
Genome-Wide CRISPR/Cas9-Based Screening for Deubiquitinase Subfamily Identifies Ubiquitin-Specific Protease 11 as a Novel Regulator of Osteogenic Differentiation

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
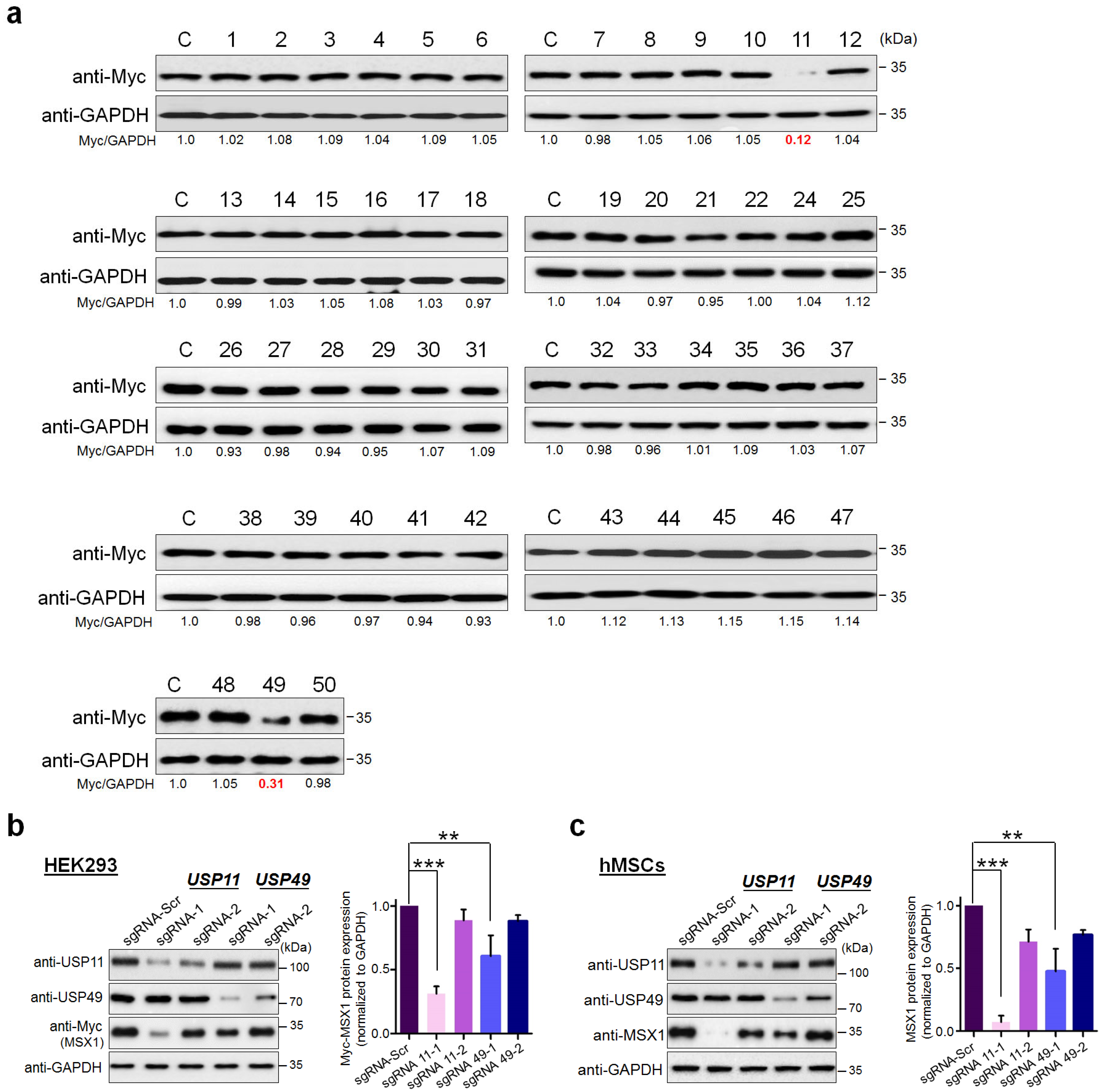
2.1. CRISPR-Based Genome-Wide Screening for the DUBs Regulating MSX1 Protein Levels
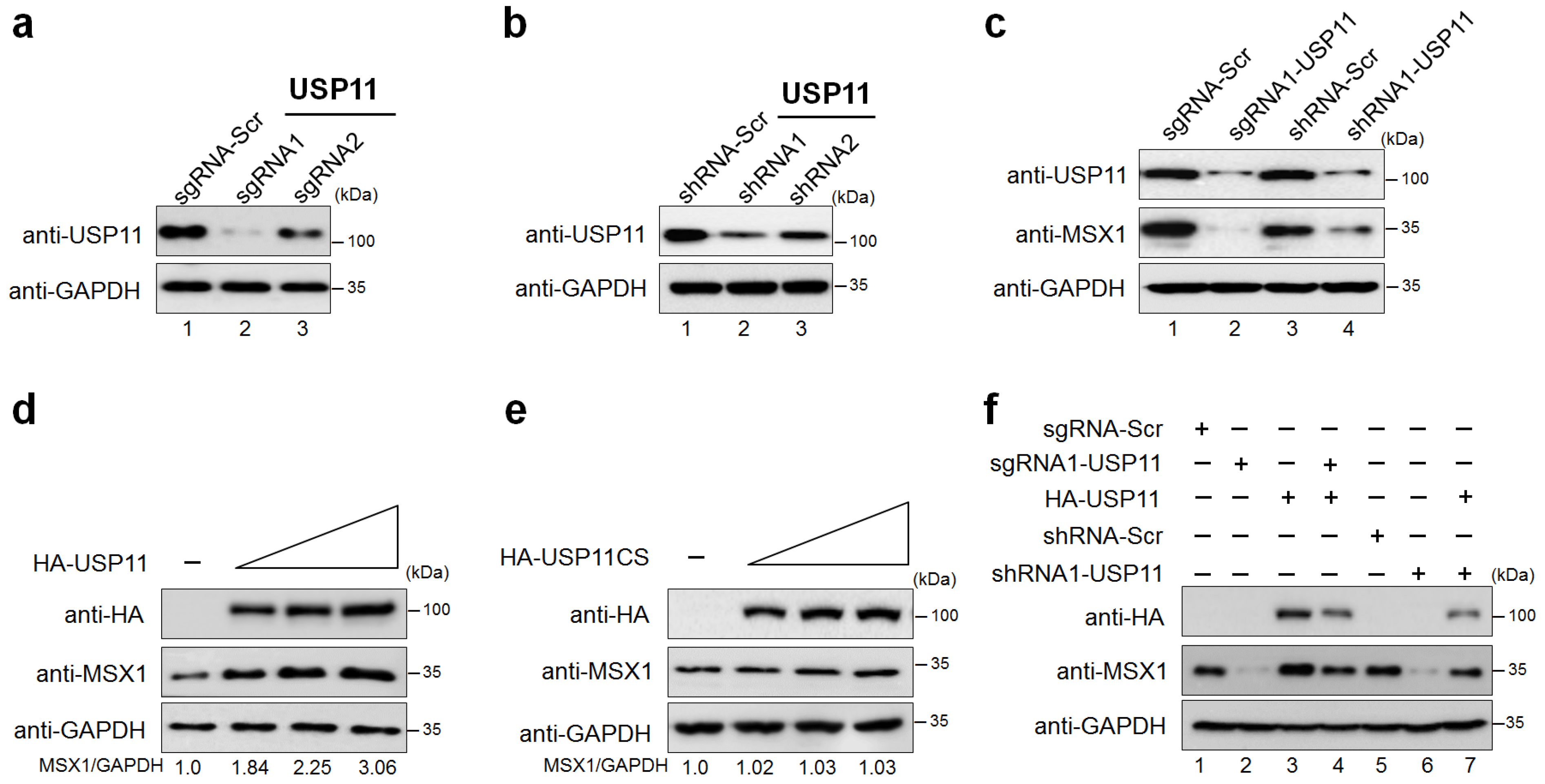
2.2. USP11 Increases MSX1 Protein Level
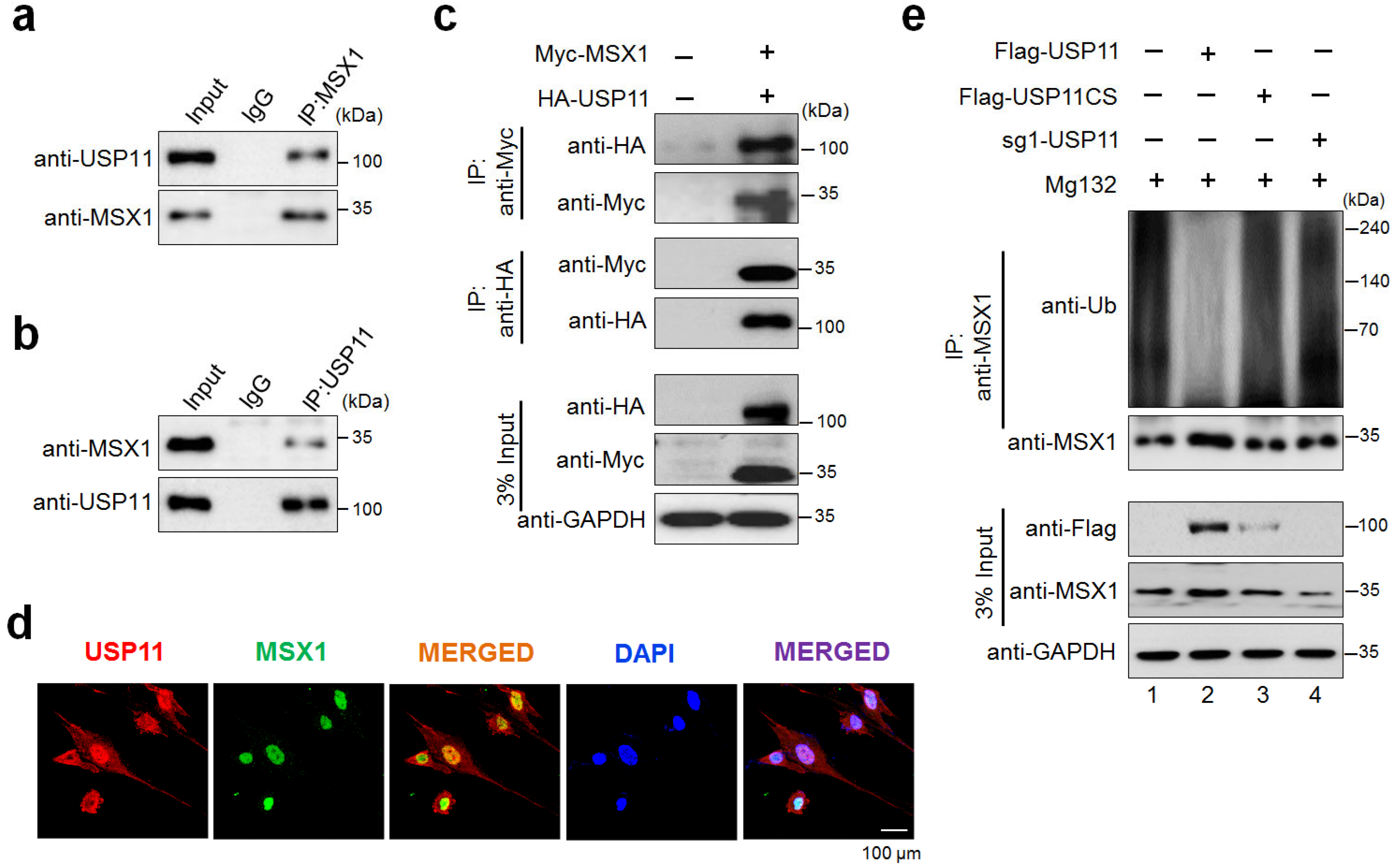
2.3. USP11 Interacts and Deubiquitinates MSX1
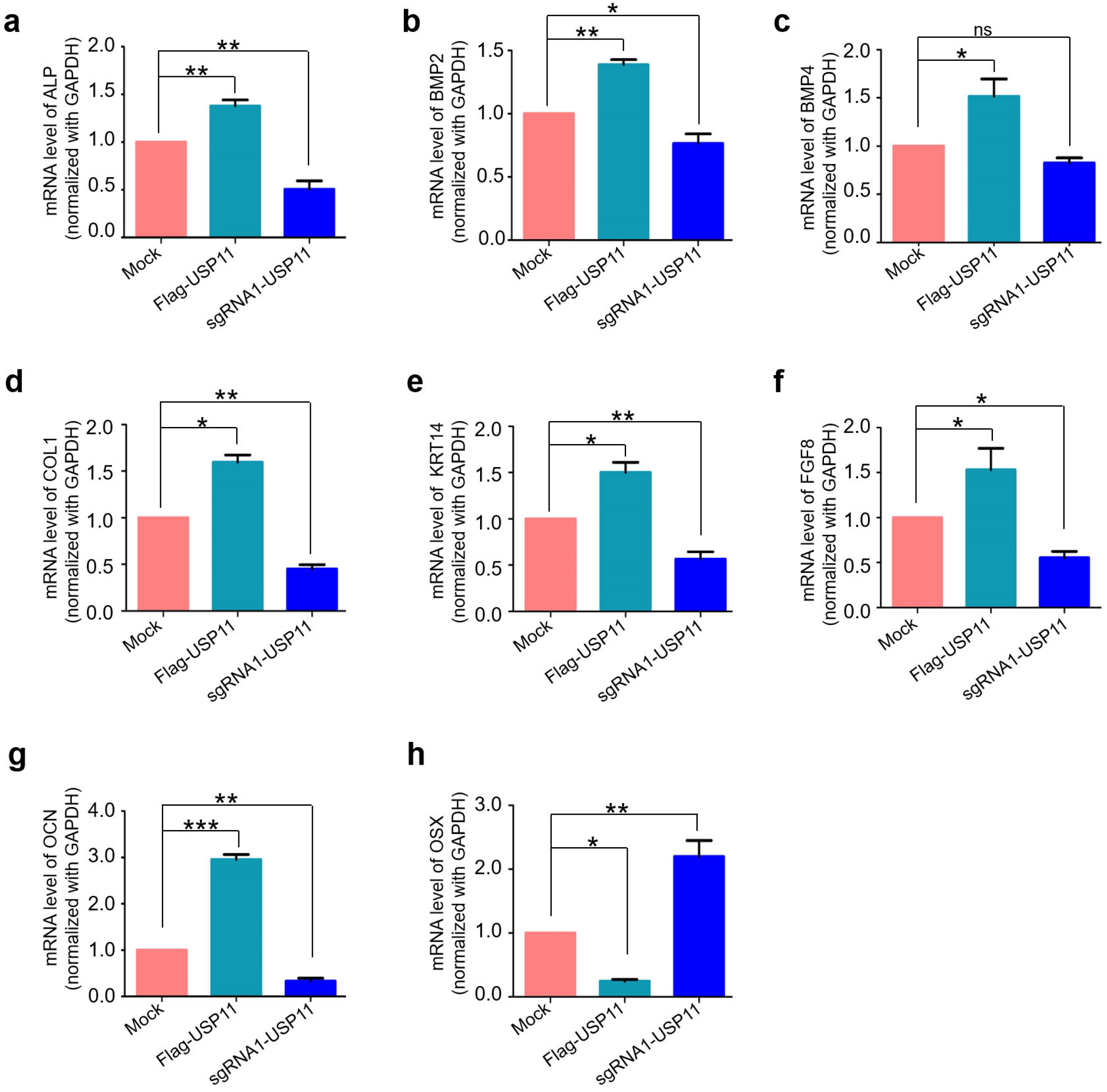
2.4. USP11 Enhances Osteogenic Transcriptional Factors in hMSCs
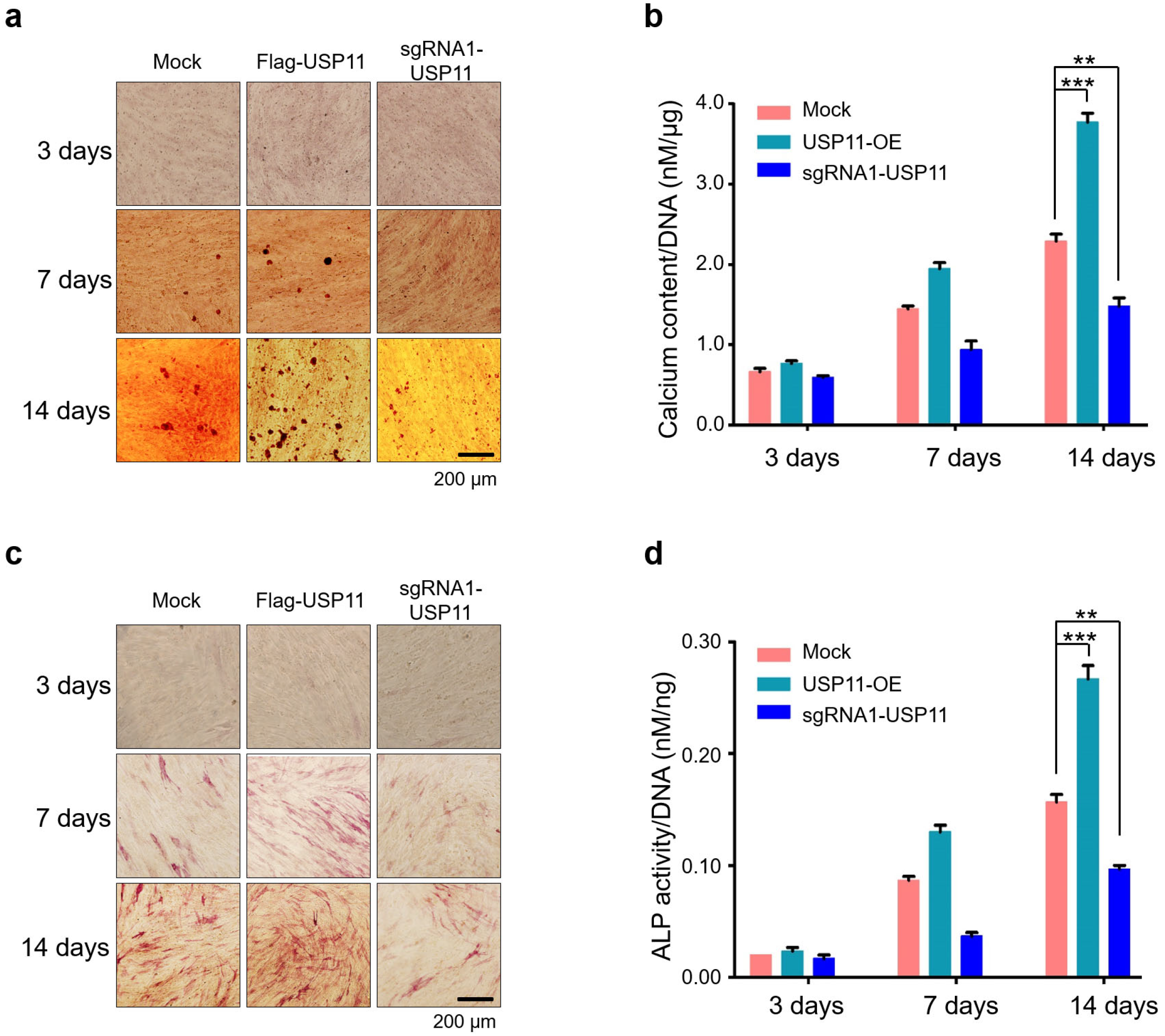
2.5. USP11 Enhances Osteogenic Activity in hMSCs
3. Discussion
4. Materials and Methods
4.1. Plasmids, sgRNA, and shRNA
4.2. Antibodies
4.3. Cell Culture
4.4. Transient Transfection and Transduction
4.5. T7E1 Assay
4.6. Immunofluorescence
4.7. Immunoprecipitation
4.8. Deubiquitination Assay
4.9. Real-Time PCR
4.10. Osteogenic Differentiation of hMSCs
4.11. Alizarin Red Staining
4.12. Alkaline Phosphatase Activity
4.13. DNA Content for Alizarin Red Staining and ALP Assays
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ALP | Alkaline phosphatase |
| BMPs | Bone morphogenetic proteins |
| BSA | Bovine serum albumin |
| CHX | Cycloheximide |
| COL1 | Collagen I |
| CRISPR | Clustered regularly interspaced short palindromic repeats |
| DSB | Double-strand break |
| DUBs | Deubiquitinating enzymes |
| FGF | Fibroblast growth factor |
| HEK293 | human embryonic kidney cells |
| hMSCs | human Mesenchymal stem cells |
| IP | Immunoprecipitation |
| KRT14 | Keratin 14 |
| MSX1 | Msh homeobox 1 |
| OCN | Osteocalcin |
| OSX | Osterix |
| PAGE | Polyacrylamide gel electrophoresis |
| PAM | Protospacer adjacent motif |
| PTM | post-translational modification, |
| RUNX2 | Runt-related transcription factor 2 |
| SDS | Sodium dodecyl sulfate |
| sgRNA | Single guide RNA |
| shRNA | Short hairpin RNA |
| TGFβ | Transforming growth factor β |
| T7E1 | T7 Endonuclease I |
| UCH | Ubiquitin carboxyl-terminal hydrolase |
| USP | Ubiquitin-specific protease |
References
- Holland, P.W.; Takahashi, T. The evolution of homeobox genes: Implications for the study of brain development. Brain Res. Bull. 2005, 66, 484–490. [Google Scholar] [CrossRef]
- Ramos, C.; Robert, B. msh/Msx gene family in neural development. Trends Genet. 2005, 21, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Lallemand, Y.; Nicola, M.A.; Ramos, C.; Bach, A.; Cloment, C.S.; Robert, B. Analysis of Msx1; Msx2 double mutants reveals multiple roles for Msx genes in limb development. Development 2005, 132, 3003–3014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orestes-Cardoso, S.M.; Nefussi, J.R.; Hotton, D.; Mesbah, M.; Orestes-Cardoso, M.D.; Robert, B.; Berdal, A. Postnatal Msx1 expression pattern in craniofacial, axial, and appendicular skeleton of transgenic mice from the first week until the second year. Dev. Dyn. 2001, 221, 1–13. [Google Scholar] [CrossRef]
- Nassif, A.; Senussi, I.; Meary, F.; Loiodice, S.; Hotton, D.; Robert, B.; Bensidhoum, M.; Berdal, A.; Babajko, S. Msx1 role in craniofacial bone morphogenesis. Bone 2014, 66, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Satokata, I.; Maas, R. Msx1 deficient mice exhibit cleft palate and abnormalities of craniofacial and tooth development. Nat. Genet 1994, 6, 348–356. [Google Scholar] [CrossRef]
- Yang, Y.; Zhu, X.; Jia, X.; Hou, W.; Zhou, G.; Ma, Z.; Yu, B.; Pi, Y.; Zhang, X.; Wang, J.; et al. Phosphorylation of Msx1 promotes cell proliferation through the Fgf9/18-MAPK signaling pathway during embryonic limb development. Nucleic Acids Res. 2020, 48, 11452–11467. [Google Scholar] [CrossRef]
- Vi-Fane, B.; Fernandes, I.; Davideau, J.L. Msx1 and its influence on craniofacial growth. Orthod Fr. 2007, 78, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Maas, R.; Chen, Y.P.; Bei, M.; Woo, I.; Satokata, I. The role of Msx genes in mammalian development. Ann. N. Y. Acad. Sci. 1996, 785, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Vastardis, H.; Karimbux, N.; Guthua, S.W.; Seidman, J.G.; Seidman, C.E. A human MSX1 homeodomain missense mutation causes selective tooth agenesis. Nat. Genet. 1996, 13, 417–421. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Machida, J.; Kamamoto, M.; Kimura, M.; Shibata, A.; Tatematsu, T.; Miyachi, H.; Higashi, Y.; Jezewski, P.; Nakayama, A.; et al. Characterization of novel MSX1 mutations identified in Japanese patients with nonsyndromic tooth agenesis. PLoS ONE 2014, 9, e102944. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Yu, M.; Liu, H.; Cai, T.; Feng, H.; Liu, Y.; Han, D. Novel MSX1 variants identified in families with nonsyndromic oligodontia. Int. J. Oral Sci. 2021, 13, 2. [Google Scholar] [CrossRef]
- Gou, Y.; Li, J.; Wu, J.; Gupta, R.; Cho, I.; Ho, T.V.; Chai, Y.; Merrill, A.; Wang, J.; Xu, J. Prmt1 regulates craniofacial bone formation upstream of Msx1. Mech. Dev. 2018, 152, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Gomathi, K.; Akshaya, N.; Srinaath, N.; Moorthi, A.; Selvamurugan, N. Regulation of Runx2 by post-translational modifications in osteoblast differentiation. Life Sci. 2020, 245, 117389. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Movérare-Skrtic, S.; Zhang, F.P.; Koskela, A.; Tuukkanen, J.; Palvimo, J.J.; Sipilä, P.; Poutanen, M.; Ohlsson, C. Androgen receptor SUMOylation regulates bone mass in male mice. Mol. Cell. Endocrinol. 2019, 479, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Sévère, N.; Dieudonné, F.X.; Marie, P.J. E3 ubiquitin ligase-mediated regulation of bone formation and tumorigenesis. Cell Death Dis. 2013, 4, e463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.C.; Zhang, S.W.; Yuan, Q. Deubiquitinating Enzymes and Bone Remodeling. Stem Cells Int. 2018, 2018, 3712083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.; Li, F.; Fang, P.; Dai, T.; Yang, B.; van Dam, H.; Jia, J.; Zheng, M.; Zhang, L. Ubiquitin-Specific Protease 4 Antagonizes Osteoblast Differentiation Through Dishevelled. J. Bone Miner. Res. 2016, 31, 1888–1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Lv, L.; Li, W.; Zhang, X.; Jiang, Y.; Ge, W.; Zhou, Y. Protein deubiquitinase USP7 is required for osteogenic differentiation of human adipose-derived stem cells. Stem Cell Res. Ther. 2017, 8, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, V.; Bei, M. Modification of Msx1 by SUMO-1. Biochem. Biophys. Res. Commun. 2006, 345, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.J.; Lee, H. PIAS1 negatively regulates ubiquitination of Msx1 homeoprotein independent of its SUMO ligase activity. Mol. Cells 2011, 32, 221–226. [Google Scholar] [CrossRef] [Green Version]
- Antao, A.M.; Tyagi, A.; Kim, K.S.; Ramakrishna, S. Advances in Deubiquitinating Enzyme Inhibition and Applications in Cancer Therapeutics. Cancers 2020, 12, 1579. [Google Scholar] [CrossRef]
- Kaushal, K.; Ramakrishna, S. Deubiquitinating Enzyme-Mediated Signaling Networks in Cancer Stem Cells. Cancers 2020, 12, 3253. [Google Scholar] [CrossRef]
- Shanmugham, A.; Ovaa, H. DUBs and disease: Activity assays for inhibitor development. Curr. Opin. Drug Discov. Dev. 2008, 11, 688–696. [Google Scholar]
- Singhal, S.; Taylor, M.C.; Baker, R.T. Deubiquitylating enzymes and disease. BMC Biochem. 2008, 9 (Suppl. 1), S3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petroski, M.D. The ubiquitin system, disease, and drug discovery. BMC Biochem. 2008, 9 (Suppl. 1), S7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurihara, L.J.; Kikuchi, T.; Wada, K.; Tilghman, S.M. Loss of Uch-L1 and Uch-L3 leads to neurodegeneration, posterior paralysis and dysphagia. Hum. Mol. Genet. 2001, 10, 1963–1970. [Google Scholar] [CrossRef] [Green Version]
- Jin, W.; Chang, M.; Paul, E.M.; Babu, G.; Lee, A.J.; Reiley, W.; Wright, A.; Zhang, M.; You, J.; Sun, S.C. Deubiquitinating enzyme CYLD negatively regulates RANK signaling and osteoclastogenesis in mice. J. Clin. Invest. 2008, 118, 1858–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, T.; Kapadia, H.; Feng, J.Q.; Raghow, R.; Peters, H.; D’Souza, R.N. Functional consequences of interactions between Pax9 and Msx1 genes in normal and abnormal tooth development. J. Biol. Chem. 2006, 281, 18363–18369. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Von den Hoff, J.; Lange, J.; Ren, Y.; Bian, Z.; Carels, C.E. MSX1 mutations and associated disease phenotypes: Genotype-phenotype relations. Eur. J. Hum. Genet. 2016, 24, 1663–1670. [Google Scholar] [CrossRef] [Green Version]
- van den Boogaard, M.J.; Dorland, M.; Beemer, F.A.; van Amstel, H.K. MSX1 mutation is associated with orofacial clefting and tooth agenesis in humans. Nat. Genet. 2000, 24, 342–343. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Chandrasekaran, A.P.; Suresh, B.; Haq, S.; Kang, J.H.; Lee, S.J.; Kim, J.; Kim, J.; Lee, S.; Kim, H.H.; et al. Genome-scale screening of deubiquitinase subfamily identifies USP3 as a stabilizer of Cdc25A regulating cell cycle in cancer. Cell Death Differ. 2020, 27, 3004–3020. [Google Scholar] [CrossRef] [PubMed]
- Goto, N.; Fujimoto, K.; Fujii, S.; Ida-Yonemochi, H.; Ohshima, H.; Kawamoto, T.; Noshiro, M.; Shukunami, C.; Kozai, K.; Kato, Y. Role of MSX1 in Osteogenic Differentiation of Human Dental Pulp Stem Cells. Stem Cells Int. 2016, 2016, 8035759. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Dezawa, M.; Ishikawa, H.; Itokazu, Y.; Yoshihara, T.; Hoshino, M.; Takeda, S.; Ide, C.; Nabeshima, Y. Bone marrow stromal cells generate muscle cells and repair muscle degeneration. Science 2005, 309, 314–317. [Google Scholar] [CrossRef] [PubMed]
- Charbord, P. Bone marrow mesenchymal stem cells: Historical overview and concepts. Hum. Gene. Ther. 2010, 21, 1045–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zernik, J.; Twarog, K.; Upholt, W.B. Regulation of alkaline phosphatase and alpha 2(I) procollagen synthesis during early intramembranous bone formation in the rat mandible. Differentiation 1990, 44, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Bendall, A.J.; Ding, J.; Hu, G.; Shen, M.M.; Abate-Shen, C. Msx1 antagonizes the myogenic activity of Pax3 in migrating limb muscle precursors. Development 1999, 126, 4965–4976. [Google Scholar] [CrossRef] [PubMed]
- Orestes-Cardoso, S.; Nefussi, J.R.; Lezot, F.; Oboeuf, M.; Pereira, M.; Mesbah, M.; Robert, B.; Berdal, A. Msx1 is a regulator of bone formation during development and postnatal growth: In vivo investigations in a transgenic mouse model. Connect. Tissue Res. 2002, 43, 153–160. [Google Scholar] [CrossRef]
- Davidson, D.R.; Crawley, A.; Hill, R.E.; Tickle, C. Position-dependent expression of two related homeobox genes in developing vertebrate limbs. Nature 1991, 352, 429–431. [Google Scholar] [CrossRef]
- Houzelstein, D.; Auda-Boucher, G.; Chéraud, Y.; Rouaud, T.; Blanc, I.; Tajbakhsh, S.; Buckingham, M.E.; Fontaine-Pérus, J.; Robert, B. The homeobox gene Msx1 is expressed in a subset of somites, and in muscle progenitor cells migrating into the forelimb. Development 1999, 126, 2689–2701. [Google Scholar] [CrossRef] [PubMed]
- Odelberg, S.J.; Kollhoff, A.; Keating, M.T. Dedifferentiation of mammalian myotubes induced by msx1. Cell 2000, 103, 1099–1109. [Google Scholar] [CrossRef] [Green Version]
- Suresh, B.; Lee, J.; Kim, K.S.; Ramakrishna, S. The Importance of Ubiquitination and Deubiquitination in Cellular Reprogramming. Stem Cells Int. 2016, 2016, 6705927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushal, K.; Antao, A.M.; Kim, K.S.; Ramakrishna, S. Deubiquitinating enzymes in cancer stem cells: Functions and targeted inhibition for cancer therapy. Drug Discov. Today 2018, 23, 1974–1982. [Google Scholar] [CrossRef]
- Rucci, N. Molecular biology of bone remodelling. Clin. Cases Miner. Bone Metab. 2008, 5, 49–56. [Google Scholar] [PubMed]
- Ensrud, K.E.; Crandall, C.J. Osteoporosis. Ann. Intern. Med. 2017, 167, itc17–itc32. [Google Scholar] [CrossRef] [PubMed]
- Singer, F.R. Bone Quality in Paget’s Disease of Bone. Curr. Osteoporos. Rep. 2016, 14, 39–42. [Google Scholar] [CrossRef]
- Zhou, J.; Fujiwara, T.; Ye, S.; Li, X.; Zhao, H. Ubiquitin E3 Ligase LNX2 is Critical for Osteoclastogenesis In Vitro by Regulating M-CSF/RANKL Signaling and Notch2. Calcif. Tissue Int. 2015, 96, 465–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strickson, S.; Emmerich, C.H.; Goh, E.T.H.; Zhang, J.; Kelsall, I.R.; Macartney, T.; Hastie, C.J.; Knebel, A.; Peggie, M.; Marchesi, F.; et al. Roles of the TRAF6 and Pellino E3 ligases in MyD88 and RANKL signaling. Proc. Natl. Acad. Sci. USA 2017, 114, E3481–E3489. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Qiao, M.; Harris, S.E.; Oyajobi, B.O.; Mundy, G.R.; Chen, D. Smurf1 inhibits osteoblast differentiation and bone formation in vitro and in vivo. J. Biol. Chem. 2004, 279, 12854–12859. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Huang, J.; Guo, R.; Wang, Y.; Chen, D.; Xing, L. Smurf1 inhibits mesenchymal stem cell proliferation and differentiation into osteoblasts through JunB degradation. J. Bone Miner. Res. 2010, 25, 1246–1256. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Yamashita, M.; Zhang, Q.; Zhou, Q.; Chen, D.; Reynolds, D.G.; Awad, H.A.; Yanoso, L.; Zhao, L.; Schwarz, E.M.; et al. Ubiquitin ligase Smurf1 mediates tumor necrosis factor-induced systemic bone loss by promoting proteasomal degradation of bone morphogenetic signaling proteins. J. Biol. Chem. 2008, 283, 23084–23092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, E.J.; Lee, K.Y.; Choi, N.S.; Lee, M.H.; Kim, H.N.; Jin, Y.H.; Ryoo, H.M.; Choi, J.Y.; Yoshida, M.; Nishino, N.; et al. Bone morphogenetic protein-2 stimulates Runx2 acetylation. J. Biol. Chem. 2006, 281, 16502–16511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Xie, Z.; Ma, Y.; Pan, X.; Wang, J.; Chen, Z.; Shi, P. TGF-β inhibits osteogenesis by upregulating the expression of ubiquitin ligase SMURF1 via MAPK-ERK signaling. J. Cell. Physiol. 2018, 233, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.; Zhang, H.; Boyce, B.F.; Xing, L. Ubiquitin E3 ligase Wwp1 negatively regulates osteoblast function by inhibiting osteoblast differentiation and migration. J. Bone Miner. Res. 2013, 28, 1925–1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Li, X.; Zhang, H.; Gu, R.; Wang, Z.; Gao, Z.; Xing, L. Ubiquitin E3 ligase Itch negatively regulates osteoblast function by promoting proteasome degradation of osteogenic proteins. Bone Joint Res. 2017, 6, 154–161. [Google Scholar] [CrossRef] [Green Version]
- Vlasschaert, C.; Xia, X.; Coulombe, J.; Gray, D.A. Evolution of the highly networked deubiquitinating enzymes USP4, USP15, and USP11. BMC Evol. Biol. 2015, 15, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Salihi, M.A.; Herhaus, L.; Macartney, T.; Sapkota, G.P. USP11 augments TGFβ signalling by deubiquitylating ALK5. Open Biol. 2012, 2, 120063. [Google Scholar] [CrossRef] [Green Version]
- Herhaus, L.; Sapkota, G.P. The emerging roles of deubiquitylating enzymes (DUBs) in the TGFβ and BMP pathways. Cell Signal. 2014, 26, 2186–2192. [Google Scholar] [CrossRef] [Green Version]
- Greenblatt, M.B.; Shin, D.Y.; Oh, H.; Lee, K.Y.; Zhai, B.; Gygi, S.P.; Lotinun, S.; Baron, R.; Liu, D.; Su, B.; et al. MEKK2 mediates an alternative β-catenin pathway that promotes bone formation. Proc. Natl. Acad. Sci. USA 2016, 113, E1226–E1235. [Google Scholar] [CrossRef] [Green Version]
- Capulli, M.; Paone, R.; Rucci, N. Osteoblast and osteocyte: Games without frontiers. Arch. Biochem. Biophys. 2014, 561, 3–12. [Google Scholar] [CrossRef]
- Danikowski, K.M.; Cheng, T. Colorimetric Analysis of Alkaline Phosphatase Activity in S. aureus Biofilm. J. Vis. Exp. 2019, 146, e59285. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, N.; Shigemasa, K.; Takagi, M. Gene expression and immunohistochemical localization of decorin and biglycan in association with early bone formation in the developing mandible. J. Oral Sci. 2001, 43, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Son, M.J.; Rho, S.B.; Kim, K.; Oh, M.; Son, C.; Song, S.Y.; Park, K. Homeoprotein Msx1-PIASy Interaction Inhibits Angiogenesis. Cells 2020, 9, 1854. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, S.; Kwaku Dad, A.B.; Beloor, J.; Gopalappa, R.; Lee, S.K.; Kim, H. Gene disruption by cell-penetrating peptide-mediated delivery of Cas9 protein and guide RNA. Genome Res. 2014, 24, 1020–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Lee, H.J.; Kim, H.; Cho, S.W.; Kim, J.S. Targeted genome editing in human cells with zinc finger nucleases constructed via modular assembly. Genome Res. 2009, 19, 1279–1288. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaushal, K.; Tyagi, A.; Karapurkar, J.K.; Kim, E.-J.; Tanguturi, P.; Kim, K.-S.; Jung, H.-S.; Ramakrishna, S. Genome-Wide CRISPR/Cas9-Based Screening for Deubiquitinase Subfamily Identifies Ubiquitin-Specific Protease 11 as a Novel Regulator of Osteogenic Differentiation. Int. J. Mol. Sci. 2022, 23, 856. https://doi.org/10.3390/ijms23020856
Kaushal K, Tyagi A, Karapurkar JK, Kim E-J, Tanguturi P, Kim K-S, Jung H-S, Ramakrishna S. Genome-Wide CRISPR/Cas9-Based Screening for Deubiquitinase Subfamily Identifies Ubiquitin-Specific Protease 11 as a Novel Regulator of Osteogenic Differentiation. International Journal of Molecular Sciences. 2022; 23(2):856. https://doi.org/10.3390/ijms23020856
Chicago/Turabian StyleKaushal, Kamini, Apoorvi Tyagi, Janardhan Keshav Karapurkar, Eun-Jung Kim, Parthasaradhireddy Tanguturi, Kye-Seong Kim, Han-Sung Jung, and Suresh Ramakrishna. 2022. "Genome-Wide CRISPR/Cas9-Based Screening for Deubiquitinase Subfamily Identifies Ubiquitin-Specific Protease 11 as a Novel Regulator of Osteogenic Differentiation" International Journal of Molecular Sciences 23, no. 2: 856. https://doi.org/10.3390/ijms23020856