
The Daily Expression of ABCC4 at the BCSFB Affects the Transport of Its Substrate Methotrexate

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
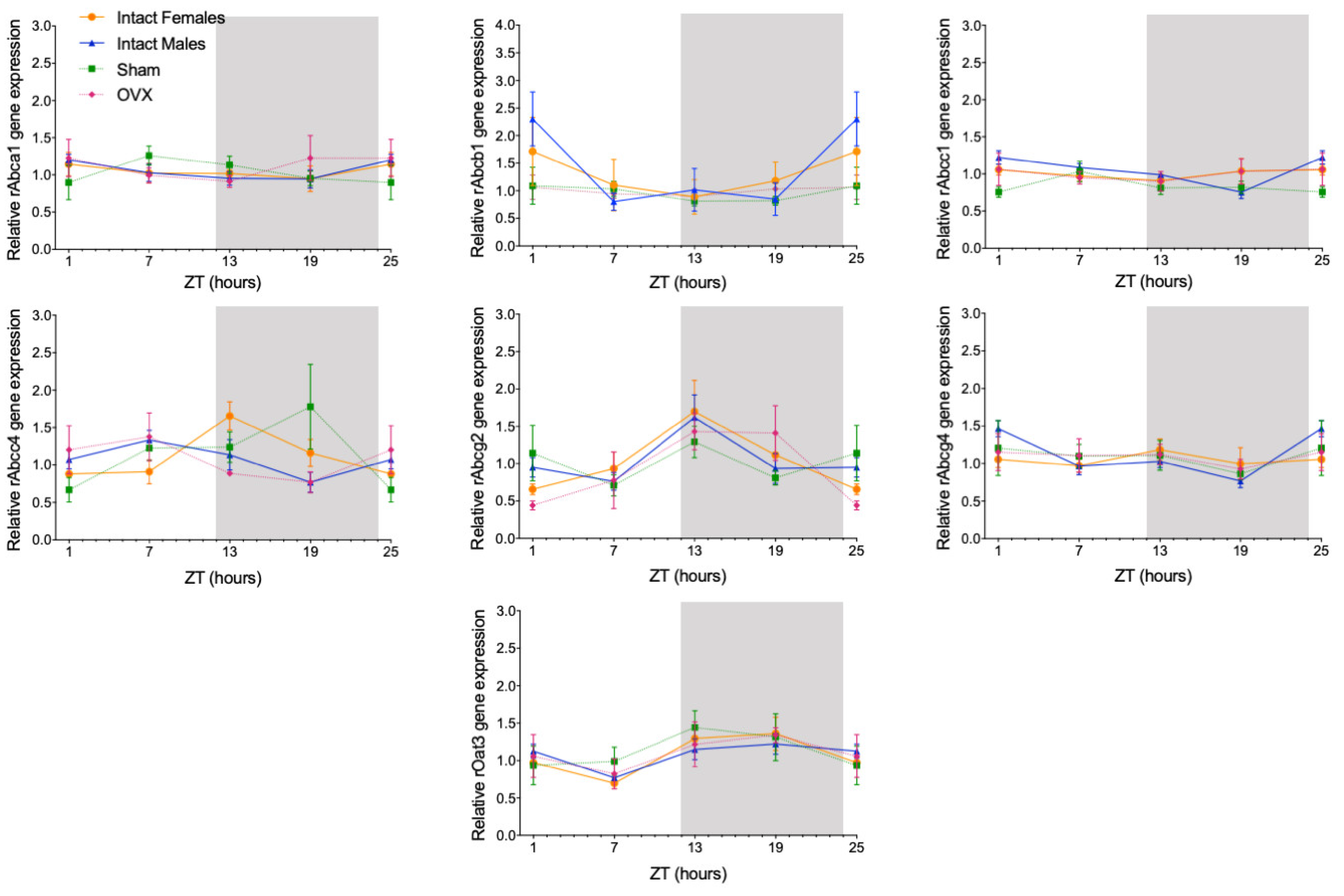
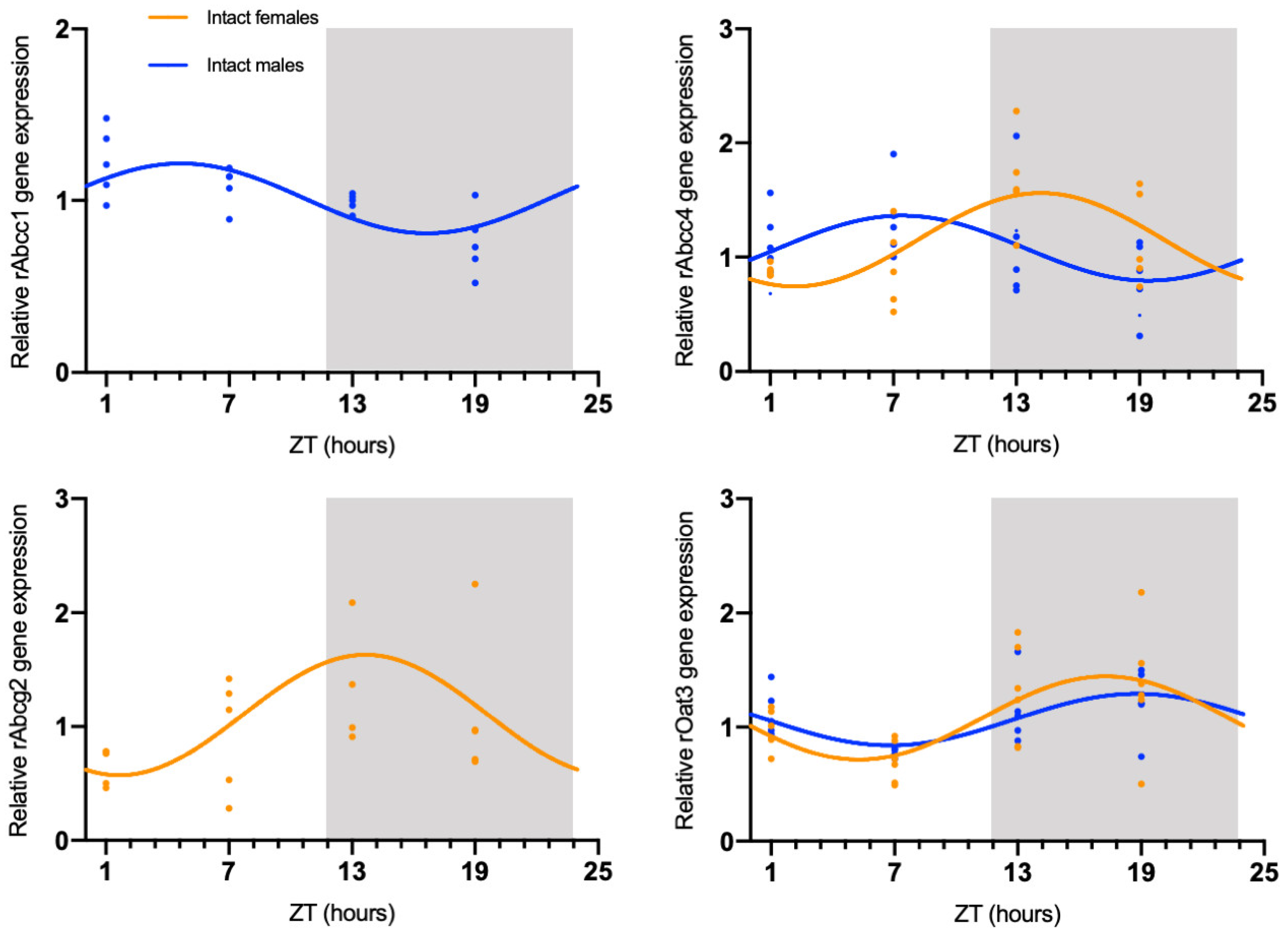
2.1. Sex-Dependent Daily Oscillations of Membrane Transporters in Rat Choroid Plexus
2.2. hAbcc4 and hAbcg2 mRNA Circadian Expression in the HIBCPP Cell Line
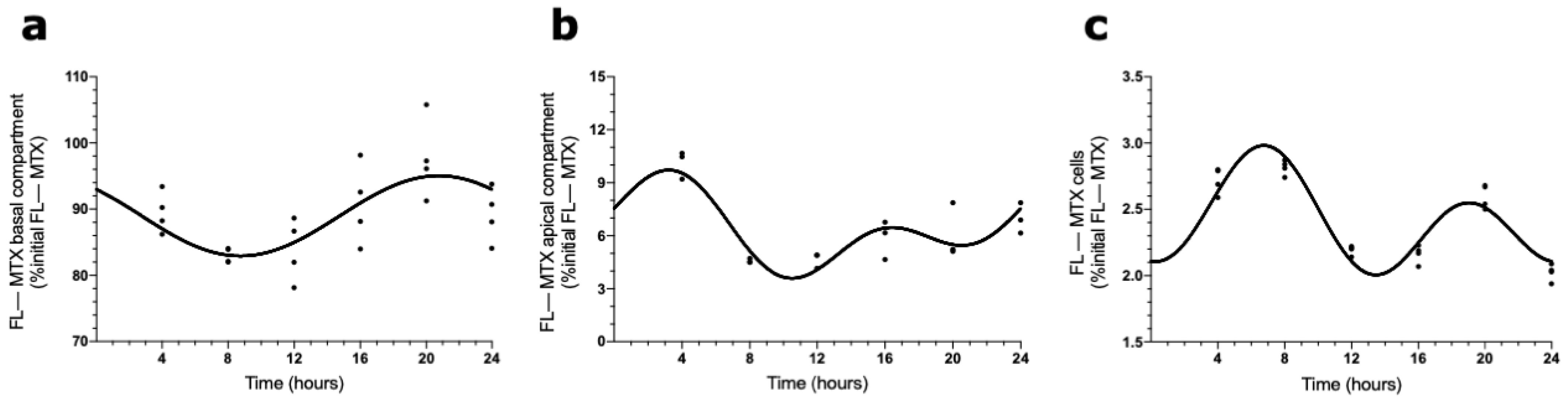
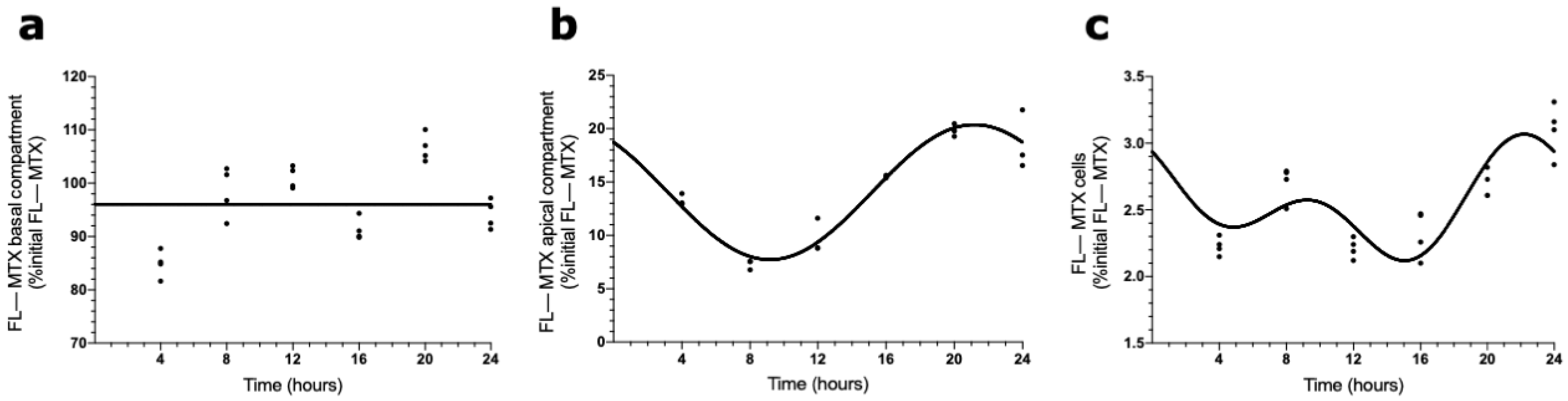
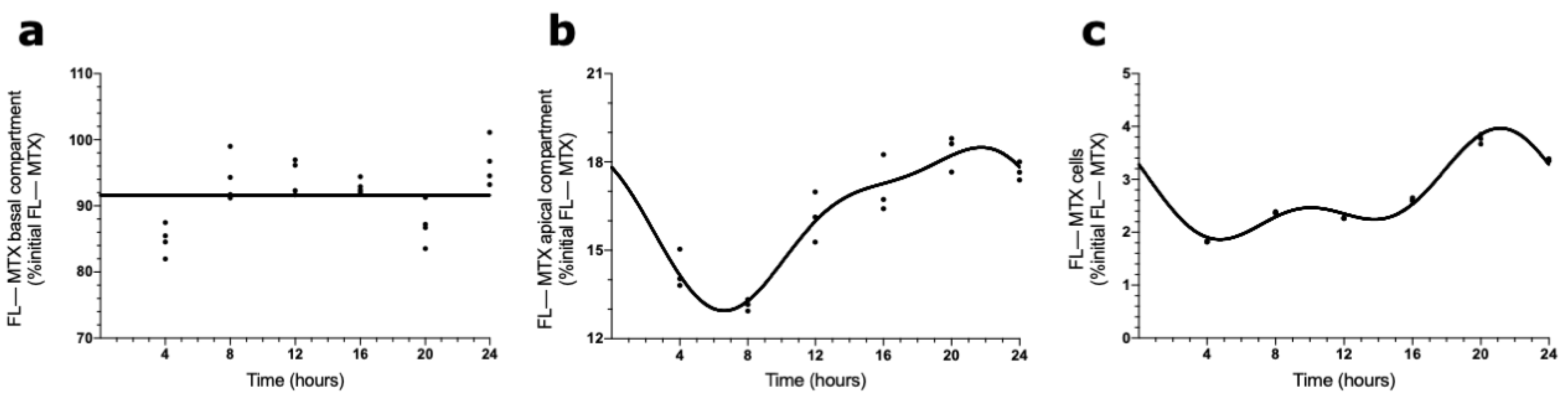
2.3. Circadian Oscillations in MTX Transport across BCSFB
2.4. Effects of ABCC4 and ABCG2 Inhibition in the MTX Circadian Transport across BCSFB
3. Discussion
4. Material and Methods
4.1. Animals and Cell Line
4.2. HIBCPP Cell Culture
4.3. Quantitative Real-Time PCR (qPCR)
4.4. MTX Uptake Assay
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bell-Pedersen, D.; Cassone, V.M.; Earnest, D.J.; Golden, S.S.; Hardin, P.E.; Thomas, T.L.; Zoran, M.J. Circadian rhythms from multiple oscillators: Lessons from diverse organisms. Nat. Rev. Genet. 2005, 6, 544–556. [Google Scholar] [CrossRef]
- Bhadra, U.; Thakkar, N.; Das, P.; Bhadra, M.P. Evolution of circadian rhythms: From bacteria to human. Sleep Med. 2017, 35, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.; Hogenesch, J.B.; Kay, S.A. Circadian rhythms from flies to human. Nature 2002, 417, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Hastings, M.H.; Maywood, E.S.; Brancaccio, M. Generation of circadian rhythms in the suprachiasmatic nucleus. Nat. Rev. Neurosci. 2018, 19, 453–469. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.S. Transcriptional architecture of the mammalian circadian clock. Nat. Rev. Genet. 2016, 18, 164–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pácha, J.; Balounová, K.; Soták, M. Circadian regulation of transporter expression and implications for drug disposition. Expert Opin. Drug Metab. Toxicol. 2021, 17, 425–439. [Google Scholar] [CrossRef]
- Zhao, M.; Xing, H.; Chen, M.; Dong, D.; Wu, B. Circadian clock-controlled drug metabolism and transport. Xenobiotica 2020, 50, 495–505. [Google Scholar] [CrossRef]
- Ballesta, A.; Innominato, P.F.; Dallmann, R.; Rand, D.A.; Lévi, F.A. Systems Chronotherapeutics. Pharmacol. Rev. 2017, 69, 161–199. [Google Scholar] [CrossRef] [Green Version]
- Gaspar, L.S.; Álvaro, A.R.; Carmo-Silva, S.; Mendes, A.F.; Relógio, A.; Cavadas, C. The importance of determining circadian parameters in pharmacological studies. Br. J. Pharmacol. 2019, 176, 2827–2847. [Google Scholar] [CrossRef]
- Baraldo, M. The influence of circadian rhythms on the kinetics of drugs in humans. Expert Opin. Drug Metab. Toxicol. 2008, 4, 175–192. [Google Scholar] [CrossRef]
- Ayyar, V.S.; Sukumaran, S. Circadian rhythms: Influence on physiology, pharmacology, and therapeutic interventions. J. Pharmacokinet. Pharmacodyn. 2021, 48, 321–338. [Google Scholar] [CrossRef] [PubMed]
- Zaki, N.F.W.; Yousif, M.; BaHammam, A.S.; Spence, D.W.; Bharti, V.K.; Subramanian, P.; Pandi-Perumal, S.R. Chronotherapeutics: Recognizing the Importance of Timing Factors in the Treatment of Disease and Sleep Disorders. Clin. Neuropharmacol. 2019, 42, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.R.; Duarte, A.C.; Quintela, T.; Tomás, J.F.M.; Albuqerque, T.; Marques, F.; Palha, J.; Gonçalves, I. The choroid plexus as a sex hormone target: Functional implications. Front. Neuroendocrinol. 2017, 44, 103–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghersi-Egea, J.-F.; Strazielle, N.; Catala, M.; Silva-Vargas, V.; Doetsch, F.; Engelhardt, B. Molecular anatomy and functions of the choroidal blood-cerebrospinal fluid barrier in health and disease. Acta Neuropathol. 2018, 135, 337–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, C.; Duarte, A.; Costa, A.R.; Tomás, J.; Quintela, T.; Gonçalves, I. The senses of the choroid plexus. Prog. Neurobiol. 2019, 182, 101680. [Google Scholar] [CrossRef] [PubMed]
- Quintela, T.; Albuqerque, T.; Lundkvist, G.; Belin, A.C.; Talhada, D.; Gonçalves, I.; Carro, E.; Santos, C.R. The choroid plexus harbors a circadian oscillator modulated by estrogens. Chronobiol. Int. 2018, 35, 270–279. [Google Scholar] [CrossRef]
- Quintela, T.; Sousa, C.; Patriarca, F.M.; Gonçalves, I.; Santos, C. Gender associated circadian oscillations of the clock genes in rat choroid plexus. Brain Struct. Funct. 2015, 220, 1251–1262. [Google Scholar] [CrossRef]
- Mohammad, I.S.; He, W.; Yin, L. Understanding of human ATP binding cassette superfamily and novel multidrug resistance modulators to overcome MDR. Biomed. Pharmacother. 2018, 100, 335–348. [Google Scholar] [CrossRef]
- Morris, M.E.; Rodriguez-Cruz, V.; Felmlee, M.A. SLC and ABC Transporters: Expression, Localization, and Species Differences at the Blood-Brain and the Blood-Cerebrospinal Fluid Barriers. AAPS J. 2017, 19, 1317–1331. [Google Scholar] [CrossRef]
- Leggas, M.; Adachi, M.; Scheffer, G.L.; Sun, D.; Wielinga, P.; Du, G.; Mercer, K.E.; Zhuang, Y.; Panetta, J.C.; Johnston, B.; et al. Mrp4 Confers Resistance to Topotecan and Protects the Brain from Chemotherapy. Mol. Cell. Biol. 2004, 24, 7612–7621. [Google Scholar] [CrossRef] [Green Version]
- Ishiwata, I.; Ishiwata, C.; Ishiwata, E.; Sato, Y.; Kiguchi, K.; Tachibana, T.; Hashimoto, H.; Ishikawa, H. Establishment and characterization of a human malignant choroids plexus papilloma cell line (HIBCPP). Hum. Cell 2005, 18, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Ballesta, A.; Dulong, S.; Abbara, C.; Cohen, B.; Okyar, A.; Clairambault, J.; Levi, F. A combined experimental and mathematical approach for molecular-based optimization of irinotecan circadian delivery. PLoS Comput. Biol. 2011, 7, e1002143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stearns, A.; Balakrishnan, A.; Rhoads, D.B.; Ashley, S.W.; Tavakkolizadeh, A. Diurnal Rhythmicity in the Transcription of Jejunal Drug Transporters. J. Pharmacol. Sci. 2008, 108, 144–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gachon, F.; Olela, F.F.; Schaad, O.; Descombes, P.; Schibler, U. The circadian PAR-domain basic leucine zipper transcription factors DBP, TEF, and HLF modulate basal and inducible xenobiotic detoxification. Cell Metab. 2006, 4, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Kotaka, M.; Onishi, Y.; Ohno, T.; Akaike, T.; Ishida, N. Identification of negative transcriptional factor E4BP4-binding site in the mouse circadian-regulated gene Mdr2. Neurosci. Res. 2008, 60, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Murakami, Y.; Higashi, Y.; Matsunaga, N.; Koyanagi, S.; Ohdo, S. Circadian Clock-Controlled Intestinal Expression of the Multidrug-Resistance Gene mdr1a in Mice. Gastroenterology 2008, 135, 1636–1644.e3. [Google Scholar] [CrossRef]
- Assem, M.; Schuetz, E.G.; Leggas, M.; Sun, D.; Yasuda, K.; Reid, G.; Zelcer, N.; Adachi, M.; Strom, S.; Evans, R.; et al. Interactions between Hepatic Mrp4 and Sult2a as Revealed by the Constitutive Androstane Receptor and Mrp4 Knockout Mice. J. Biol. Chem. 2004, 279, 22250–22257. [Google Scholar] [CrossRef] [Green Version]
- Renga, B.; Migliorati, M.; Mencarelli, A.; Cipriani, S.; D’Amore, C.; Distrutti, E.; Fiorucci, S. Farnesoid X receptor suppresses constitutive androstane receptor activity at the multidrug resistance protein-4 promoter. Biochim. Biophys. Acta 2011, 1809, 157–165. [Google Scholar] [CrossRef]
- Whyte-Allman, S.-K.; Hoque, T.; Jenabian, M.-A.; Routy, J.-P.; Bendayan, R. Xenobiotic Nuclear Receptors Pregnane X Receptor and Constitutive Androstane Receptor Regulate Antiretroviral Drug Efflux Transporters at the Blood-Testis Barrier. J. Pharmacol. Exp. Ther. 2017, 363, 324–335. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Weerachayaphorn, J.; Cai, S.-Y.; Soroka, C.J.; Boyer, J.L. Aryl hydrocarbon receptor and NF-E2-related factor 2 are key regulators of human MRP4 expression. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G126–G135. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-K.J.; Yeager, R.L.; Klaassen, C.D. Circadian Expression Profiles of Drug-Processing Genes and Transcription Factors in Mouse Liver. Drug Metab. Dispos. 2009, 37, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, V.M.; Santostefano, M.J.; Birnbaum, L.S. Daily cycle of bHLH-PAS proteins, Ah receptor and Arnt, in multiple tissues of female Sprague-Dawley rats. Biochem. Biophys. Res. Commun. 1998, 252, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.; Choi, Y.; Shim, J.; Yoo, I.; Ka, H. Prostaglandin Transporters ABCC4 and SLCO2A1 in the Uterine Endometrium and Conceptus During Pregnancy in Pigs. Biol. Reprod. 2014, 90, 100. [Google Scholar] [CrossRef] [PubMed]
- Maher, J.; Cheng, X.; Tanaka, Y.; Scheffer, G.; Klaassen, C. Hormonal regulation of renal multidrug resistance-associated proteins 3 and 4 (Mrp3 and Mrp4) in mice. Biochem. Pharmacol. 2006, 71, 1470–1478. [Google Scholar] [CrossRef]
- Koraïchi, F.; Inoubli, L.; Lakhdari, N.; Meunier, L.; Vega, A.; Mauduit, C.; Benahmed, M.; Prouillac, C.; Lecoeur, S. Neonatal exposure to zearalenone induces long term modulation of ABC transporter expression in testis. Toxicology 2013, 310, 29–38. [Google Scholar] [CrossRef]
- Cai, C.; Omwancha, J.; Hsieh, C.-L.; Shemshedini, L. Androgen induces expression of the multidrug resistance protein gene MRP4 in prostate cancer cells. Prostate Cancer Prostatic Dis. 2007, 10, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Ho, L.L.; Kench, J.; Handelsman, D.; Scheffer, G.; Stricker, P.; Grygiel, J.; Sutherland, R.; Henshall, S.; Allen, J.; Horvath, L. Androgen regulation of multidrug resistance-associated protein 4 (MRP4/ABCC4) in prostate cancer. Prostate 2008, 68, 1421–1429. [Google Scholar] [CrossRef]
- Mihara, T.; Kikuchi, T.; Kamiya, Y.; Koga, M.; Uchimoto, K.; Kurahashi, K.; Goto, T. Day or Night Administration of Ketamine and Pentobarbital Differentially Affect Circadian Rhythms of Pineal Melatonin Secretion and Locomotor Activity in Rats. Anesthesia Analg. 2012, 115, 805–813. [Google Scholar] [CrossRef]
- Hamdan, A.M.; Koyanagi, S.; Wada, E.; Kusunose, N.; Murakami, Y.; Matsunaga, N.; Ohdo, S. Intestinal expression of mouse Abcg2/breast cancer resistance protein (BCRP) gene is under control of circadian clock-activating transcription factor-4 pathway. J. Biol. Chem. 2012, 287, 17224–17231. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.H.; Lee, J.; Han, D.-H.; Cho, S.; Lee, Y.-J. Circadian Clock Is Involved in Regulation of Hepatobiliary Transport Mediated by Multidrug Resistance-Associated Protein 2. J. Pharm. Sci. 2017, 106, 2491–2498. [Google Scholar] [CrossRef] [Green Version]
- Nikolaeva, S.; Ansermet, C.; Centeno, G.; Pradervand, S.; Bize, V.; Mordasini, D.; Henry, H.; Koesters, R.; Maillard, M.; Bonny, O.; et al. Nephron-Specific Deletion of Circadian Clock Gene Bmal1 Alters the Plasma and Renal Metabolome and Impairs Drug Disposition. J. Am. Soc. Nephrol. 2016, 27, 2997–3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinken, M.; Halwachs, S.; Kneuer, C.; Honscha, W. Subcellular localization and distribution of the reduced folate carrier in normal rat tissues. Eur. J. Histochem. 2011, 55, e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.-S.; Hopper-Borge, E.; Belinsky, M.G.; Shchaveleva, I.; Kotova, E.; Kruh, G.D. Characterization of the Transport Properties of Human Multidrug Resistance Protein 7 (MRP7, ABCC10). Mol. Pharmacol. 2003, 63, 351–358. [Google Scholar] [CrossRef]
- Inoue, K.; Yuasa, H. Molecular Basis for Pharmacokinetics and Pharmacodynamics of Methotrexate in Rheumatoid Arthritis Therapy. Drug Metab. Pharmacokinet. 2014, 29, 12–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, A.; Bernardes, M.; Azevedo, R.; Monteiro, J.; Sousa, H.; Medeiros, R.; Seabra, V. SLC19A1, SLC46A1 and SLCO1B1 Polymorphisms as Predictors of Methotrexate-Related Toxicity in Portuguese Rheumatoid Arthritis Patients. Toxicol. Sci. 2014, 142, 196–209. [Google Scholar] [CrossRef] [Green Version]
- Norris, M.D.; De Graaf, D.; Haber, M.; Kavallaris, M.; Madafiglio, J.; Gilbert, J.; Kwan, E.; Stewart, B.; Mechetner, E.; Gudkov, A.; et al. Involvement of MDR1 P-glycoprotein in multifactorial resistance to methotrexate. Int. J. Cancer 1996, 65, 613–619. [Google Scholar] [CrossRef]
- Pizzagalli, F.; Hagenbuch, B.; Stieger, B.; Klenk, U.; Folkers, G.; Meier, P.J. Identification of a Novel Human Organic Anion Transporting Polypeptide as a High Affinity Thyroxine Transporter. Mol. Endocrinol. 2002, 16, 2283–2296. [Google Scholar] [CrossRef] [Green Version]
- Tamai, I.; Sai, Y.; Ono, A.; Kido, Y.; Yabuuchi, H.; Takanaga, H.; Satoh, E.; Ogihara, T.; Amano, O.; Izeki, S.; et al. Immunohistochemical and functional characterization of pH-dependent intestinal absorption of weak organic acids by the monocarboxylic acid transporter MCT1. J. Pharm. Pharmacol. 1999, 51, 1113–1121. [Google Scholar] [CrossRef]
- Assaraf, Y.G. The role of multidrug resistance efflux transporters in antifolate resistance and folate homeostasis. Drug Resist. Updates 2006, 9, 227–246. [Google Scholar] [CrossRef]
- Stieger, B.; Gao, B. Drug transporters in the central nervous system. Clin. Pharmacokinet. 2015, 54, 225–242. [Google Scholar] [CrossRef] [Green Version]
- Wollack, J.B.; Makori, B.; Ahlawat, S.; Koneru, R.; Picinich, S.C.; Smith, A.; Goldman, I.D.; Qiu, A.; Cole, P.D.; Glod, J.; et al. Characterization of folate uptake by choroid plexus epithelial cells in a rat primary culture model. J. Neurochem. 2008, 104, 1494–1503. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Min, S.H.; Wang, Y.; Puchulu-Campanella, E.; Low, P.; Goldman, I.D. A Role for the Proton-coupled Folate Transporter (PCFT-SLC46A1) in Folate Receptor-mediated Endocytosis. J. Biol. Chem. 2009, 284, 4267–4274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazzin, S.; Strazielle, N.; Schmitt, C.; Fevre-Montange, M.; Ostrow, J.D.; Tiribelli, C.; Ghersi-Egea, J.-F. Differential expression of the multidrug resistance-related proteins ABCb1 and ABCc1 between blood-brain interfaces. J. Comp. Neurol. 2008, 510, 497–507. [Google Scholar] [CrossRef]
- Roberts, L.M.; Woodford, K.; Zhou, M.; Black, D.S.; Haggerty, J.E.; Tate, E.H.; Grindstaff, K.K.; Mengesha, W.; Raman, C.; Zerangue, N. Expression of the Thyroid Hormone Transporters Monocarboxylate Transporter-8 (SLC16A2) and Organic Ion Transporter-14 (SLCO1C1) at the Blood-Brain Barrier. Endocrinology 2008, 149, 6251–6261. [Google Scholar] [CrossRef]
- Tachikawa, M.; Watanabe, M.; Hori, S.; Fukaya, M.; Ohtsuki, S.; Asashima, T.; Terasaki, T. Distinct spatio-temporal expression of ABCA and ABCG transporters in the developing and adult mouse brain. J. Neurochem. 2005, 95, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Grapp, M.; Wrede, A.; Schweizer, M.; Hüwel, S.; Galla, H.-J.; Snaidero, N.; Simons, M.; Bückers, J.; Low, P.S.; Urlaub, H.; et al. Choroid plexus transcytosis and exosome shuttling deliver folate into brain parenchyma. Nat. Commun. 2013, 4, 2123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prémaud, A.; Rousseau, A.; Gicquel, M.; Ragot, S.; Manceau, J.; Laurentie, M.; Marquet, P. An Animal Model for the Study of Chronopharmacokinetics of Drugs and Application to Methotrexate and Vinorelbine. Toxicol. Appl. Pharmacol. 2002, 183, 189–197. [Google Scholar] [CrossRef]
- Bjarnason, G.A. Clinical chronotolerance to anticancer drugs: Relevance for dose-intensity. In Vivo 1995, 9, 557–564. [Google Scholar]
- Ferrazzini, G.; Sohl, H.; Robieux, I.; Johnson, D.; Giesbrecht, E.; Koren, G. Diurnal variation of methotrexate disposition in children with acute leukaemia. Eur. J. Clin. Pharmacol. 1991, 41, 425–427. [Google Scholar] [CrossRef]
- Gumustekin, M.; Kalkan, S.; Murat, N.; Gur, O.; Hocaoglu, N.; Gidener, S. The role of circadian rhythm on the pharmacokinetic of methotrexate in streptozotocin-induced diabetes mellitus rats. Biol. Rhythm Res. 2005, 36, 277–285. [Google Scholar] [CrossRef]
- Sane, R.; Wu, S.-P.; Zhang, R.; Gallo, J.M. The Effect of ABCG2 and ABCC4 on the Pharmacokinetics of Methotrexate in the Brain. Drug Metab. Dispos. 2014, 42, 537–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernd, A.; Ott, M.; Ishikawa, H.; Schroten, H.; Schwerk, C.; Fricker, G. Characterization of efflux transport proteins of the human choroid plexus papilloma cell line HIBCPP, a functional in vitro model of the blood-cerebrospinal fluid barrier. Pharm. Res. 2015, 32, 2973–2982. [Google Scholar] [CrossRef] [PubMed]
- Schwerk, C.; Papandreou, T.; Schuhmann, D.; Nickol, L.; Borkowski, J.; Steinmann, U.; Quednau, N.; Stump, C.; Weiss, C.; Berger, J.; et al. Polar Invasion and Translocation of Neisseria meningitidis and Streptococcus suis in a Novel Human Model of the Blood-Cerebrospinal Fluid Barrier. PLoS ONE 2012, 7, e30069. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Gil-Martins, E.; Barbosa, D.J.; Silva, V.; Remião, F.; Silva, R. Dysfunction of ABC transporters at the blood-brain barrier: Role in neurological disorders. Pharmacol. Ther. 2020, 213, 107554. [Google Scholar] [CrossRef]
- Duarte, A.C.; Furtado, A.; Hrynchak, M.V.; Costa, A.R.; Talhada, D.; Gonçalves, I.; Lemos, M.C.; Quintela, T.; Santos, C.R. Age, Sex Hormones, and Circadian Rhythm Regulate the Expression of Amyloid-Beta Scavengers at the Choroid Plexus. Int. J. Mol. Sci. 2020, 21, 6813. [Google Scholar] [CrossRef]
- Van der Spek, R.; Fliers, E.; la Fleur, S.; Kalsbeek, A. Daily Gene Expression Rhythms in Rat White Adipose Tissue Do Not Differ Between Subcutaneous and Intra-Abdominal Depots. Front. Endocrinol. 2018, 9, 206. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| rAbcc1 | Intact males | p-value = 0.0086 COG = 4.65 |
| rAbcc4 | Intact females | p-value = 0.0080 COG = 14.20 |
| Intact males | p-value = 0.0429 COG = 7.43 | |
| rAbcg2 | Intact females | p-value = 0.0447 COG = 13.66 |
| rOat3 | Intact females | p-value = 0.0079 COG = 17.27 |
| Intact males | p-value = 0.0312 COG = 18.80 |
| hAbcc4 | p-value = 0.0068 COG = 9.91 |
| hAbcg2 | p-value = 0.0001 COG = 10.74 |
| Gene | Primer Sequence (5′–3′) | Amplicon Size (bp) |
|---|---|---|
| hAbcc4 | FW: TGTGGCTTTGAACACAGCGTA | 105 |
| RV: CCAGCACACTGAACGTGATAA | ||
| hAbcg2 | FW: ACGAACGGATTAACAGGGTCA | 93 |
| RV: CTCCAGACACACCACGGAT | ||
| hGapdh * | FW: ATGGGGAAGGTGAAGGTCG | 108 |
| RV: GGGGTCATTGATGGCAACAATA | ||
| rAbca1 | FW: CGGCGGAGTAGAAAGGGTTT | 84 |
| RV: CACGATCAGGCTGAAGACCAG | ||
| rAbcb1 | FW: AAGGGGCTACAGGGTCTAGG | 100 |
| RV: AGTGTCAATTGCCAGCCGTA | ||
| rAbcc1 | FW: TTCATATCTGCTTCGTCACCG | 60 |
| RV: CGTAAACAGCACCCACCACAGC | ||
| rAbcc4 | FW: TTCCCCTTCGACCTTATCCT | 124 |
| RV: TAGGCAGCTGTTGTCAGTGG | ||
| rAbcg2 | FW: GGCCTGGACAAAGTAGCAGA | 137 |
| RV: GTTGTGGGCTCATCCAGGAA | ||
| rAbcg4 | FW: ATGGCTGATGTACCCTTCCAGGTT | 155 |
| RV: ATCAAGAGTCCCAAAGACTGGGCA | ||
| rOat3 | FW: GAGGACCTGTGATTGGAGAACTG | 82 |
| RV: CTG GCT GCC AGC ATG AGA TA | ||
| rCycA * | FW: CAAGACTGAGTGGCTGGATGG | 163 |
| RV: GCCCGCAAGTCAAAGAAATTAGAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furtado, A.; Mineiro, R.; Duarte, A.C.; Gonçalves, I.; Santos, C.R.; Quintela, T. The Daily Expression of ABCC4 at the BCSFB Affects the Transport of Its Substrate Methotrexate. Int. J. Mol. Sci. 2022, 23, 2443. https://doi.org/10.3390/ijms23052443
Furtado A, Mineiro R, Duarte AC, Gonçalves I, Santos CR, Quintela T. The Daily Expression of ABCC4 at the BCSFB Affects the Transport of Its Substrate Methotrexate. International Journal of Molecular Sciences. 2022; 23(5):2443. https://doi.org/10.3390/ijms23052443
Chicago/Turabian StyleFurtado, André, Rafael Mineiro, Ana Catarina Duarte, Isabel Gonçalves, Cecília R. Santos, and Telma Quintela. 2022. "The Daily Expression of ABCC4 at the BCSFB Affects the Transport of Its Substrate Methotrexate" International Journal of Molecular Sciences 23, no. 5: 2443. https://doi.org/10.3390/ijms23052443