCTCACTCCATCAT | AGGAAGGCCTTGACCTTTTC | TGGATACAAAAGGGTCCTGG |
| Gene | Accession Number | Assay ID |
|---|---|---|
| Bdnf long 3′UTR | EF125675.1 | Rn02531967_s1 |
| Bdnf isoform IV | EF125679.1 | Rn01484927_m1 |
| Bdnf isoform VI | EF125680.1 | Rn01484928_m1 |
| Nr4a1 | BC097313.1 | Rn01452530_g1 |
| Gadd45β | BC085337.1 | Rn01452530_g1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sbrini, G.; Hanswijk, S.I.; Brivio, P.; Middelman, A.; Bader, M.; Fumagalli, F.; Alenina, N.; Homberg, J.R.; Calabrese, F. Peripheral Serotonin Deficiency Affects Anxiety-like Behavior and the Molecular Response to an Acute Challenge in Rats. Int. J. Mol. Sci. 2022, 23, 4941. https://doi.org/10.3390/ijms23094941
Sbrini G, Hanswijk SI, Brivio P, Middelman A, Bader M, Fumagalli F, Alenina N, Homberg JR, Calabrese F. Peripheral Serotonin Deficiency Affects Anxiety-like Behavior and the Molecular Response to an Acute Challenge in Rats. International Journal of Molecular Sciences. 2022; 23(9):4941. https://doi.org/10.3390/ijms23094941
Chicago/Turabian StyleSbrini, Giulia, Sabrina I. Hanswijk, Paola Brivio, Anthonieke Middelman, Michael Bader, Fabio Fumagalli, Natalia Alenina, Judith R. Homberg, and Francesca Calabrese. 2022. "Peripheral Serotonin Deficiency Affects Anxiety-like Behavior and the Molecular Response to an Acute Challenge in Rats" International Journal of Molecular Sciences 23, no. 9: 4941. https://doi.org/10.3390/ijms23094941