
Conversion of Human Fibroblasts into Induced Neural Stem Cells by Small Molecules

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
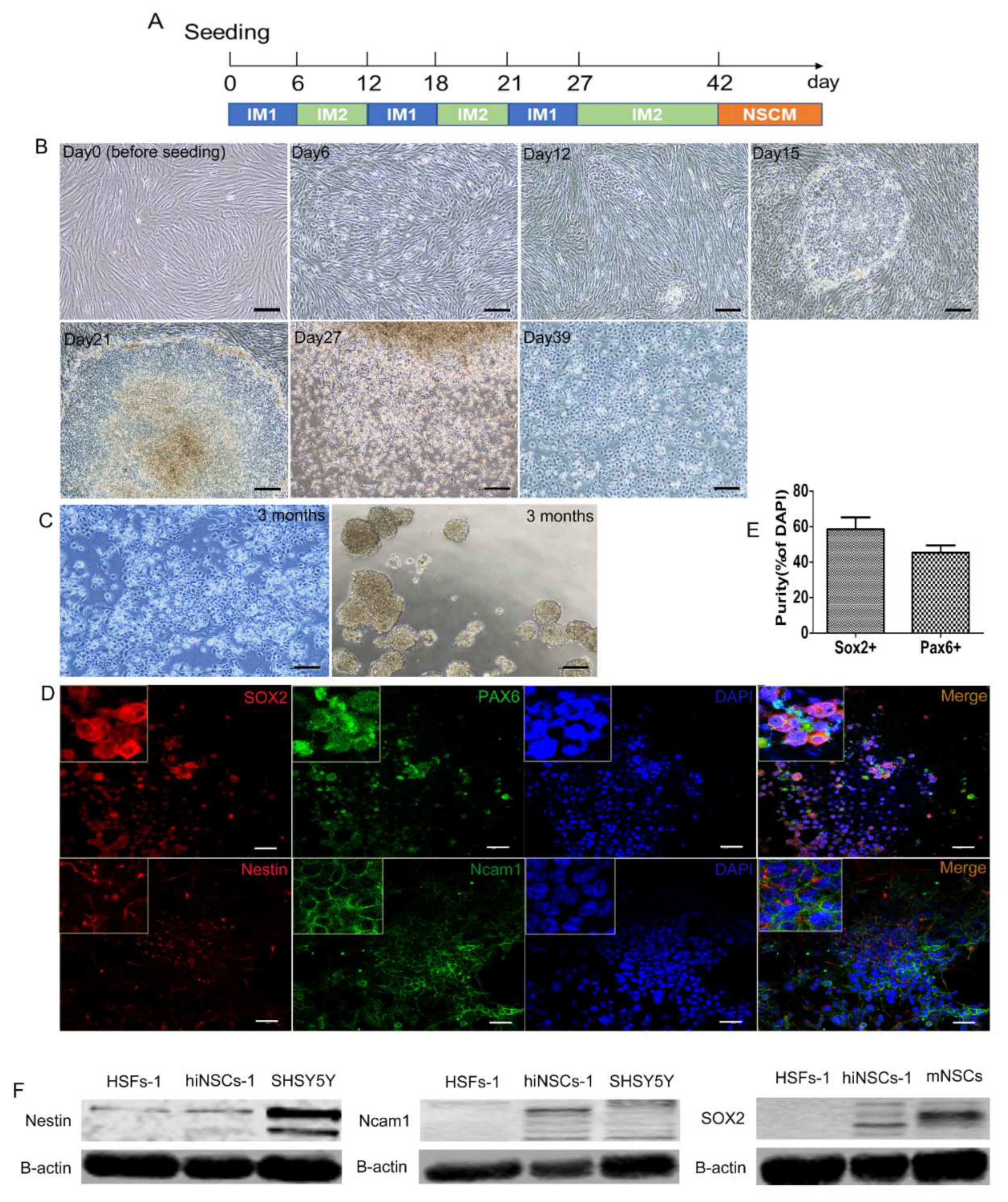
2.1. Generation of hiNSCs from Human Fibroblasts by Small Molecules
2.2. hiNSCs-1 Express Multiple Up-Regulated NSCs Makers and Down-Regulated Fibroblast-Specific Genes
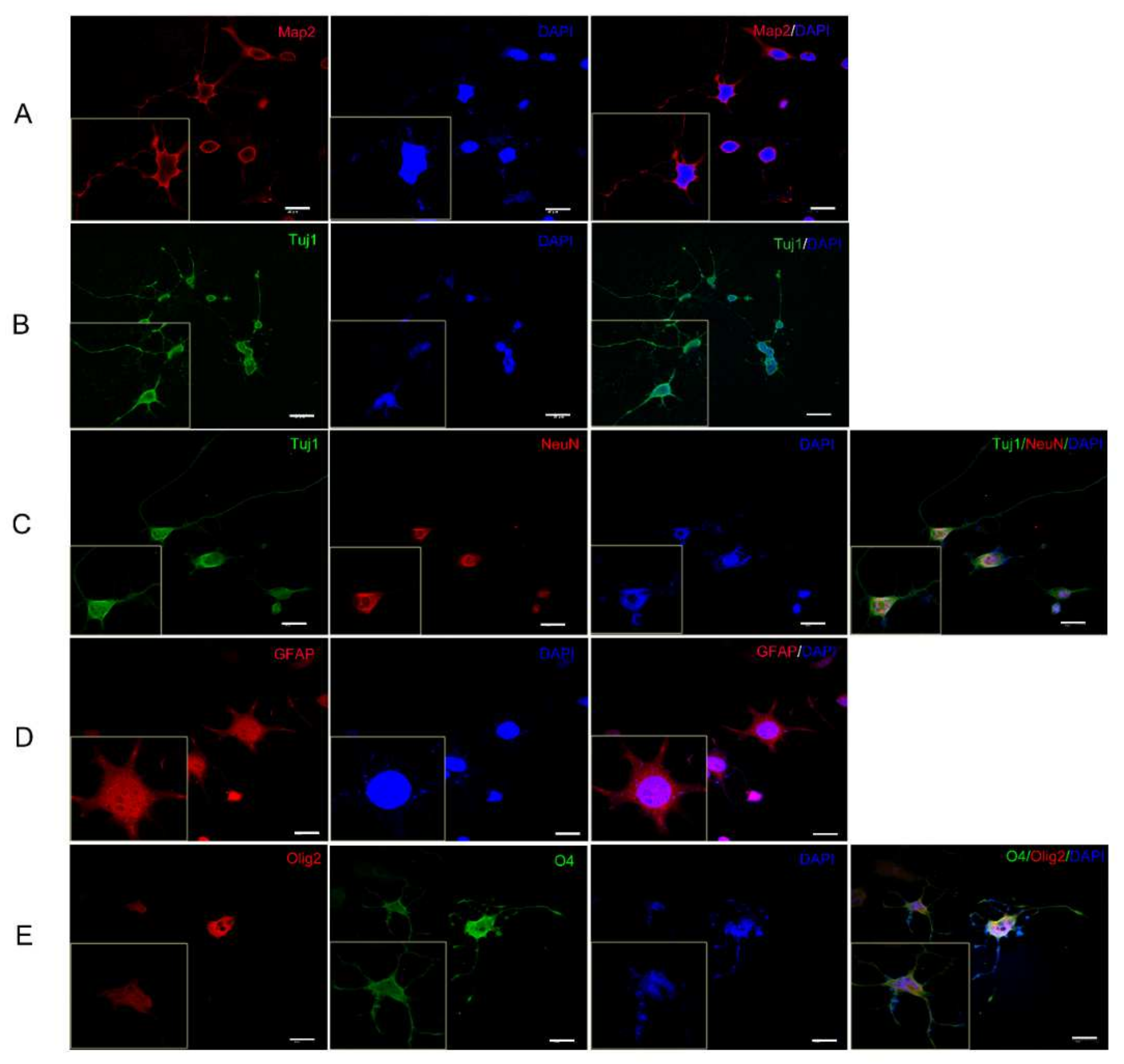
2.3. hiNSCs-1 Can Differentiate into Astrocytes, Oligodendrocytes, and Neurons In Vitro
2.4. Differentiated Neurons from hiNSCs-1 Possess Mature Electrophysiological Properties as Bona Fide Neurons
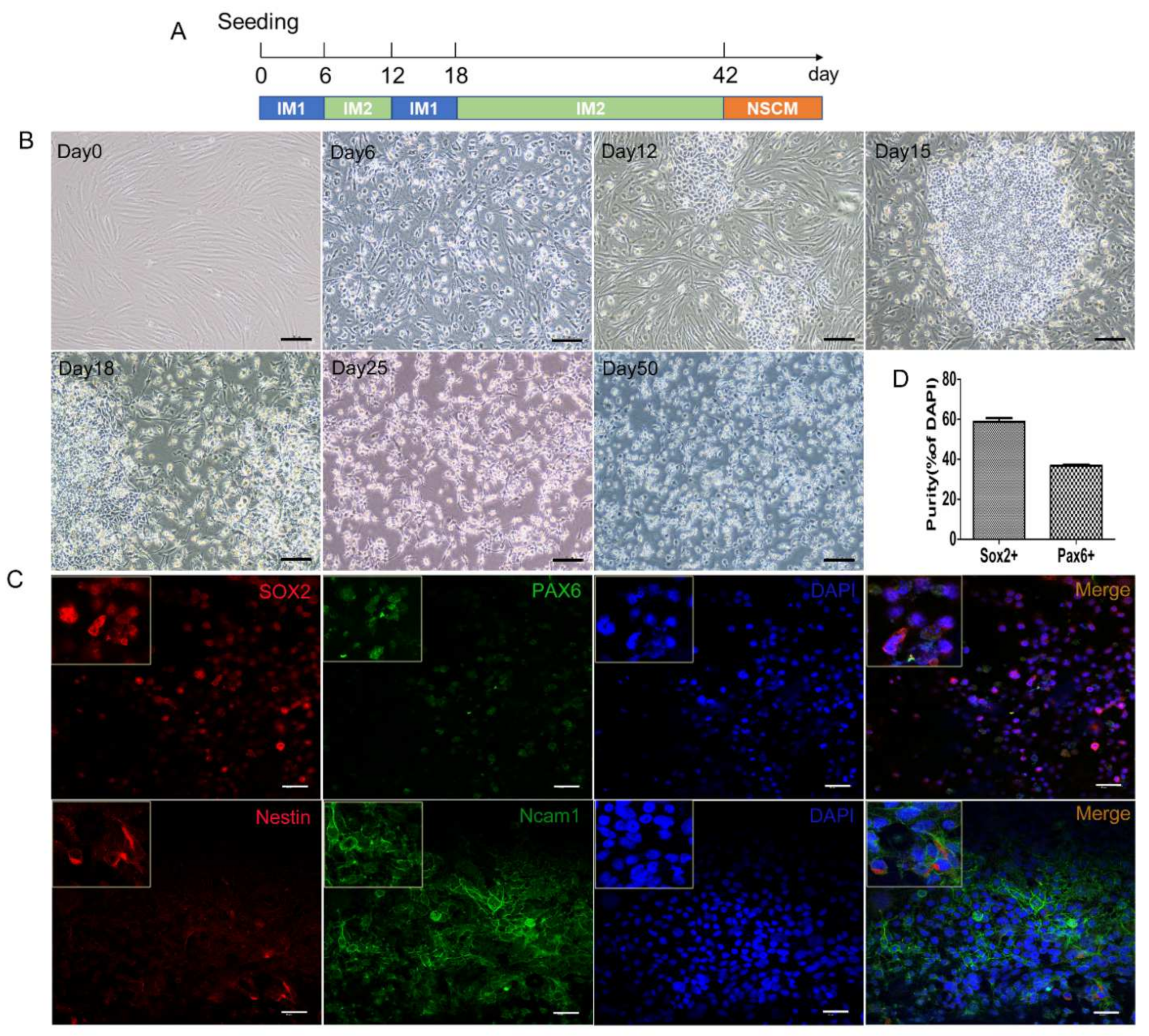
2.5. Generation of hiNSCs from Another Fibroblast Cell Line
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Generation of iNSCs from Human Fibroblasts
4.3. In Vitro Differentiation of iNSCs
4.4. Immunocytochemistry Staining
4.5. Western Blot
4.6. Real-Time qPCR
4.7. RNA-Sequencing
4.8. Whole-Cell Patch Clamping
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGinley, L.M.; Kashlan, O.N.; Bruno, E.S.; Chen, K.S.; Hayes, J.M.; Kashlan, S.R.; Raykin, J.; Johe, K.; Murphy, G.G.; Feldman, E.L. Human neural stem cell transplantation improves cognition in a murine model of alzheimer’s disease. Sci. Rep. 2018, 8, 14776. [Google Scholar] [CrossRef] [PubMed]
- Yasuhara, T.; Kameda, M.; Sasaki, T.; Tajiri, N.; Date, I. Cell therapy for parkinson’s disease. Cell Transpl. 2017, 26, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Kokaia, Z.; Darsalia, V. Human neural stem cells for ischemic stroke treatment. Results Probl. Cell Differ. 2018, 66, 249–263. [Google Scholar] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Wei, W.; Zhu, S.; Zhu, J.; Shi, Y.; Lin, T.; Hao, E.; Hayek, A.; Deng, H.; Ding, S. Generation of rat and human induced pluripotent stem cells by combining genetic reprogramming and chemical inhibitors. Cell Stem Cell 2009, 4, 16–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellin, M.; Marchetto, M.C.; Gage, F.H.; Mummery, C.L. Induced pluripotent stem cells: The new patient? Nat. Rev. Mol. Cell Biol. 2012, 13, 713–726. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.S.; Malik, N. Assessing ipsc reprogramming methods for their suitability in translational medicine. J. Cell Biochem. 2012, 113, 3061–3068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahbazi, E.; Mirakhori, F.; Ezzatizadeh, V.; Baharvand, H. Reprogramming of somatic cells to induced neural stem cells. Methods 2018, 133, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Han, D.W.; Tapia, N.; Hermann, A.; Hemmer, K.; Hoing, S.; Arauzo-Bravo, M.J.; Zaehres, H.; Wu, G.M.; Frank, S.; Moritz, S.; et al. Direct reprogramming of fibroblasts into neural stem cells by defined factors. Cell Stem Cell 2012, 10, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, D.C.; Liu, X.N.; Zhang, M.; Zou, M.; Deng, Q.Q.; Sun, D.Y.; Bian, X.T.; Cai, Y.L.; Guo, Y.A.; Liu, S.T.; et al. Direct reprogramming of fibroblasts into neural stem cells by single non-neural progenitor transcription factor ptf1a. Nat. Commun. 2018, 9, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, T.; Sugawara, T.; Fukuda, A.; Tamoto, R.; Kawasaki, T.; Umezawa, A.; Akutsu, H. Generation of primitive neural stem cells from human fibroblasts using a defined set of factors. Biol. Open 2015, 4, 1595–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirakhori, F.; Zeynali, B.; Rassouli, H.; Salekdeh, G.H.; Baharvand, H. Direct conversion of human fibroblasts into dopaminergic neural progenitor-like cells using tat-mediated protein transduction of recombinant factors. Biochem. Biophys. Res. Commun. 2015, 459, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhang, L.L.; An, J.; Zhang, Q.; Liu, C.C.; He, B.R.; Hao, D.J. Microrna-mediated reprogramming of somatic cells into neural stem cells or neurons. Mol. Neurobiol. 2017, 54, 1587–1600. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Choi, K.A.; Bang, P.J.; Hyeon, S.; Kwon, S.; Moon, J.H.; Hwang, I.; Kim, Y.I.; Kim, Y.S.; Yoon, B.S.; et al. A combination of small molecules directly reprograms mouse fibroblasts into neural stem cells. Biochem. Biophys. Res. Commun. 2016, 476, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Hu, W.X.; Qiu, B.L.; Zhao, J.; Yu, Y.C.; Guan, W.Q.; Wang, M.; Yang, W.Z.; Pei, G. Generation of neural progenitor cells by chemical cocktails and hypoxia. Cell Res. 2014, 24, 665–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.C.; Lim, Y.; Duffieldl, M.D.; Li, H.; Liu, J.; Manaph, N.P.A.; Yang, M.; Keating, D.J.; Zhou, X.F. Direct reprogramming of mouse fibroblasts to neural stem cells by small molecules. Stem Cells Int. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Pavathuparambil Abdul Manaph, N.; Al-Hawwas, M.; Zhou, X.F.; Liao, H. Small molecules for neural stem cell induction. Stem Cells Dev. 2018, 27, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Bobrovskaya, L.; Zhou, X.F. Cell therapy for neurological disorders: The perspective of promising cells. Biology 2021, 10, 1142. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Rychkov, G.; Al-Hawwas, M.; Manaph, N.P.A.; Zhou, F.; Bobrovskaya, L.; Liao, H.; Zhou, X.F. Conversion of human urine-derived cells into neuron-like cells by small molecules. Mol. Biol. Rep. 2020, 47, 2713–2722. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Rychkov, G.; Hurtado, P.; Luo, H.-Y.; Zhang, T.; Bobrovskaya, L.; Zhou, X.-F. Conversion of Human Fibroblasts into Induced Neural Stem Cells by Small Molecules. Int. J. Mol. Sci. 2022, 23, 1740. https://doi.org/10.3390/ijms23031740
Liu D, Rychkov G, Hurtado P, Luo H-Y, Zhang T, Bobrovskaya L, Zhou X-F. Conversion of Human Fibroblasts into Induced Neural Stem Cells by Small Molecules. International Journal of Molecular Sciences. 2022; 23(3):1740. https://doi.org/10.3390/ijms23031740
Chicago/Turabian StyleLiu, Donghui, Grigori Rychkov, Plinio Hurtado, Hai-Yun Luo, Tao Zhang, Larisa Bobrovskaya, and Xin-Fu Zhou. 2022. "Conversion of Human Fibroblasts into Induced Neural Stem Cells by Small Molecules" International Journal of Molecular Sciences 23, no. 3: 1740. https://doi.org/10.3390/ijms23031740