
Advanced Glycation End-Products (AGEs) of Lysine and Effects of Anti-TCR/Anti-TNF-α Antibody-Based Therapy in the LEW.1AR1-iddm Rat, an Animal Model of Human Type 1 Diabetes

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Analytical Aspects
2.2. Furosine, Nε-Carboxymethyl Lysine (CML) and Nε-Monomethyl Lysine (MML)
2.2.1. Pancreas
2.2.2. Kidney
2.2.3. Liver and Lymph Nodes
2.2.4. Spleen
2.3. Correlations of Amino Acids with Blood Glucose
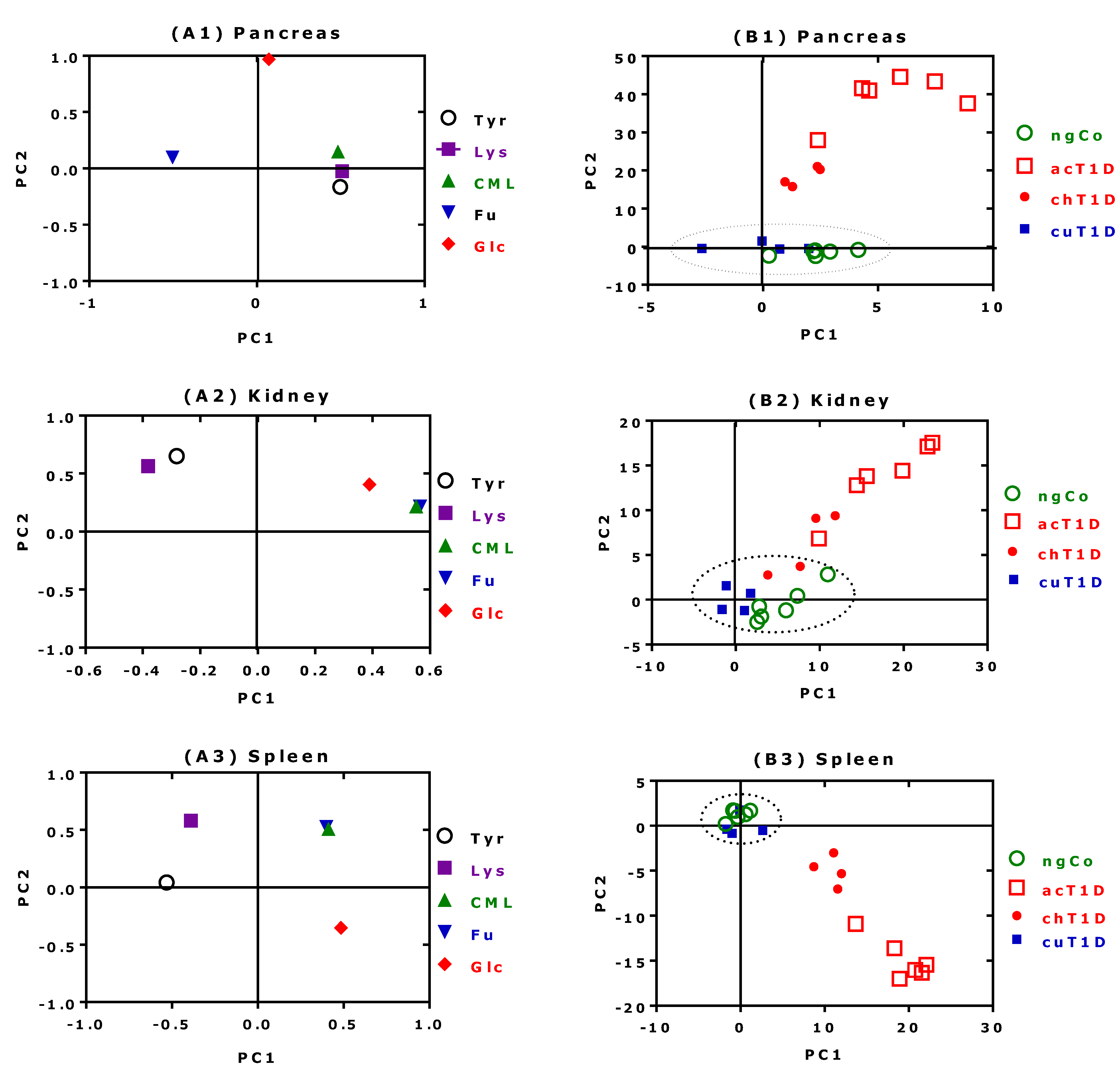
2.4. PCA Results for CML, Furosine, Lysine, Tyrosine and Glucose
2.4.1. Pancreas
2.4.2. Kidney
2.4.3. Spleen
2.5. OPLS-DA Results for Amino Acids
2.5.1. Pancreas
2.5.2. Spleen
2.5.3. Kidney
2.5.4. Liver
3. Discussion and Conclusions
4. Materials and Methods
4.1. Experimental Groups of the LEW.1AR1-iddm Model of Human T1D
4.2. Organ Homogenization and GC-MS Measurement of Tissue Free and Proteinic Amino Acids and Metabolites
4.3. Statistical Analysis and Data Presentation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tsikas, D. Post-translational modifications (PTM): Analytical approaches, signaling, physiology and pathophysiology-part I. Amino Acids 2021, 53, 485–487. [Google Scholar] [CrossRef]
- Falnes, P.Ø.; Jakobsson, M.E.; Davydova, E.; Ho, A.; Małecki, J. Protein lysine methylation by seven-beta-strand methyltransferases. Biochem. J. 2016, 473, 1995–2009. [Google Scholar] [CrossRef]
- Hornbeck, P.V.; Kornhauser, J.M.; Tkachev, S.; Zhang, B.; Skrzypek, E.; Murray, B.; Latham, V.; Sullivan, M. PhosphoSitePlus: A comprehensive resource for investigating the structure and function of experimentally determined post-translational modifications in man and mouse. Nucleic Acids Res. 2016, 40, D261–D270. [Google Scholar] [CrossRef] [Green Version]
- Tsikas, D.; Sandmann, J.; Savva, A.; Luessen, P.; Böger, R.H.; Gutzki, F.M.; Mayer, B.; Frölich, J.C. Assessment of nitric oxide synthase activity in vitro and in vivo by gas chromatography-mass spectrometry. J. Chromatogr. B Biomed. Sci. Appl. 2000, 742, 143–153. [Google Scholar] [CrossRef]
- Baskal, S.; Bollenbach, A.; Mels, C.; Kruger, R.; Tsikas, D. Development, validation of a GC-MS method for the simultaneous measurement of amino acids, their PTM metabolites and AGEs in human urine, and application to the bi-ethnic ASOS study with special emphasis to lysine. Amino Acids 2021, in press. [CrossRef]
- Baskal, S.; Büttner, P.; Werner, S.; Besler, C.; Lurz, P.; Thiele, H.; Tsikas, D. Profile of urinary amino acids and their post-translational modifications (PTM) including advanced glycation end-products (AGEs) of lysine, arginine and cysteine in lean and obese ZSF1 rats. Amino Acids 2021, in press. [CrossRef]
- Schmidt, A.M.; Hofmann, M.; Taguchi, A.; Yan, S.D.; Stern, D.M. RAGE: A multiligand receptor contributing to the cellular response in diabetic vasculopathy and inflammation. Semin. Thromb. Hemost. 2000, 26, 485–493. [Google Scholar] [CrossRef]
- Xue, J.; Rai, V.; Singer, D.; Chabierski, S.; Xie, J.; Reverdatto, S.; Burz, D.S.; Schmidt, A.M.; Hoffmann, R.; Shekhtman, A. Advanced glycation end product recognition by the receptor for AGEs. Structure 2011, 19, 722–732. [Google Scholar] [CrossRef] [Green Version]
- Oimomi, M.; Hatanaka, H.; Ishikawa, K.; Kubota, S.; Yoshimura, Y.; Baba, S. Increased fructose-lysine of nail protein in diabetic patients. Klin. Wochenschr. 1984, 62, 477–478. [Google Scholar] [CrossRef]
- Gugliucci, A. Formation of Fructose-Mediated Advanced Glycation End Products and Their Roles in Metabolic and Inflammatory Diseases. Adv. Nutr. 2017, 8, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Palanissami, G.; Paul, S.F.D. RAGE and Its Ligands: Molecular Interplay Between Glycation, Inflammation, and Hallmarks of Cancer-a Review. Horm. Cancer 2018, 9, 295–325. [Google Scholar] [CrossRef]
- Steenbeke, M.; De Bruyne, S.; De Buyzere, M.; Lapauw, B.; Speeckaert, R.; Petrovic, M.; Delanghe, J.R.; Speeckaert, M.M. The role of soluble receptor for advanced glycation end-products (sRAGE) in the general population and patients with diabetes mellitus with a focus on renal function and overall outcome. Crit. Rev. Clin. Lab. Sci. 2021, 58, 113–130. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Q.; Wei, D.; Chang, X. Association Between Soluble Receptor for Advanced Glycation End Product and Endogenous Secretory Soluble Receptor for Advanced Glycation End Product Levels and Carotid Atherosclerosis in Diabetes: A Systematic Review and Meta-Analysis. Can. J. Diabetes 2021, 45, 634–640. [Google Scholar] [CrossRef]
- Wu, X.Q.; Zhang, D.D.; Wang, Y.N.; Tan, Y.Q.; Yu, X.Y.; Zhao, Y.Y. AGE/RAGE in diabetic kidney disease and ageing kidney. Free Radic. Biol. Med. 2021, 171, 260–271. [Google Scholar] [CrossRef]
- Bierhaus, A.; Schiekofer, S.; Schwaninger, M.; Andrassy, M.; Humpert, P.M.; Chen, J.; Hong, M.; Luther, T.; Henle, T.; Klöting, I.; et al. Diabetes-associated sustained activation of the transcription factor nuclear factor-kappaB. Diabetes 2001, 50, 2792–2808. [Google Scholar] [CrossRef] [Green Version]
- Mu, L.; Zhang, Y.; Sun, B.; Wang, J.; Xie, X.; Li, N.; Zhang, J.; Kong, Q.; Liu, Y.; Han, Z.; et al. Activation of the receptor for advanced glycation end products (RAGE) exacerbates experimental autoimmune myasthenia gravis symptoms. Clin. Immunol. 2011, 141, 36–48. [Google Scholar] [CrossRef]
- Chen, Y.; Shi, S.; Yan, D.; Colgan, J.; Zhang, H.; Luban, J.; Schmidt, A.M.; Stern, D.; Herold, K.C. Blockade of late stages of autoimmune diabetes by inhibition of the receptor for advanced glycation end products. J. Immunol. 2004, 173, 1399–1405. [Google Scholar] [CrossRef] [Green Version]
- Willemsen, S.; Hartog, J.W.L.; van Veldhuisen, D.J.; van der Meer, P.; Roze, J.F.; Jaarsma, T.; Schalkwijk, C.; van der Horst, I.C.C.; Hillege, H.L.; Voors, A.A. The role of advanced glycation end-products and their receptor on outcome in heart failure patients with preserved and reduced ejection fraction. Am. Heart J. 2012, 164, 742–749.e3. [Google Scholar] [CrossRef]
- Tian, Y.; Charles, E.J.; Yan, Z.; Wu, D.; French, B.A.; Kron, I.L.; Yang, Z. The myocardial infarct-exacerbating effect of cell-free DNA is mediated by the high-mobility group box 1-receptor for advanced glycation end products-Toll-like receptor 9 pathway. J. Thorac. Cardiovasc. Surg. 2019, 157, 2256–2269.e3. [Google Scholar] [CrossRef] [Green Version]
- Saleh, I.; Maritska, Z.; Parisa, N.; Hidayat, R. Inhibition of Receptor for Advanced Glycation End Products as New Promising Strategy Treatment in Diabetic Retinopathy. Open Access Maced. J. Med. Sci. 2019, 7, 3921–3924. [Google Scholar] [CrossRef] [Green Version]
- Angelopoulou, E.; Paudel, Y.N.; Piperi, C. Unraveling the Role of Receptor for Advanced Glycation End Products (RAGE) and Its Ligands in Myasthenia Gravis. ACS Chem. Neurosci. 2020, 11, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Lenzen, S.; Tiedge, N.; Elsner, M.; Lortz, S.; Weiss, H.; Jörns, A.; Klöppel, G.; Wedekind, D.; Prokop, C.M.; Hedrich, H.J. The LEW.1AR1/Ztm-iddm rat: A new model of spontaneous insulin-dependent diabetes mellitus. Diabetologia 2001, 44, 1189–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jörns, A.; Kubat, B.; Tiedge, M.; Wedekind, D.; Hedrich, H.J.; Klöppel, G.; Lenzen, S. Pathology of the pancreas and other organs in the diabetic LEW.1AR1/Ztm- iddm rat, a new model of spontaneous insulin-dependent diabetes mellitus. Virchows Arch. 2004, 444, 183–189. [Google Scholar] [CrossRef]
- Jörns, A.; Günther, A.; Hedrich, H.J.; Wedekind, D.; Tiedge, M.; Lenzen, S. Immune cell infiltration, cytokine expression, and beta-cell apoptosis during the development of type 1 diabetes in the spontaneously diabetic LEW.1AR1/Ztm-iddm rat. Diabetes 2005, 54, 2041–2052. [Google Scholar] [CrossRef] [Green Version]
- Jörns, A.; Arndt, T.; Meyer zu Vilsendorf, A.; Klempnauer, J.; Wedekind, D.; Hedrich, H.J.; Marselli, L.; Marchetti, P.; Harada, N.; Nakaya, Y.; et al. Islet infiltration, cytokine expression and beta cell death in NOD mouse, BB rat, Komeda rat, LEW.1AR1-iddm rat and humans with type 1 diabetes. Diabetologia 2014, 57, 512–521. [Google Scholar] [CrossRef]
- Jörns, A.; Ertekin, Ü.G.; Arndt, T.; Terbish, T.; Wedekind, D.; Lenzen, S. TNF-α Antibody Therapy in Combination With the T-Cell-Specific Antibody Anti-TCR Reverses the Diabetic Metabolic State in the LEW.1AR1-iddm Rat. Diabetes 2015, 64, 2880–2891. [Google Scholar] [CrossRef] [Green Version]
- Tsikas, D.; Hanff, E.; Bollenbach, A.; Kruger, R.; Pham, V.V.; Chobanyan-Jürgens, K.; Wedekind, D.; Arndt, T.; Jörns, A.; Berbée, J.F.P.; et al. Results, meta-analysis and a first evaluation of UNOxR, the urinary nitrate-to-nitrite molar ratio, as a measure of nitrite reabsorption in experimental and clinical settings. Amino Acids 2018, 50, 799–821. [Google Scholar] [CrossRef] [Green Version]
- Jörns, A.; Ishikawa, D.; Teraoku, H.; Yoshimoto, T.; Wedekind, D.; Lenzen, S. Remission of autoimmune diabetes by anti-TCR combination therapies with anti-IL-17A or/and anti-IL-6 in the IDDM rat model of type 1 diabetes. BMC Med. 2020, 18, 33. [Google Scholar] [CrossRef]
- Bollenbach, A.; Tsikas, D.; Lenzen, S.; Jörns, A. Asymmetric dimethylation and citrullination in the LEW.1AR1-iddm rat, an animal model of human type 1 diabetes, and effects of anti-TCR/anti-TNF-α antibody-based therapy. Amino Acids 2020, 52, 103–110. [Google Scholar] [CrossRef]
- Felig, P.; Wahren, J.; Räf, L. Evidence of inter-organ amino-acid transport by blood cells in humans. Proc. Natl. Acad. Sci. USA 1973, 70, 1775–1779. [Google Scholar] [CrossRef] [Green Version]
- Griffith, O.W.; Meister, A. Glutathione: Interorgan translocation, turnover, and metabolism. Proc. Natl. Acad. Sci. USA 1979, 76, 5606–5610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorn, B.; Dunstan, R.H.; Macdonald, M.M.; Borges, N.; Roberts, T.K. Evidence that human and equine erythrocytes could have significant roles in the transport and delivery of amino acids to organs and tissues. Amino Acids 2020, 52, 711–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, S.S.; Forbes, J.M.; Borg, D.J. Receptor for Advanced Glycation End Products (RAGE) in Type 1 Diabetes Pathogenesis. Curr. Diabetes Rep. 2016, 16, 100. [Google Scholar] [CrossRef] [PubMed]
- Kehm, R.; Rückriemen, J.; Weber, D.; Deubel, S.; Grune, T.; Höhn, A. Endogenous advanced glycation end products in pancreatic islets after short-term carbohydrate intervention in obese, diabetes-prone mice. Nutr. Diabetes 2019, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Guilbaud, A.; Howsam, M.; Niquet-Léridon, C.; Delguste, F.; Boulanger, E.; Tessier, F.J. The LepR db/db mice model for studying glycation in the context of diabetes. Diabetes Metab. Res. Rev. 2019, 35, e3103. [Google Scholar] [CrossRef] [PubMed]
- Borg, D.J.; Yap, F.Y.T.; Keshvari, S.; Simmons, D.G.; Gallo, L.A.; Fotheringham, A.K.; Zhuang, A.; Slattery, R.M.; Hasnain, S.Z.; Coughlan, M.T.; et al. Perinatal exposure to high dietary advanced glycation end products in transgenic NOD8.3 mice leads to pancreatic beta cell dysfunction. Islets 2018, 10, 10–24. [Google Scholar] [CrossRef]
- Tian, Y.; Pan, D.; Chordia, M.D.; French, B.A.; Kron, I.L.; Yang, Z. The spleen contributes importantly to myocardial infarct exacerbation during post-ischemic reperfusion in mice via signaling between cardiac HMGB1 and splenic RAGE. Basic Res. Cardiol. 2016, 111, 62. [Google Scholar] [CrossRef] [Green Version]
- Dhawan, R.; Chaney, M.A. Commentary: RAGE against the spleen. J. Thorac. Cardiovasc. Surg. 2019, 157, 2272–2273. [Google Scholar] [CrossRef] [Green Version]
- Gagnerault, M.C.; Luan, J.J.; Lotton, C.; Lepault, F. Pancreatic lymph nodes are required for priming of beta cell reactive T cells in NOD mice. J. Exp. Med. 2002, 196, 369–377. [Google Scholar] [CrossRef]
- Ko, H.J.; Chee, J.; Sutherland, R.M.; Thomas, H.E.; Zhan, Y.; Krishnamurthy, B.; Kay, T.W.; Lew, A.M. Functional cytotoxic T lymphocytes against IGRP206-214 predict diabetes in the non-obese diabetic mouse. Immunol. Cell Biol. 2014, 92, 640–644. [Google Scholar] [CrossRef]
- Lewis, S.M.; Williams, A.; Eisenbarth, S.C. Structure and function of the immune system in the spleen. Sci. Immunol. 2019, 1, eaau6085. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Chandel, S.; Dey, D.; Ghosh, A.; Roy, S.; Ravichandiran, V.; Ghosh, D. Epigenetic modification and therapeutic targets of diabetes mellitus. Biosci. Rep. 2020, 40, BSR20202160. [Google Scholar] [CrossRef] [PubMed]
- Leslie, R.D.; Beyan, H.; Sawtell, P.; Boehm, B.O.; Spector, T.D.; Snieder, H. Level of an advanced glycated end product is genetically determined: A study of normal twins. Diabetes 2003, 52, 2441–2444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snieder, H.; Sawtell, P.A.; Ross, L.; Walker, J.; Spector, T.D.; Leslie, R.D. HbA1c levels are genetically determined even in type 1 diabetes: Evidence from healthy and diabetic twins. Diabetes 2001, 50, 2858–2863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabbani, N.; Thornalley, P.J. Protein glycation—Biomarkers of metabolic dysfunction and early-stage decline in health in the era of precision medicine. Redox Biol. 2021, 42, 101920. [Google Scholar] [CrossRef] [PubMed]
- Roep, B.O.; Thomaidou, S.; van Tienhoven, R.; Zaldumbide, A. Type 1 diabetes mellitus as a disease of the β-cell (do not blame the immune system?). Nat. Rev. Endocrinol. 2021, 17, 150–161. [Google Scholar] [CrossRef]
- Myint, K.M.; Yamamoto, Y.; Doi, T.; Kato, I.; Harashima, A.; Yonekura, H.; Watanabe, T.; Shinohara, H.; Takeuchi, M.; Tsuneyama, K.; et al. RAGE control of diabetic nephropathy in a mouse model: Effects of RAGE gene disruption and administration of low-molecular weight heparin. Diabetes 2006, 55, 2510–2522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baskal, S.; Post, A.; Kremer, D.; Bollenbach, A.; Bakker, S.J.L.; Tsikas, D. Urinary excretion of amino acids and their advanced glycation end-products (AGEs) in adult kidney transplant recipients with emphasis on lysine: Furosine excretion is associated with cardiovascular and all-cause mortality. Amino Acids 2021, 53, 1679–1693. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Free | Proteinic | Free | Proteinic |
|---|---|---|---|---|
| Furosine | Carboxymethyllysine | |||
| PANCREAS | ||||
| ngCo | 0.0412 ± 0.0094 | 0.0206 ± 0.0089 | 0.6033 ± 0.0092 | 0.1200 ± 0.0656 |
| acT1D | 0.0410 ± 0.0043 | 0.0743 ± 0.0183 | 0.7700 ± 0.0856 | 0.1250 ± 0.0296 |
| chT1D | n.d. | 0.0883 ± 0.0102 | 0.2800 ± 0.0212 | 0.0550 ± 0.0119 |
| cuT1D | n.d. | 0.0825 ± 0.0168 | 0.2525 ± 0.0364 | 0.0625 ± 0.0225 |
| n.s. | p = 0.0289 | p = 0.0023 | n.s. | |
| KIDNEY | ||||
| ngCo | 0.2022 ± 0.0489 | 0.5583 ± 0.0260 | 0.885 ± 0.1356 | 0.3283 ± 0.0456 |
| acT1D | 0.0708 ± 0.0118 | 0.4950 ± 0.0796 | 0.5483 ± 0.1269 | 0.1950 ± 0.0273 |
| chT1D | 0.0863 ± 0.0209 | 0.5175 ± 0.0984 | 0.4175 ± 0.0978 | 0.215 ± 0.0444 |
| cuT1D | 0.1410 ± 0.0146 | 0.5450 ± 0.0491 | 0.4375 ± 0.0379 | 0.1600 ± 0.0234 |
| n.s. | p = 0.0076 | n.s. | p = 0.0270 | |
| LIVER | ||||
| ngCo | 0.0193 ± 0.0030 | 0.1500 ± 0.0146 | 0.0752 ± 0.0083 | 0.1288 ± 0.0146 |
| acT1D | 0.0165 ± 0.0010 | 0.1800 ± 0.0159 | 0.0727 ± 0.0073 | 0.1362 ± 0.0112 |
| n.s. | n.s. | n.s. | n.s. | |
| SPLEEN | ||||
| ngCo | 0.0592 ± 0.0089 | 0.1210 ± 0.0123 | 0.1617 ± 0.0260 | 0.2250 ± 0.0272 |
| acT1D | 0.0318 ± 0.0036 | 0.1033 ± 0.0194 | 0.1483 ± 0.0162 | 0.1367 ± 0.0158 |
| chT1D | n.d. | 0.2800 ± 0.0235 | 0.1800 ± 0.0227 | 0.2275 ± 0.0229 |
| cuT1D | n.d. | 0.1600 ± 0.0235 | 0.1025 ± 0.0165 | 0.2250 ± 0.0272 |
| p = 0.0368 | p = 0.0114 | n.s. | n.s. | |
| LYMPH NODE | ||||
| ngCo | 0.0313 ± 0.0107 | 0.0703 ± 0.0432 | 0.1010 ± 0.0149 | 0.0808 ± 0.0390 |
| acT1D | 0.0140 ± 0.0026 | 0.0252 ± 0.0150 | 0.0892 ± 0.0102 | 0.0276 ± 0.0086 |
| n.s. | n.s. | n.s. | n.s. | |
| Group | Nε-Monomethyllysine (MML) | |
|---|---|---|
| Free | Proteinic | |
| PANCREAS | ||
| ngCo | 0.0897 ± 0.0215 | 0.0385 ± 0.0108 |
| acT1D | 0.1183 ± 0.0168 | 0.1370 ± 0.0131 |
| chT1D | 0.0755 ± 0.0114 | 0.1295 ± 0.0231 |
| cuT1D | 0.1038 ± 0.0118 | 0.1538 ± 0.0303 |
| p = 0.0014 | n.s. | |
| KIDNEY | ||
| ngCo | 0.1123 ± 0.0322 | 0.1733 ± 0.0353 |
| acT1D | 0.0988 ± 0.0146 | 0.1238 ± 0.0159 |
| chT1D | 0.0803 ± 0.0109 | 0.1715 ± 0.0404 |
| cuT1D | 0.0755 ± 0.0029 | 0.1263 ± 0.0175 |
| n.s. | n.s. | |
| LIVER | ||
| ngCo | 0.0413 ± 0.0026 | 0.0630 ± 0.0039 |
| acT1D | 0.0323 ± 0.0037 | 0.0638 ± 0.0075 |
| n.s. | n.s. | |
| SPLEEN | ||
| ngCo | 0.1357 ± 0.0150 | 0.2867 ± 0.0320 |
| acT1D | 0.1058 ± 0.0077 | 0.2500 ± 0.0171 |
| chT1D | 0.1198 ± 0.0127 | 0.3825 ± 0.0364 |
| cuT1D | 0.1033 ± 0.024 | 0.3925 ± 0.0325 |
| n.s. | p = 0.0191 | |
| LYMPH | ||
| ngCo | 0.0858 ± 0.0198 | 0.3633 ± 0.1016 |
| acT1D | 0.059 ± 0.0083 | 0.1983 ± 0.0736 |
| n.s. | n.s. | |
| Phe | Tyr | Lys | Arg | Trp | Ala | Thr | Gly | Val | Ser | Leu/ Ile a | Asp/ Asn a | Pro | Met | Glu/ Gln a | Orn/ Cit a | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PANCREAS | ||||||||||||||||
| ngCo | 28.5 | 15.5 | 25.6 | 35.7 | 6.0 | 12.6 | 30.2 | 5.4 | 84.6 | 12.5 | 41.7 | 36.6 | 24.6 | 3.22 | 8.75 | 0.84 |
| acT1D | 25.5 | 11.8 | 19.6 | 27.0 | 6.5 | 9.6 | 21.9 | 4.5 | 52.9 | 10.9 | 33.4 | 27.8 | 19.7 | 2.70 | 6.6 | 0.59 |
| chT1D | 25.8 | 16.8 | 28.5 | 29.2 | 5.0 | 8.6 | 47.2 | 5.0 | 63.8 | 10.9 | 36.9 | 21.9 | 27.5 | 3.32 | 8.7 | 0.98 |
| cuT1D | 21.8 | 12.8 | 35.6 | 38.6 | 3.5 | 7.7 | 25.4 | 5.2 | 85.1 | 12.1 | 36.3 | 25.2 | 25.4 | 3.3 | 8.7 | 1.38 |
| KIDNEY | ||||||||||||||||
| ngCo | 11.7 | 5.1 | 10.9 | 11.3 | 4.0 | 4.5 | 8.7 | 2.8 | 12.6 | 4.3 | 13.1 | 10.1 | 9.4 | 2.8 | 6.3 | 0.46 |
| acT1D | 8.9 | 4.4 | 7.4 | 8.7 | 3.9 | 3.5 | 5.9 | 2.6 | 8.4 | 3.1 | 9.1 | 7.1 | 7.1 | 2.5 | 5.2 | 0.33 |
| chT1D | 17.4 | 9.0 | 23.5 | 18.7 | 5.5 | 8.5 | 15.3 | 3.7 | 26.4 | 5.8 | 22.6 | 17.7 | 12.2 | 2.9 | 7.2 | 1.3 |
| cuT1D | 19.7 | 9.8 | 25.6 | 22.1 | 3.4 | 9.7 | 16.9 | 3.9 | 30.8 | 6.6 | 25.4 | 18.6 | 12.5 | 3.3 | 8.9 | 1.5 |
| LIVER | ||||||||||||||||
| ngCo | 37.2 | 18.7 | 32.2 | 64.3 | 8.5 | 8.7 | 37.6 | 5.7 | 75.2 | 15.1 | 46.3 | 70.8 | 54.0 | 5.4 | 13.7 | 1.9 |
| acT1D | 34.8 | 17.0 | 28.3 | 66.5 | 6.3 | 9.9 | 34.5 | 5.5 | 59.0 | 13.0 | 42.0 | 54.7 | 57.8 | 5.1 | 11.1 | 1.0 |
| SPLEEN | ||||||||||||||||
| ngCo | 12.6 | 4.3 | 18.2 | 11.6 | 3.3 | 9.2 | 13.7 | 3.6 | 19.0 | 6.8 | 17.1 | 8.3 | 13.7 | 2.8 | 5.8 | 2.7 |
| acT1D | 13.2 | 4.2 | 19.1 | 11.2 | 3.5 | 8.4 | 12.3 | 3.6 | 19.6 | 6.3 | 17.3 | 9.8 | 13.3 | 2.4 | 5.2 | 2.3 |
| chT1D | 16.9 | 7.3 | 26.5 | 18.4 | 3.9 | 9.9 | 19.6 | 4.2 | 29.1 | 7.1 | 22.2 | 16.1 | 18.5 | 3.3 | 7.9 | 3.2 |
| cuT1D | 15.6 | 5.1 | 28.4 | 19.2 | 5.7 | 10.2 | 18.7 | 4.3 | 30.3 | 7.9 | 22.2 | 13.5 | 18.0 | 2.8 | 6.9 | 4.0 |
| LYMPH | ||||||||||||||||
| ngCo | 6.1 | 2.6 | 10.1 | 9.01 | 3.2 | 5.6 | 9.5 | 3.4 | 12.4 | 4.9 | 10.5 | 6.0 | 10.3 | 2.4 | 5.2 | 2.8 |
| acT1D | 6.2 | 2.5 | 8.2 | 7.1 | 2.5 | 4.4 | 7.2 | 2.8 | 10.8 | 3.9 | 9.9 | 5.5 | 8.8 | 2.0 | 3.8 | 1.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baskal, S.; Tsikas, S.A.; Begou, O.; Bollenbach, A.; Lenzen, S.; Jörns, A.; Tsikas, D. Advanced Glycation End-Products (AGEs) of Lysine and Effects of Anti-TCR/Anti-TNF-α Antibody-Based Therapy in the LEW.1AR1-iddm Rat, an Animal Model of Human Type 1 Diabetes. Int. J. Mol. Sci. 2022, 23, 1541. https://doi.org/10.3390/ijms23031541
Baskal S, Tsikas SA, Begou O, Bollenbach A, Lenzen S, Jörns A, Tsikas D. Advanced Glycation End-Products (AGEs) of Lysine and Effects of Anti-TCR/Anti-TNF-α Antibody-Based Therapy in the LEW.1AR1-iddm Rat, an Animal Model of Human Type 1 Diabetes. International Journal of Molecular Sciences. 2022; 23(3):1541. https://doi.org/10.3390/ijms23031541
Chicago/Turabian StyleBaskal, Svetlana, Stefanos A. Tsikas, Olga Begou, Alexander Bollenbach, Sigurd Lenzen, Anne Jörns, and Dimitrios Tsikas. 2022. "Advanced Glycation End-Products (AGEs) of Lysine and Effects of Anti-TCR/Anti-TNF-α Antibody-Based Therapy in the LEW.1AR1-iddm Rat, an Animal Model of Human Type 1 Diabetes" International Journal of Molecular Sciences 23, no. 3: 1541. https://doi.org/10.3390/ijms23031541