
Expression and Function of Nicotinic Acetylcholine Receptors in Induced Regulatory T Cells


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
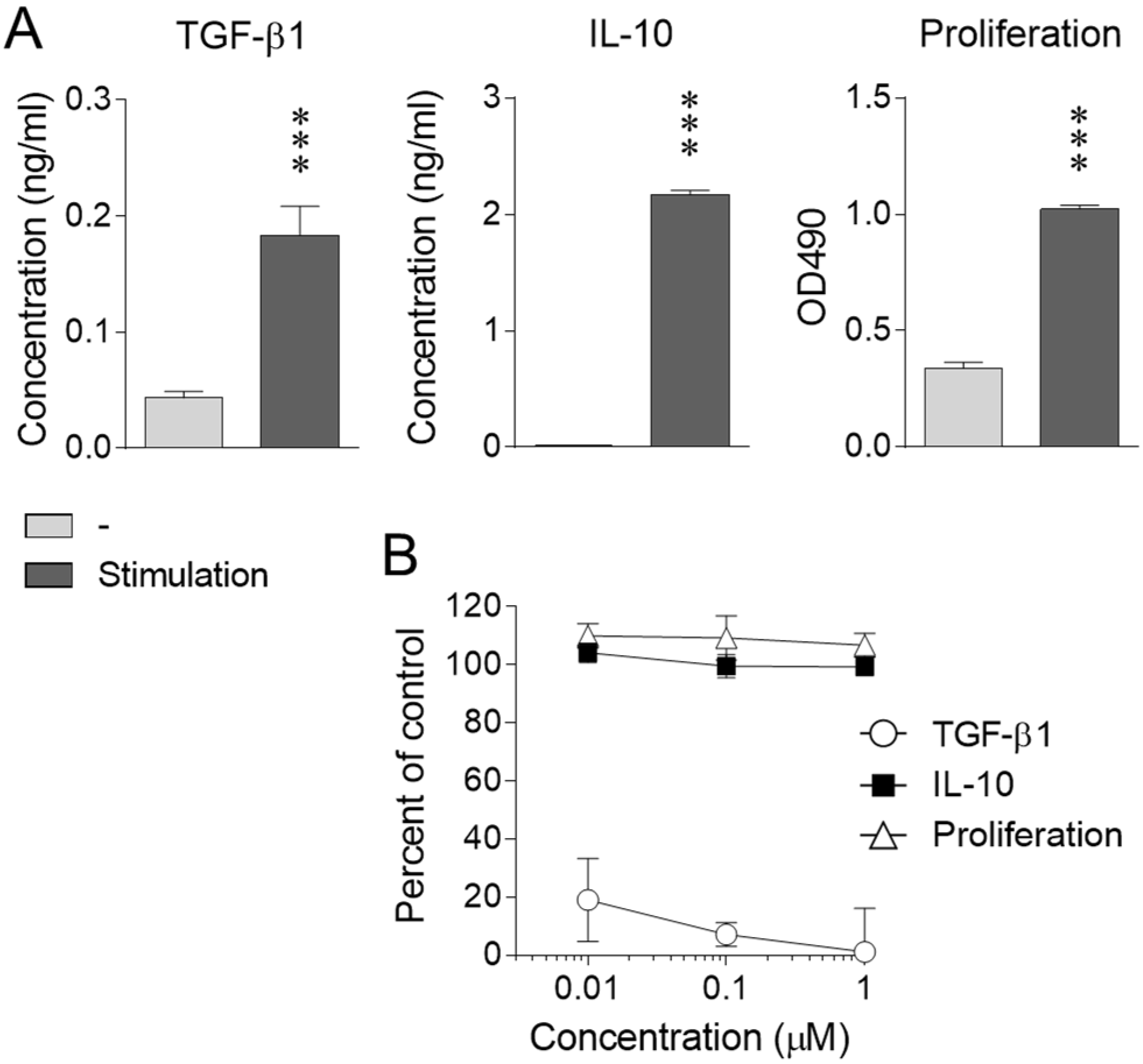
2.1. Effect of Nicotine on iTreg Cells
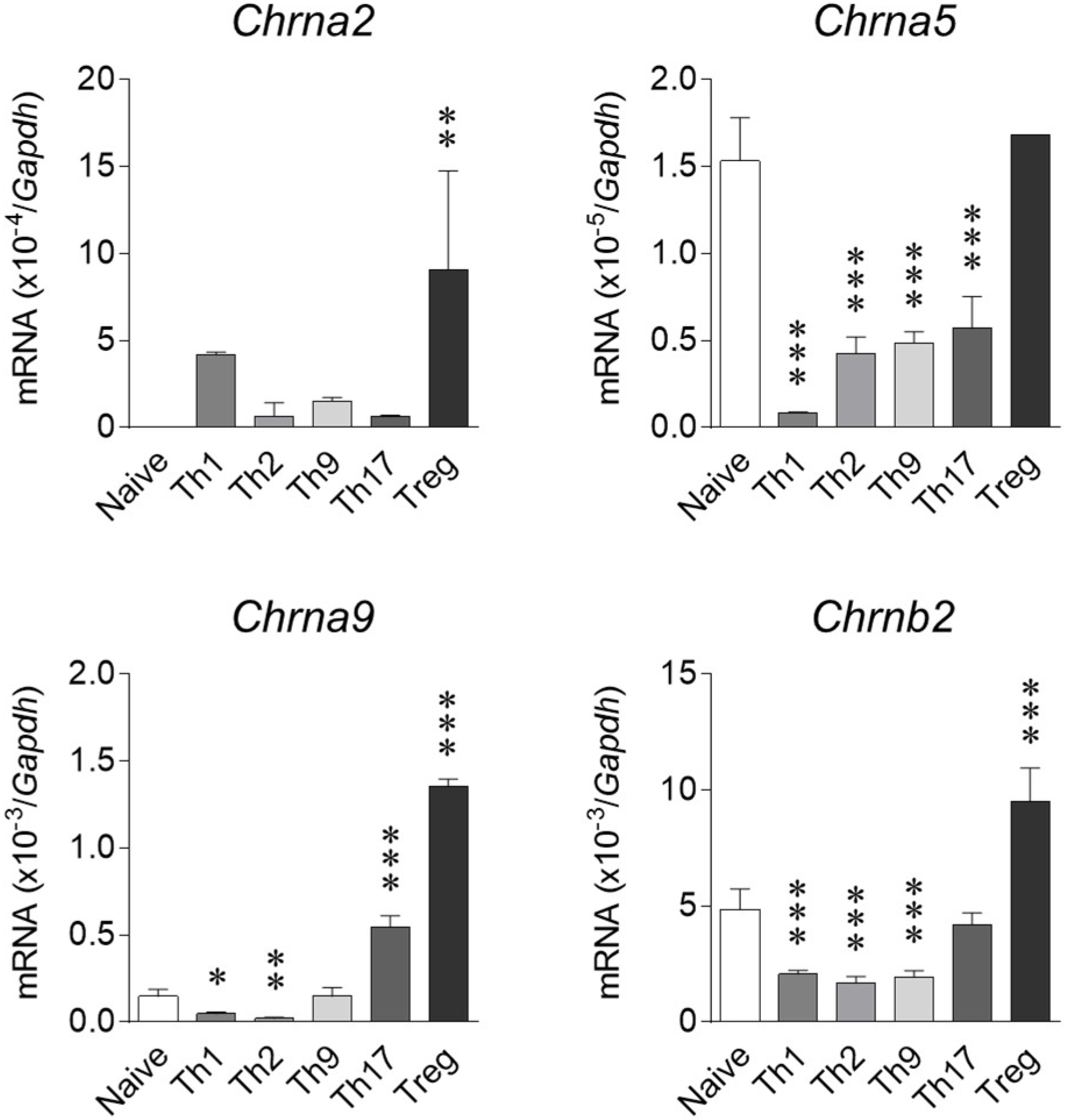
2.2. Expression of nAChRs on T Cell Subsets
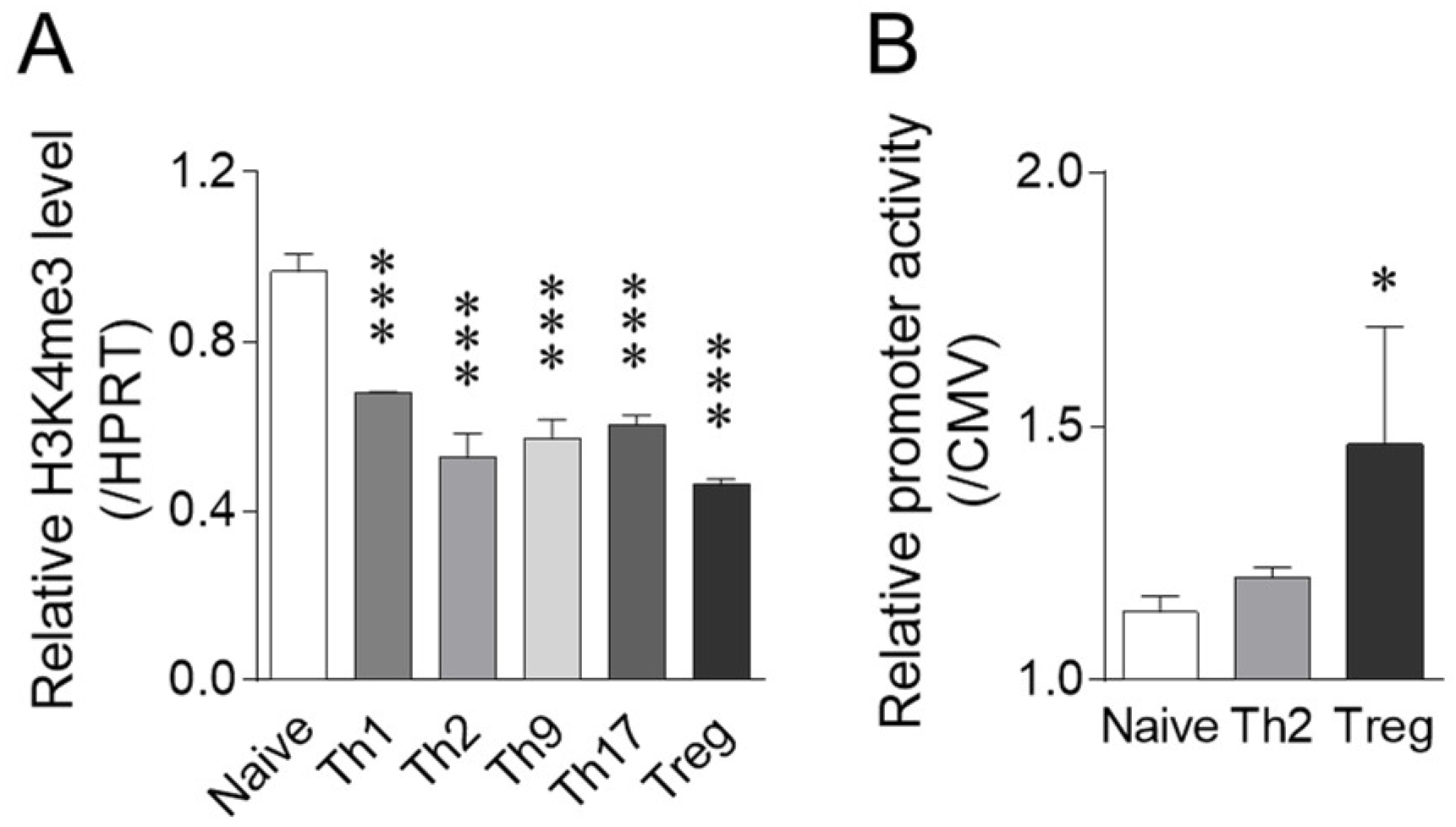
2.3. Mechanisms Regulating nAChR9 Expression in iTreg Cells
3. Discussion
4. Materials and Methods
4.1. T-Cell Subset Preparation
4.2. Cytokine Production and Proliferation of iTreg Cells
4.3. Expression of nAChRs
4.4. Chromatin Immunoprecipitation (ChIP) Assay
4.5. Reporter Assay
4.6. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Kalamida, D.; Poulas, K.; Avramopoulou, V.; Fostieri, E.; Lagoumintzis, G.; Lazaridis, K.; Sideri, A.; Zouridakis, M.; Tzartos, S.J. Muscle and neuronal nicotinic acetylcholine receptors. Structure, function and pathogenicity. FEBS J. 2007, 274, 3799–3845. [Google Scholar] [CrossRef] [PubMed]
- Cordero-Erausquin, M.; Marubio, L.M.; Klink, R.; Changeux, J.P. Nicotinic receptor function: New perspectives from knockout mice. Trends Pharmacol. Sci. 2000, 21, 211–217. [Google Scholar] [CrossRef]
- Reale, M.; Costantini, E. Cholinergic Modulation of the Immune System in Neuroinflammatory Diseases. Diseases 2021, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Mashimo, M.; Moriwaki, Y.; Misawa, H.; Kawashima, K.; Fujii, T. Regulation of Immune Functions by Non-Neuronal Acetylcholine (ACh) via Muscarinic and Nicotinic ACh Receptors. Int. J. Mol. Sci. 2021, 22, 6818. [Google Scholar] [CrossRef] [PubMed]
- Mazloomi, E.; Ilkhanizadeh, B.; Zare, A.; Mohammadzadeh, A.; Delirezh, N.; Shahabi, S. Evaluation of the efficacy of nicotine in treatment of allergic asthma in BALB/c mice. Int. Immunopharmacol. 2018, 63, 239–245. [Google Scholar] [CrossRef]
- Blanchet, M.R.; Israël-Assayag, E.; Cormier, Y. Modulation of airway inflammation and resistance in mice by a nicotinic receptor agonist. Eur. Respir. J. 2005, 26, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Inoue, K.; Ogura, A.; Kaminuma, O. Role of CD4(+) T Cells in Allergic Airway Diseases: Learning from Murine Models. Int. J. Mol. Sci. 2020, 21, 7480. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Fu, D.; Bidgoli, A.; Paczesny, S. T Cell Subsets in Graft Versus Host Disease and Graft Versus Tumor. Front. Immunol. 2021, 12, 761448. [Google Scholar] [CrossRef] [PubMed]
- Kaminuma, O.; Ohtomo, T.; Mori, A.; Nagakubo, D.; Hieshima, K.; Ohmachi, Y.; Noda, Y.; Katayama, K.; Suzuki, K.; Motoi, Y.; et al. Selective down-regulation of Th2 cell-mediated airway inflammation in mice by pharmacological intervention of CCR4. Clin. Exp. Allergy 2012, 42, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Saeki, M.; Kaminuma, O.; Nishimura, T.; Kitamura, N.; Mori, A.; Hiroi, T. Th9 cells elicit eosinophil-independent bronchial hyperresponsiveness in mice. Allergol. Int. 2016, 65, S24–S29. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, N.; Kaminuma, O.; Kitamura, N.; Hiroi, T. Induced Treg cells augment the Th17-mediated intestinal inflammatory response in a CTLA4-dependent manner. PLoS ONE 2016, 11, e0150244. [Google Scholar] [CrossRef] [PubMed]
- Galitovskiy, V.; Qian, J.; Chernyavsky, A.I.; Marchenko, S.; Gindi, V.; Edwards, R.A.; Grando, S.A. Cytokine-induced alterations of α7 nicotinic receptor in colonic CD4 T cells mediate dichotomous response to nicotine in murine models of Th1/Th17- versus Th2-mediated colitis. J. Immunol. 2011, 187, 2677–2687. [Google Scholar] [CrossRef] [PubMed]
- Li, M.O.; Flavell, R.A. TGF-beta: A master of all T cell trades. Cell 2008, 134, 392–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashimo, M.; Iwasaki, Y.; Inoue, S.; Saito, S.; Kawashima, K.; Fujii, T. Acetylcholine released from T cells regulates intracellular Ca(2+), IL-2 secretion and T cell proliferation through nicotinic acetylcholine receptor. Life Sci. 2017, 172, 13–18. [Google Scholar] [CrossRef]
- Qian, J.; Galitovskiy, V.; Chernyavsky, A.I.; Marchenko, S.; Grando, S.A. Plasticity of the murine spleen T-cell cholinergic receptors and their role in in vitro differentiation of naïve CD4 T cells toward the Th1, Th2 and Th17 lineages. Genes Immun. 2011, 12, 222–230. [Google Scholar] [CrossRef] [Green Version]
- Mashimo, M.; Komori, M.; Matsui, Y.Y.; Murase, M.X.; Fujii, T.; Takeshima, S.; Okuyama, H.; Ono, S.; Moriwaki, Y.; Misawa, H.; et al. Distinct Roles of α7 nAChRs in Antigen-Presenting Cells and CD4(+) T Cells in the Regulation of T Cell Differentiation. Front. Immunol. 2019, 10, 1102. [Google Scholar] [CrossRef]
- Nishimura, T.; Kaminuma, O.; Saeki, M.; Kitamura, N.; Matsuoka, K.; Yonekawa, H.; Mori, A.; Hiroi, T. Essential contribution of CD4+ T cells to antigen-induced nasal hyperresponsiveness in experimental allergic rhinitis. PLoS ONE 2016, 11, e0146686. [Google Scholar] [CrossRef]
- Bessis, A.; Savatier, N.; Devillers-Thiéry, A.; Bejanin, S.; Changeux, J.P. Negative regulatory elements upstream of a novel exon of the neuronal nicotinic acetylcholine receptor alpha 2 subunit gene. Nucleic Acids Res. 1993, 21, 2185–2192. [Google Scholar] [CrossRef] [Green Version]
- Shilatifard, A. Molecular implementation and physiological roles for histone H3 lysine 4 (H3K4) methylation. Curr. Opin. Cell Biol. 2008, 20, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, T.; Kaminuma, O.; Saeki, M.; Kitamura, N.; Mori, A.; Hiroi, T. Suppressive effect of environmental tobacco smoke on murine Th2 cell-mediated nasal eosinophilic inflammation. Asia Pac. Allergy 2020, 10, e18. [Google Scholar] [CrossRef]
- Neumann, C.; Scheffold, A.; Rutz, S. Functions and regulation of T cell-derived interleukin-10. Semin. Immunol. 2019, 44, 101344. [Google Scholar] [CrossRef]
- Kawashima, K.; Fujii, T.; Moriwaki, Y.; Misawa, H.; Horiguchi, K. Reconciling neuronally and nonneuronally derived acetylcholine in the regulation of immune function. Ann. N. Y. Acad. Sci. 2012, 1261, 7–17. [Google Scholar] [CrossRef]
- Razani-Boroujerdi, S.; Boyd, R.T.; Dávila-García, M.I.; Nandi, J.S.; Mishra, N.C.; Singh, S.P.; Pena-Philippides, J.C.; Langley, R.; Sopori, M.L. T cells express alpha7-nicotinic acetylcholine receptor subunits that require a functional TCR and leukocyte-specific protein tyrosine kinase for nicotine-induced Ca2+ response. J. Immunol. 2007, 179, 2889–2898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, R.; Ushiyama, N.; Fujii, T.; Kawashima, K. Nicotine-induced Ca2+ signaling and down-regulation of nicotinic acetylcholine receptor subunit expression in the CEM human leukemic T-cell line. Life Sci. 2003, 72, 2155–2158. [Google Scholar] [CrossRef]
- Lee, C.H.; Chang, Y.C.; Chen, C.S.; Tu, S.H.; Wang, Y.J.; Chen, L.C.; Chang, Y.J.; Wei, P.L.; Chang, H.W.; Chang, C.H.; et al. Crosstalk between nicotine and estrogen-induced estrogen receptor activation induces α9-nicotinic acetylcholine receptor expression in human breast cancer cells. Breast Cancer Res. Treat. 2011, 129, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Mashimo, M.; Moriwaki, Y.; Misawa, H.; Ono, S.; Horiguchi, K.; Kawashima, K. Expression and Function of the Cholinergic System in Immune Cells. Front. Immunol. 2017, 8, 1085. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.D.; Liao, Y.C.; Ho, Y.S.; Chen, L.C.; Chang, H.W.; Cheng, T.C.; Liu, D.; Lee, W.R.; Shen, S.C.; Wu, C.H.; et al. The α9 Nicotinic Acetylcholine Receptor Mediates Nicotine-Induced PD-L1 Expression and Regulates Melanoma Cell Proliferation and Migration. Cancers 2019, 11, 1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simard, A.R.; Gan, Y.; St-Pierre, S.; Kousari, A.; Patel, V.; Whiteaker, P.; Morley, B.J.; Lukas, R.J.; Shi, F.D. Differential modulation of EAE by α9*- and β2*-nicotinic acetylcholine receptors. Immunol. Cell Biol. 2013, 91, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, M.; Whiteaker, P.; Shi, F.D.; Morley, B.J.; Lukas, R.J. Attenuation in Nicotinic Acetylcholine Receptor α9 and α10 Subunit Double Knock-Out Mice of Experimental Autoimmune Encephalomyelitis. Biomolecules 2019, 9, 827. [Google Scholar] [CrossRef] [Green Version]
- Tai, P.; Wang, J.; Jin, H.; Song, X.; Yan, J.; Kang, Y.; Zhao, L.; An, X.; Du, X.; Chen, X.; et al. Induction of regulatory T cells by physiological level estrogen. J. Cell. Physiol. 2008, 214, 456–464. [Google Scholar] [CrossRef]
- Polanczyk, M.J.; Hopke, C.; Vandenbark, A.A.; Offner, H. Treg suppressive activity involves estrogen-dependent expression of programmed death-1 (PD-1). Int. Immunol. 2007, 19, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Valor, L.M.; Castillo, M.; Ortiz, J.A.; Criado, M. Transcriptional regulation by activation and repression elements located at the 5’-noncoding region of the human alpha9 nicotinic receptor subunit gene. J. Biol. Chem. 2003, 278, 37249–37255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, T.; Nakata, Y.; Nagamachi, A.; Yamasaki, N.; Kanai, A.; Sera, Y.; Sasaki, M.; Matsui, H.; Honda, Z.; Oda, H.; et al. Propagation of trimethylated H3K27 regulated by polycomb protein EED is required for embryogenesis, hematopoietic maintenance, and tumor suppression. Proc. Natl. Acad. Sci. USA 2016, 113, 10370–10375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaminuma, O.; Kitamura, N.; Nishito, Y.; Nemoto, S.; Tatsumi, H.; Mori, A.; Hiroi, T. Downregulation of NFAT3 Due to Lack of T-Box Transcription Factor TBX5 Is Crucial for Cytokine Expression in T Cells. J. Immunol. 2018, 200, 92–100. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakata, Y.; Miura, K.; Yamasaki, N.; Ogata, S.; Miura, S.; Hosomi, N.; Kaminuma, O. Expression and Function of Nicotinic Acetylcholine Receptors in Induced Regulatory T Cells. Int. J. Mol. Sci. 2022, 23, 1779. https://doi.org/10.3390/ijms23031779
Nakata Y, Miura K, Yamasaki N, Ogata S, Miura S, Hosomi N, Kaminuma O. Expression and Function of Nicotinic Acetylcholine Receptors in Induced Regulatory T Cells. International Journal of Molecular Sciences. 2022; 23(3):1779. https://doi.org/10.3390/ijms23031779
Chicago/Turabian StyleNakata, Yuichiro, Kento Miura, Norimasa Yamasaki, Sawako Ogata, Shuka Miura, Naohisa Hosomi, and Osamu Kaminuma. 2022. "Expression and Function of Nicotinic Acetylcholine Receptors in Induced Regulatory T Cells" International Journal of Molecular Sciences 23, no. 3: 1779. https://doi.org/10.3390/ijms23031779