
Phytosterols: Potential Metabolic Modulators in Neurodegenerative Diseases


1
Bionano Research Institute, Gachon University, 1342 Seongnam-daero, Sujeong-gu, Seongnam-si 461-701, Gyeonggi-do, Korea
2
Research Center for the Natural and Applied Sciences, College of Science, University of Santo Tomas, Manila 1015, Philippines
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to work.
Int. J. Mol. Sci. 2021, 22(22), 12255; https://doi.org/10.3390/ijms222212255
Submission received: 15 October 2021
/
Revised: 3 November 2021
/
Accepted: 9 November 2021
/
Published: 12 November 2021
(This article belongs to the Special Issue Health Properties of Plant Bioactive Compounds: Immune, Antioxidant and Metabolic Effects)
Abstract
:Phytosterols constitute a class of natural products that are an important component of diet and have vast applications in foods, cosmetics, and herbal medicines. With many and diverse isolated structures in nature, they exhibit a broad range of biological and pharmacological activities. Among over 200 types of phytosterols, stigmasterol and β-sitosterol were ubiquitous in many plant species, exhibiting important aspects of activities related to neurodegenerative diseases. Hence, this mini-review presented an overview of the reported studies on selected phytosterols related to neurodegenerative diseases. It covered the major phytosterols based on biosynthetic considerations, including other phytosterols with significant in vitro and in vivo biological activities.
1. Introduction
Neurodegenerative disease (ND) is a group of disorders (Alzheimer’s, Parkinson’s, multiple sclerosis, amyotrophic lateral sclerosis, Huntington’s disease, prion disease, etc.) that result in progressive degeneration of structure or function of the neurons. Neurodegeneration may progress differently with various levels and locations of neuronal circuitry in the brain, ranging from molecular to systemic pathways.
Alzheimer’s disease (AD) is the most common form of dementia, characterized by progressive memory loss, cognitive impairment, and behavioral complications. Pathophysiologically, AD is characterized by excessive amyloid-beta (Aβ) peptide aggregation, intracellular neurofibrillary tangles, highly phosphorylated tau protein, deficiency of essential neurotransmitters, and oxidative stress-induced neuronal damage [1,2,3]. To date, the FDA approved drugs have included medicines that may help minimize or alleviate symptoms by regulating chemicals implicated in neurotransmission, such as acetyl cholinesterase inhibitors (donepezil, galantamine, and rivastigmine) and glutamate receptor antagonists (memantine). Aducanumab (marketed as Aduhelm, an anti-amyloid antibody intravenous (IV) infusion therapy), a new drug recently approved by the FDA, may delay clinical degeneration with benefits to both cognitive and motor function in patients with AD. However, all of these treatments suffer from side effects ranging from headache, nausea, confusion, dizziness, falls, and amyloid-related imaging abnormalities (ARIA; including swelling in the brain and micro hemorrhaging/superficial siderosis). This justified the screening of new and safe therapeutic agents from natural sources. In this context, several plants and their bioactive components were reported to inhibit Aβ formation, cholinesterases, tau proteins aggregation, and free radicals [4,5,6,7]. Various phytosterols are anti-inflammatory in action and are known to abrogate production and activation of nitric oxide (NO), tumor necrosis factor-α (TNF-α), cyclooxygenase-2 (COX-2), inducible nitric oxide synthase (iNOS), and phosphorylated extracellular signal-regulated protein kinase (p-ERK) [8]. Hence, phytosterols may possess therapeutic implications in neurodegenerative diseases.
Phytosterol is a collective term for plant sterols and their saturated forms (stanols). They are present in dietary sources such as unrefined plant oils, nuts, seeds, and legumes [9]. Phytosterols can be called ‘plant cholesterol’ due to their structural similarity to cholesterol. An additional methyl group (campesterol) or ethyl group (sitosterol) at the C-24 position or an additional double bond at the C-22 position (brassicasterol or stigmasterol) differentiate them from cholesterol. Depending on the nature of the diet, sitosterol (65%), campesterol (23%), and stigmasterol (10%) belong to the major group among over 200 types of phytosterols [10]. Other important phytosterols such as brassicasterol, avenasterol, and fucosterol, along with the saturated forms of phytosterols (sitostanol, campestanol, and stigmastanol), are present in minor quantities in the diet. The dietary intake of phytosterols varies from 150–450 mg/day [11], but their intestinal absorption is less than 2% (phytosterols) and 0.2% (phytostanols) in comparison to cholesterol, which is nearly 50% [12,13]. As a result of low absorption and rapid biliary elimination, physiological concentrations of phytosterols in the plasma are on the order of 10−3 in comparison to cholesterol [14].
Phytosterols have recently gained the attention of researchers due to cholesterol and lipid lowering, anti-atherogenic and immunomodulating properties, especially with a report on the changes in cholesterol metabolism of patients with AD [15,16,17]. Essentially, there is rarely any information in association with their toxicity apart from the indistinct anabolic effect for some [18], and they have already been declared safe (up to 3 g/day) by the Food and Drug Administration (FDA) and the European Union Scientific Committee (EUSC) [19].
Phytosterols have the ability to cross the blood–brain barrier (BBB) and accumulate in the brain [20]. Hence, they might have a significant role in modulating various pathways in the brain linked to neurodegeneration. As the research or review reports on this important aspect were limited, herein, the influences of phytosterols in neurodegenerative diseases were reviewed and discussed. The most prevalent plant sterols, stigmasterol, β-sitosterol, and campesterol [21], were mainly discussed with several minor sterols— brassicasterol, lanosterol, 24(S)-saringosterol, 4,4-dimethyl phytosterols, and ergosterol—for their action mechanisms in potentially treating AD.
2. Biosynthesis of the Phytosterols in Plants
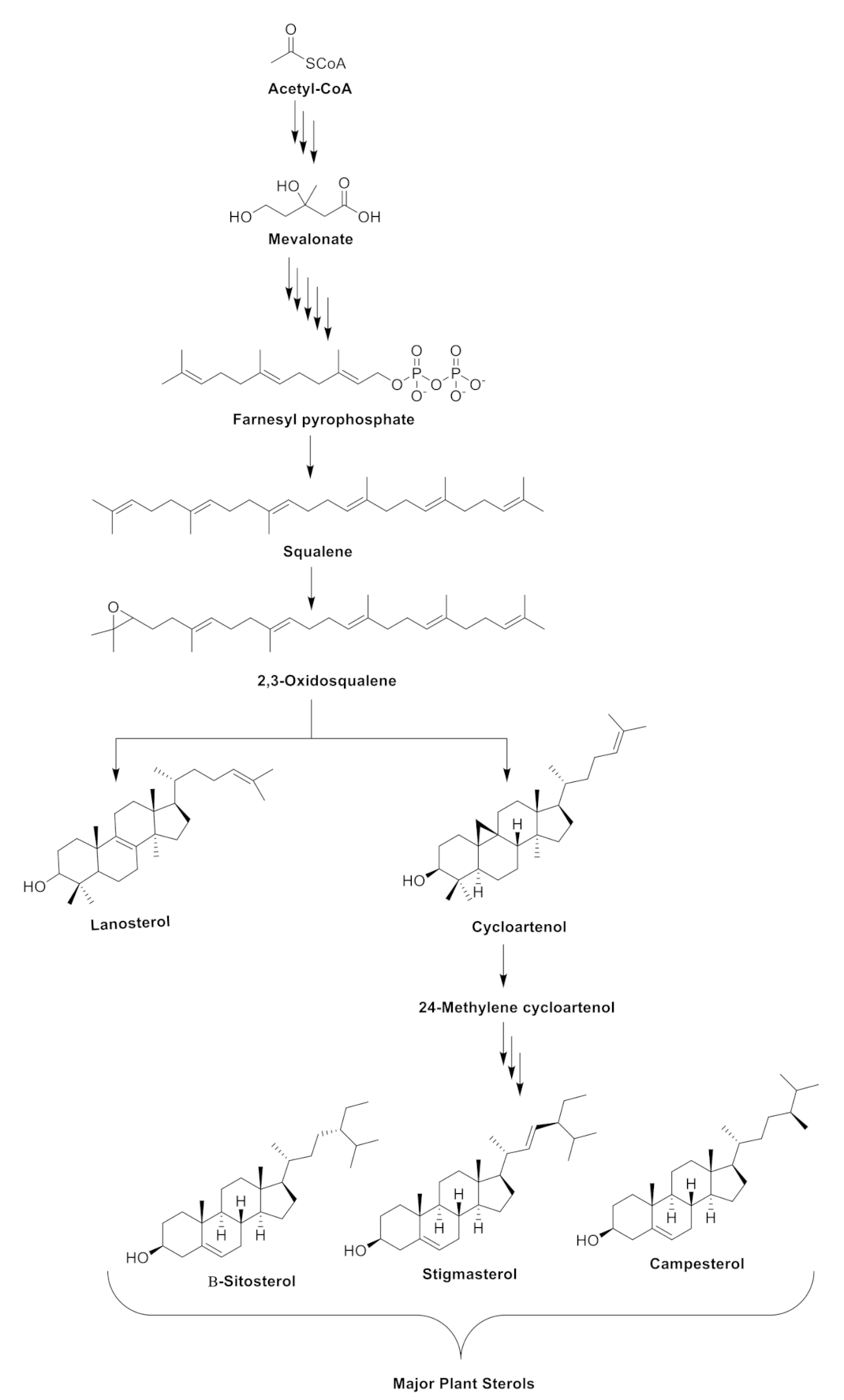
The biosynthesis of phytosterols in plants consists of three major steps. As highlighted in Zhang et al. [22], step one involves the mevalonate or isoprenoid pathway. With acetyl-CoA as the starting point, the process lead to the formation of mevalonate, which serves as an intermediate in the biosynthesis of isoprenoids. Step two is described as the cyclization of squalene and its subsequent conversion to 2,3-oxidosqualene, generating various triterpenes and the phytosterols (lanosterol and cycloartenol). In step three, cycloartenol is converted to 24-methylene cycloartenol by the action of C-24-sterol methyltransferase 1 (SMT1). As shown in Figure 1, the formation of the 24-methylene cycloartenol would facilitate the biosynthesis of the major plant sterols β-sitosterol, stigmasterol, and campesterol.
3. Plant Sterols in the Brain
As the exact mechanism through which phytosterols cross the BBB is still not fully understood, it was speculated that high density lipoprotein (HDL)-like particles carrying plant sterols and apolipoprotein E (ApoE) assist this process in brain [18,23,24]. The HDL-like particles are transported through the scavenger receptor class B type 1 (SR-B1) HDL receptors, present on the apical side of the BBB and released into the brain through ATP-binding cassette (ABCA/ABCG1) transporters, on the basolateral side of the BBB and on the astrocytes membrane. Low density and very low density lipoprotein (LDL/VLDL) receptors could transfer these HDL-like particles loaded with plant sterols on the surface of microglia and oligodendrocytes (Figure 2). Even though the brain accounts for 2.1% of the total body weight, it contains nearly 23% of all free cholesterol [25], which is synthesized by the brain itself (in situ), because the cholesterol in the circulation will not cross the BBB. A steady cholesterol turnover plays a critical role in synapse formation, cell–cell interactions, and intracellular signaling [25,26]. The level of cholesterol is maintained in the brain by removal of excess cholesterol out of the BBB in the form of polar 24-(S)-OH-cholesterol [27]. On the other hand, the metabolism of phytosterols is entirely different in the brain. Due to the presence of alkyl groups at C-24, they cannot be converted to more polarized derivatives. Therefore, after entering the brain circulation, phytosterols are irretrievably accumulated and incorporated into the cell membranes [28], where they display a positive role in reducing inflammation, Aβ levels, and β-secretase activity [29]. Again, the mechanism is still not confirmed, but is expected to occur through peroxisome proliferator-activated receptors (PPAR) signaling, involving the activation of liver X receptor/retinoid X receptor (LXR/RXR) and the regulation of apolipoprotein E (ApoE) [18]. The sterols from Aloe vera were reported to act through PPAR receptors and increased the expression of fatty acid transporter (FATP1), acyl-CoA- oxidase 1 (ACOX1), and carnitine palmitoyl transferase 1 (CPT1) in a dose-dependent manner, as well as by increasing glutathione and decreasing IL-18 expression [30]. Cholesterol is highly amyloidogenic; however, stigmasterol (having an additional double-bond at C-22/C-23 and an ethyl-group at C-24 in cholesterol skeleton) significantly decreased Aβ levels [31]. Evidence linked raised cholesterol levels to increased Aβ generation in cellular and most animal models of AD [32], and studies have indicated that drugs that inhibit cholesterol synthesis also lower Aβ in these models [33]; however, some other results are contradictory [34]. Studies showed that changes in cholesterol homeostasis, distribution within neurons, and compartmentation affected the processing of APP and Aβ generation [35]. The identification of a variant of the apolipoprotein E (APOE) gene as a major genetic risk factor for AD is also consistent with a role for cholesterol in the pathogenesis of AD [36]. The activity of an enzyme responsible for clearing cholesterol from peripheral cells, lecithin cholesterol acyltransferase (LCAT), is significantly decreased in atherosclerosis and AD [37].
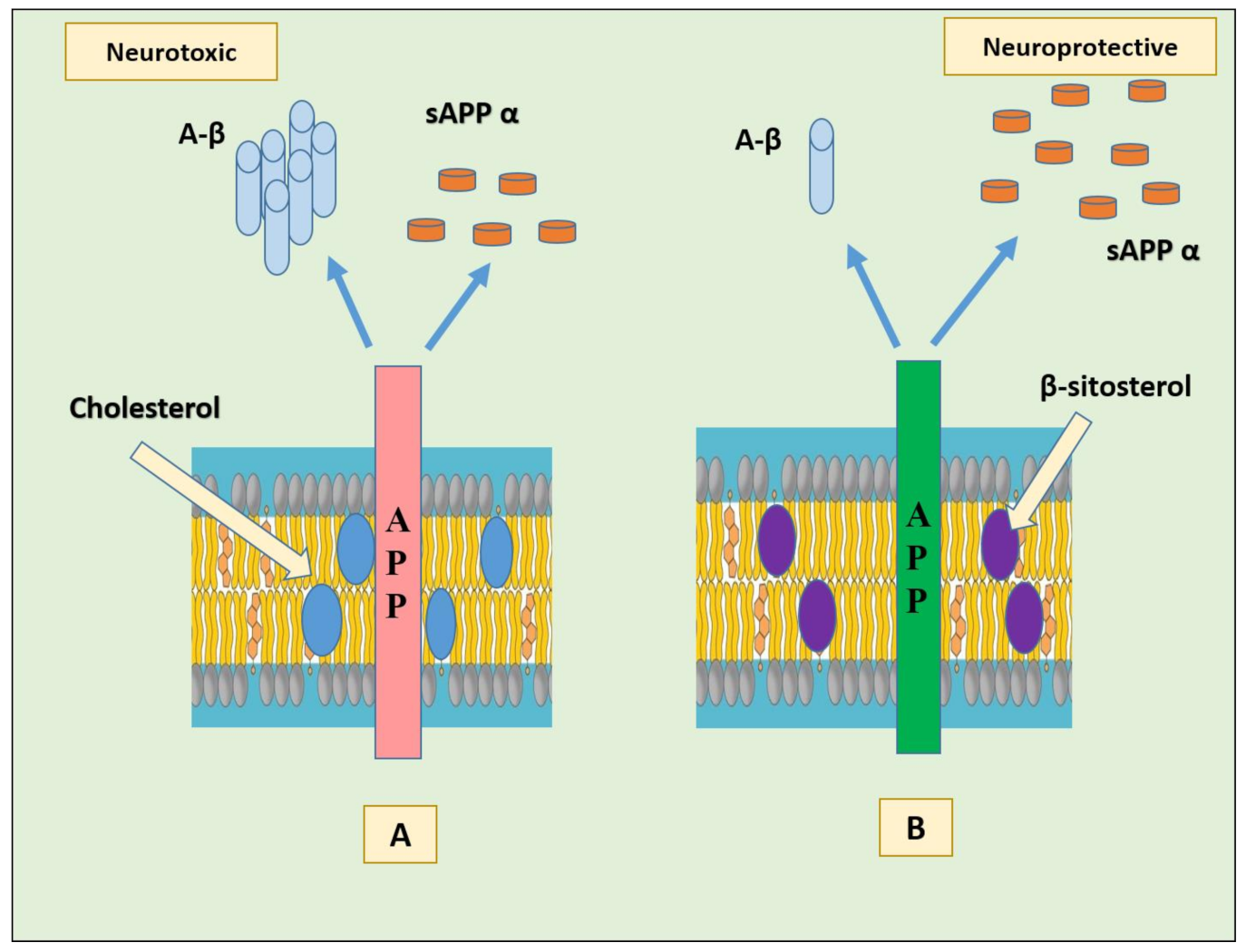
In particular, binding of cholesterol to the C-terminal transmembrane domain (C99, also known as the β-CTF) of the amyloid precursor protein (APP) seems to favor the amyloidogenic pathway in cells by endorsing localization of C99 in lipid rafts where γ-secretase and possibly β-secretase are present [38]. The products formed (amyloid-β and the intracellular domain of the APP (AICD)) might possibly down-regulate ApoE-mediated cholesterol uptake and cholesterol biosynthesis. Another possible mechanism by which cholesterol/C99 promote amyloidogenesis is via directly modifying substrate binding, catalysis, or product dissociation by β- or γ-secretase. The activity of these enzymes is much higher in the presence of cholesterol than in its absence [39]. The APP appears as a cellular cholesterol sensor; when membrane cholesterol levels are high, APP binds cholesterol to form a complex and promote the amyloidogenic pathway [40]. In a study using HT22 mouse hippocampal cells, the effect of substituting cholesterol with β- sitosterol was evaluated on APP metabolism. The results showed that the substitution stimulated non-amyloidogenic APP processing possibly by redistribution of APP from lipid rafts toward non-raft regions, without affecting membrane fluidity (Figure 3) [41].
In fact, they significantly reduced brain Aβ levels and β- and γ-secretase activities in vivo, suggesting that a diet enriched in plant sterols might be beneficial for AD. Additionally, the administration of a blend of sitosterol (60%), campesterol (25%), and stigmasterol (15%) has been shown to have an anti-inflammatory effect on central nervous system (CNS) demyelination in an autoimmune encephalomyelitis (EAE) model of multiple sclerosis (MS) [42]. The similar outcomes were expected for AD, as both AD and MS were tightly related in lipid metabolism. Phytosterols are also known to prevent disease progression and to improve motor defects by increasing myelin content by facilitating incorporation of proteolipid protein (PLP) into myelin membranes, improving density of oligodendrocytes and reducing inflammation in animal models [43,44]. RT-PCR results indicated the role of phytosterols in modulating the action of a variety of growth factors from their involvement in the differentiation and survival of oligodendrocyte precursor cells [43]. Phytosterol supplementation elevated the expression levels of fibroblast growth factor 1 (FGF1) and Sonic hedgehog (SHH), without affecting insulin-like growth factor 1 (IGF1), epidermal growth factor (EGF), and ciliary neurotrophic factor (CNTF).
4. Neuroprotective Effects of the Selected Phytosterols
This section highlights various biological activities in vitro or in vivo by the major phytosterols, such as β-sitosterol, stigmasterol, and campesterol, and other minor sterols brassicasterol, lanosterol, 24(S)-saringosterol, and the 4,4-dimethyl sterols. The structures of the selected phytosterols included in this review are shown in Figure 4, and a summary of their implicated neuroprotective activities is shown in Table 1 and Figure 5.
4.1. β-Sitosterol
β–Sitosterol is one of the main dietary phytosterols belonging to the class of organic compounds known as stigmastanes. These are sterol lipids with a structure based on the stigmastane skeleton, which consists of a cholestane moiety bearing an ethyl group at the C-24 position. In a comprehensive study, the effects of β-sitosterol in behavioral studies were observed in transgenic mice using a shallow water maze (SWM), Y-maze, and balance beam tests. The in vivo (in frontal cortex and hippocampus) and in vitro anti-acetyl choline esterase (AChE), anti-butyl choline esterase (BChE) inhibitory potentials, antioxidant activity, and molecular docking were also analyzed [45,46]. β-sitosterol revealed strong Ache properties under both in vitro and in vivo conditions. An IC50 value of 55 and 50 μg/mL against AChE and BChE, respectively [45], and 62 μg/mL against both AChE and BChE [46] were reported in vitro, while the activity was significantly lower in brain tissue homogenates. The AChE inhibition was further confirmed by in silico studies in which β-sitosterol was strongly bound to the active sites of AChE and BChE through the para-hydroxyl group of the phenolic moiety networked with the active site water molecule and the side chain carbonyl residues through H-bonding. The rest of the active compound was packed in a shallow pocket through H-bonding [46]. In the antioxidant assays, the IC50 values were observed as 140, 120, and 280 μg/mL in the 2,2-diphenyl-1-picryl-hydrazyl-hydrate (DPPH), 2,2 -azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) (ABTS), and hydrogen peroxide (H2O2) assays, respectively. The treatment group had lower oxidative stress in comparison to the disease control. β-Sitosterol was reported to increase levels of antioxidant enzymes by activating the estrogen receptor/PI3-kinase pathway. It also regulated glutathione levels, suggesting its role as an effective free radical scavenger [47,48]. Furthermore, glucose oxidase-mediated oxidative stress and lipid peroxidation might be inhibited from the incorporation of β-sitosterol into cell membrane, which showed valuable effects of this compound in neurodegenerative disorders including AD [47]. β-sitosterol also restored behavioral deficits, working memory, and motor coordination in transgenic animals [45]. It also hastened neuron degeneration in mice deficient for LXRβ [49], suggesting that LXRβ activation by non-glucosylated sitosterol is neuroprotective.
In another study, the effect and mechanism of β-sitosterol on deficits in learning and memory in amyloid protein precursor/presenilin 1 (APP/PS1) double transgenic mice was investigated. APP/PS1 mice were treated with β-sitosterol for 4 weeks, from the age of 7 months. Brain Aβ metabolism was evaluated using ELISA and Western blotting. β-sitosterol treatment ameliorated spatial learning and recognition memory capacity, along with reduction in plaque deposition and renovation of the excitatory postsynaptic current frequency in the hippocampus [50]. Substitution of cholesterol by β-sitosterol promoted non-amyloidogenic AP processing by migrating it to non-lipid raft region [41]. However, contradictory results were obtained in another study [31], where β-sitosterol increased the secretion of amyloid by 115.2% ± 2.2%.
Reports have suggested the role of cholesterol in amyloid precursor protein (APP) processing [40,51,52,53]; thus, cholesterol may be considered as a target for developing drugs to treat AD. Phytosterols are structurally similar to cholesterol and have been widely used to reduce blood cholesterol [54]. It was found that β-sitosterol efficiently repressed the release of high cholesterol-driven platelet Aβ. In addition, β-sitosterol also prevented high cholesterol-induced increases in β- and γ-secretase activity [55].
Mitochondrial dysfunction has a critical role in neuronal degeneration [56]. Therefore, augmentation of mitochondrial function by increasing the mitochondrial membrane potential (ΔΨm) and mitochondrial adenosine triphosphate (ATP) may be valuable in treating AD. Previously, it was suggested that an enhancement in mitochondrial ATP levels may be beneficial for neurodegenerative diseases [57]. Integration of β-sitosterol into mitochondrial membrane augmented the mitochondrial function by endorsing inner mitochondrial membrane fluidity, thereby raising the ∆Ψm and ATP concentrations. Hence, β-sitosterol could be an effective dietary therapy for neurodegenerative diseases such as AD [58].
Continual neuroinflammation in neurodegenerative diseases damages the neurons due to the release of toxic factors such as nitric oxide. In a study, β-sitosterol displayed anti-inflammatory action in BV2 cells upon exposure to LPS by reducing the expression of pro-inflammatory markers, such as interleukin-6 (IL-6), inducible nitric oxide (iNOS), tumor necrosis factor-α (TNF-α), and cyclooxygenase-2 (COX-2). It also suppressed the phosphorylation and degradation of inhibitor of nuclear factor kappa B (IκB) and inhibited the phosphorylation of nuclear factor kappa B (NF-κB) and extracellular signal-regulated kinase (ERK), which regulated various cytokines in inflammatory pathway [59].
All of these studies confirmed the multi-target potential of β-sitosterol in management of memory deficit disorders such as AD.
4.2. Stigmasterol
Stigmasterol is a steroid derivative having a hydroxyl group in position C-3 of the steroid skeleton, unsaturated bonds in position C-5/C-6 of the B ring, and alkyl substituents in position C-22/C-23. Stigmasterol is shown to be involved in neuroprotection through a multi-target approach. Stigmasterol isolated from Rhazya stricta fruits displayed in vitro AChE inhibitory activity with an IC50 of 644 ± 11.75 µM [60]. It was also reported to reduce amyloid plaques by decreasing the β-secretase cleavage of APP [31]. Stigmasterol displayed neuroprotective activity against glutamate-induced toxicity by inhibition of reactive oxygen species (ROS) and Ca2+ production in in vitro studies [61]. To investigate the possible neuroprotective mechanisms of stigmasterol, an H2O2-induced oxidative stress model in SH-SY5Y neuroblastoma cells was established [62]. H2O2 exposure raised the levels of ROS significantly within the cells, inducing apoptosis. However, pre-incubation with stigmasterol prevented oxidative stress-induced cell death by reducing the level of ROS in the cells. It also upregulated catalase, forkhead box O (FoxO) 3a, and anti-apoptotic protein B-cell lymphoma 2 (Bcl-2) in the neurons. Additionally, it also increased the expression levels of sirtuin 1 (SIRT1) and decreased the levels of acetylated lysine [62]. Sirtuins are highly conserved NAD (+)-dependent enzymes that have positive effects in age-related neurodegenerative diseases. In vitro and in vivo studies revealed that the increased SIRT1 protein ameliorated AD-like symptoms by reducing the memory decline [63]. Similarly, reduction of Aβ aggregations was reported through activation of SIRT1 causing upregulation of APP metabolism by α-secretase [64]. Moreover, overexpression of SIRT1 protected SH-SY5Y cells from toxicity-induced cell death [65]. Using docking studies, the possible interaction between stigmasterol and the activator binding site of SIRT1 was also inspected. According to the binding analysis, SIRT1 interacted with stigmasterol and resveratrol; both compounds bind to Thr209 and Pro212 by van der Waals and hydrophobic interactions, respectively. Moreover, they also shared binding interactions with Phe414, Asp292, Gln294, and Ala295 [62]. Stigmasterol was also reported to inhibit several pro-inflammatory cytokines in IL-1β-treated cells without affecting IL-6 levels, suggesting its role in the IL-1β-induced NF-κB inflammatory pathway [66].
Transcriptomic analysis implicated the role of stigmasterol in upregulating genes involved in neuritogenesis (Map2, Dcx, Reln) and synaptogenesis (Arc, Egr1, Nr4a1), thereby stimulating neuronal architecture in primary hippocampal neurons for processing memory and learning. Stigmasterol also decreased the expression of K+ transport genes to sustain neuronal excitability under adverse conditions [67]. Thus, the broad spectrum of action has made stigmasterol a potential therapeutic candidate for the prevention and treatment of brain disorders, especially AD. Mice fed with stigmasterol-enriched diets exhibited the reduction of Aβ generation by diminishing β-secretase activity, decreasing expression of all γ-secretase components, reducing cholesterol and presenilin distribution in lipid rafts involved in amyloidogenic APP cleavage, and by decreasing β-secretase 1 (BACE1) internalization to endosomal compartments [31].
4.3. Campesterol
Campesterol is characterized by the presence of a hydroxyl group at C-3 of the steroid skeleton and saturated bonds throughout the sterol structure, with the exception of the C-5/C-6 double bond in the B ring. Campesterol only marginally affected Aβ secretion in APP695-expressing SH-SY5Y cells, mainly due to increased γ-secretase activity. The β-secretase activity was also significantly increased in purified membranes of mouse brains (104.8% ± 1.4), whereas no effect was observed in SH-SY5Y wild type (wt) membranes. Both gene expression and protein levels of BACE1 were unchanged. However, campesterol significantly increased γ-secretase activity in the purified membranes of SH-SY5Y wt cells and mouse brains (115.5% ± 3.0 and 106.9%, respectively). Gene expression of all components of the γ-secretase complex remained unchanged in the presence of campesterol, reflected by unchanged PS1 protein level [31].
In a clinical trial of AD patients, sitosterol and campesterol levels in the brain were almost similar in comparison to the healthy control group (6.3 ± 0.8 ng/mg and 6.2 ± 0.8 ng/mg versus 5.0 ± 0.8 ng/mg and 5.0 ± 0.8 ng/mg, p < 0.05, in the assayed and control groups, respectively). However, the levels of 27-hydroxycholesterol were decreased due to oxidative damage, where the BBB was not disturbed [68].
4.4. Brassicasterol
Brassicasterol is a 3β-sterol, that is, (22E)-ergosta-5,22-diene substituted by a hydroxy group at position 3β. It is found in marine algae, fish, and rapeseed oil. In the early stages of AD, the functions of the BBB and choroid plexus are impaired, resulting in reduced concentrations of phytosterols in cerebrospinal fluid (CSF). Both sitosterol and brassicasterol were significantly reduced, but when corrected for CSF cholesterol concentrations, only brassicasterol was reduced significantly in CSF of AD patients. Brassicasterol also amended the predictive value, when added to the biomarkers pTau and Aβ-42. Therefore, brassicasterol may be a novel additional CSF biomarker of AD patients [69]. Although their study was not designed to determine whether brassicasterol levels in CSF may be prognostic of AD progression, their observations nonetheless generated interesting hypotheses about the utility of plant sterol metabolism as a potential biomarker in AD.
However, in another study [31], brassicasterol directly increased β-secretase activity in purified membranes of SH-SY5Y wt cells and mouse brains (108.5% ± 1.9 and 106.6% ± 1.7, respectively) without affecting gene expression and protein level of BACE1. Similarly, γ-secretase activity was also slightly, but significantly, increased in purified membranes of SH-SY5Y wt cells and mouse brains (108.2% ± 1.0 and 111.4% ± 1.6%, respectively). Considerable changes in gene expression were only found for the γ-secretase components presenilin 2(PS2)/anterior-pharynx-defective protein 1 (APH1B), but not for the homologues PS1 and APH1A, presenilin enhancer 2 (PEN2) [31].
4.5. Lanosterol
Lanosterol is a Δ8,24 phytosterol formed from the cyclization of 2,3-oxidosqualene catalyzed by lanosterol synthetase [22]. It was shown to be a potential candidate in the treatment of cataracts by disrupting the aggregations of intracrystalline proteins [70]. Several studies suggested it as a promising modulator of neurodegenerative diseases [71,72,73,74]. In silico studies revealed that lanosterol disrupted the aggregations of the amyloidogenic KLVFFA peptide chain in Aβ42 by hydrophobic interaction with Phe-19 and Phe-20 and interfering with the β-sheet-β-sheet packing interactions. This was further correlated with the thioflavin T (ThT) fluorescence assay and atomic force microscope (AFM) imaging. Furthermore, lanosterol also revealed neuroprotective effects with PC12 cells upon Aβ42 treatment [71].
The aggregation of misfolded proteins in neuronal cells is interconnected with neurodegeneration. In vitro report utilizing HeLa and HEK-293A cells revealed that lanosterol and lanosterol synthase facilitated the renewal of the cell’s trapped misfolded proteins in intracellular proteostasis. Lanosterol effectively minimized the number and size of sequestosomes/aggresomes in HeLa and HEK-293A cells. These results suggested a cytoprotective property of lanosterol in facilitating cell proliferation and survival by disaggregating and refolding ubiquitinated misfolded proteins [72]. Lanosterol treatment reduced the accumulations and cytotoxicity of misfolded protein aggregations through induction of co-chaperone (CHIP) and by promoting autophagy [73].
The neuroprotective effects of lanosterol was also demonstrated in vivo [74]. Administration of lanosterol to 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-treated mice showed lanosterol reduction in the nigrostriatal region, suggesting an altered lanosterol metabolism involvement in Parkinson’s disease pathogenesis. Treatment with lanosterol also indicated the increased cell viability in dopaminergic neurons, when cells were treated with 1-methyl-4-phenylridium, by regulating and inducing mitochondrial function and depolarization, and promoting autophagy.
4.6. 24(S)-Saringosterol
24(S)-Saringosterol or (3β)-stigmasta-5,28-diene-3,24-diol is a marine phytosterol commonly isolated from the edible seaweed Sargassum fusiforme. 24(S)-saringosterol was shown to be a potential anti-atherosclerotic natural agent by lowering cholesterol and a selective LXR-β agonist [75,76]. An in vivo study of APPswePS1DE9 mice treated with 24(S)-saringosterol for 10 weeks revealed a slowing of cognitive decline based on enhanced spatial and object memory assessments. It also reduced the in vivo expression of ionized calcium-binding adapter molecule 1 (Iba1), a marker for microglial activation and inflammation. No effect was detected on the amount of Aβ plaques, hence suggesting the neuroinflammatory effects of 24(S)-saringosterol in the deterrence of cognitive decline or limited penetration through BBB [77].
Dietary supplementation of S. fusiforme enriched in 24(S)-saringosterol in an AD mouse model indicated a reduction in hippocampal Aβ plaques and improvement in short-term memory. In vitro treatment with the same extract on mouse neuroblastoma (N2a) cells also exhibited a reduced secretion of Aβ plaques [78]. The reduction in the levels of Aβ plaques by dietary supplementation of S. fusiforme extracts enriched with phytosterols including 24(S)-saringosterol may indicate a synergistic model of the phytosterols in the prevention of neurodegenerative diseases.
4.7. 4,4-Dimethyl Phytosterols
4,4-Dimethyl phytosterols are a class of bioactive compounds having two methyl groups at the C-4 position of the aliphatic A-ring. Significant amounts of α-amyrin, β-amyrin, taraxerol, and lupeol are present in plants and vegetable oils. In scopolamine-induced cognitive impairment in mice, elevated levels of memory-related proteins in hippocampus were reported in the presence of α-amyrin and β-amyrin. Improvement in cognitive function was found to be induced through the activation of extracellular signal-regulated kinase (ERK) and glycogen synthase kinase-3β (GSK-3β). Additionally, β-amyrin and not α-amyrin displayed anti-AChE activity [79,80]. In scopolamine- and streptozotocin-induced memory deficit studies in mice, taraxerol also displayed anti-AChE activity through activation of the AChE receptor system [81]. Molecular docking studies confirmed high affinity of taraxerol for fibrils and Aβ [82]. Lupeol was found to inhibit BACE1 in both enzymatic and docking studies and is therefore a promising candidate for AD treatment [83,84]. The reduced levels of LDL-C were typically related to the prevalence of AD and PD [85]. Consequently, the anti-Alzheimer’s and anti-Parkinsonian potential of 4,4-dimethy sterols may also be linked to their cholesterol metabolism regulating activity [86].
In addition, the elimination of the misfolded proteins through autophagy is an important mechanism in preventing neurodegenerative diseases. It was found that β-amyrin participated in the LGG-1 (ubiquitin-like modifier involved in the formation of autophagosomes) related autophagy pathway by improving LGG-1 expression and exhibiting a protective effect on dopaminergic neurons by decreasing cell damage, and α-synuclein aggregation, which improves PD symptoms [87].
4.8. Ergosterol
Ergosterol is the most abundant fungal phytosterol bearing the Δ5,7 diene oxysterol skeleton [88]. In in vitro β- and γ-secretase assays utilizing N2a cells, ergosterol slightly decreased the β-secretase activity at 20–80 µM concentrations, while strongly inhibiting the γ-secretase activity at 40 µM [53].
These data suggest the promising potentials of the phytosterols as modulators of neurodegenerative diseases. The neuroprotective effects exhibited by the phytosterols are summarized in Table 1.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Neuroprotective mechanisms of the reported phytosterols.
| Phytosterol | Mode of Action | Study | References |
|---|---|---|---|
| β–Sitosterol | AChE and BChE inhibitory activity | in vivo (mice), in vitro, in silico | [45,46] |
| Increased levels of antioxidant enzymes by activating estrogen receptor/PI3-kinase pathway | in vitro (RAW 264.7; HT22) | [47,48] | |
| Anti-inflammatory | in vitro (BV12) | [59] | |
| Increase mitochondrial potential | in vitro (HT22) | [58] | |
| Stigmasterol | AChE inhibitory activity | in vitro | [60] |
| Reduced the β-secretase activity. Reduced the expression of all γ-secretase components Reduced the cholesterol and presenilin distribution in lipid rafts implicated in amyloidogenic APP cleavage. Decreased the BACE1 internalization to endosomal compartments, involved in APP β-secretase cleavage | in vivo (mice), in vitro (HT22) | [31,55] | |
| Decrease ROS | in vitro (SH-SY5Y) | [61,62] | |
| Anti-inflammatory | in vitro (mouse chondrocytes and human osteoarthritis chondrocytes) | [66] | |
| Brassicasterol | Marker in CSF of AD patients | cerebrospinal fluid (CSF) | [69] |
| Minor or no effect on Aβ secretion | in vivo (mice) | [31] | |
| Campesterol | Minor or no effect on Aβ secretion | in vivo (mice) | [31] |
| Lanosterol | Reduced the accumulations and cytotoxicity of Aβ aggregation through induction of co-chaperone and by promoting autophagy | in silico, in vitro (HeLa, PC12, HEK-293A), in vivo (mice) | [71,72,73,74] |
| 24(S)-Saringosterol | Reduced the in vivo expressions of Iba1 | in vivo (mice) | [77] |
| Dietary supplementation of S. fusiforme enriched in 24(S)-saringosterol | Reduced secretion of Aβ plaques. Improves memory in AD mice model | in vivo (mice), in vitro (N2a) | [78] |
| α-Amyrin | Elevated levels of memory related proteins through the activation of ERKGSK-3β | in vivo (mice) | [79] |
| β-Amyrin | Elevated levels of memory related proteins through the activation of ERKGSK-3β. AChE inhibitory activity | in vivo (mice) | [79,80] |
| Taraxerol | AChE inhibitory activity | in vivo (mice) | [83] |
| High affinity of taraxerol for fibrils and amyloid- β | in silico | [84] | |
| Lupeol | BACE-I inhibitory activity | in vitro, in silico | [85,86] |
| Ergosterol | Reduced the β- and γ-secretase activity | in vitro | [44] |
5. Physiological versus Pathological Features of Phytosterols
In diet, phytosterols primarily reduce blood cholesterol concentrations [89] by lowering serum LDL-cholesterol concentrations, with no effect on either serum HDL-cholesterol concentrations or triacylglycerol levels [90]. They are also anti-inflammatory and have physiological functions such as growth regulation and the promotion of protein synthesis, immune regulation, and hormone-like effects [91]. The hypocholesterolemic effect of 4-desmethyl sterols is well known [92], whereas 4,4-dimethylsterols (lupeol, α-amyrin, cycloartenol) have limited action on cholesterol reduction [93]. As phytosterols reduce the solubility of cholesterol, some other lipophilic compounds such as lipophilic antioxidant nutrients may also be displaced. Randomized trials have shown that phytosterols lower the blood concentrations of β-carotene (by about 25%), α-carotene (by 10%), and vitamin E (by 8%) [94,95], which protect the oxidation of LDL cholesterol. However, after the action of phytosterols, β-carotene was found to be reduced by 8–19%, while fat-soluble vitamins (A, D, E, K) remained unchanged [94]. Therefore, intake of food rich in carotenes may balance this side effect.
The most adversarial effect of phytosterols is seen in a rare inherited disease, sitosterolemia, described by tendon xanthoma and premature coronary disease [96]. These patients have mutated ABCG5 and ABCG8 transporters that cause reduced transport of phytosterols from enterocytes back into the intestinal lumen and reduced secretion into bile. Hence, these patients have increased phytosterol absorption (3–4 fold) and low biliary excretion, resulting in build-up of these compounds in plasma and tissues [97]. A relationship between increased plasma phytosterols (7–16%) and increased risk of coronary heart disease (CHD) has been reported [98]. However, various contradictory reports rule out any relation between phytosterols and the risk of incident CHD. Additionally, plasma concentrations of the main phytosterols (sitosterol and campesterol) can be used as bio markers for cardiometabolic risk, as moderately elevated plasma sitosterol, but not campesterol, may possibly indicate decreased risk for CHD [14]. Thus, the available data are contradictory and more in-depth studies are required to confirm whether phytosterols are friend or foe.
6. Safety Concerns for the Phytosterols
Even though intake of phytosterols (up to 3 g/day) is considered safe by the FDA, information on their toxicity is limited, including their indistinct anabolic effect [18]. No adverse effect of phytosterols on mental or cognitive activity has been reported yet. However, oxidized phytosterols were reported to exhibit neurotoxicity with glutamate excitotoxicity and ROS generation [87,99]. Several unfavorable effects of phytosterols on endothelium-dependent vasorelaxation in wt mice have been reported [100,101,102,103]. The types and concentrations of phytosterols or their structure may determine and influence the production of superoxides by endothelial cells [101,102,103]. Collectively, these studies indicated that phytosterols could modulate various endothelium-dependent processes, such as vasorelaxation, oxidative stress, ischemia–reperfusion, and neuroinflammation, which are key biological processes in the progression of CNS disorders. Therefore, depending on the nature, concentrations, and target cells, the phytosterols may have critical influences on neurodegenerative disorders [104].
7. Future Perspectives and Conclusions
Phytosterols are an important component of diet. They have wide applications in foods and cosmetics. To date, our knowledge has been limited to their cholesterol-lowering properties, protective effects mostly against cardiovascular diseases, and their anti-cancer and anti-inflammatory potential. Extensive research on the use of phytosterols for the preventive and therapeutic management of other diseases should be highly explored. The impact of phytosterols on the central nervous system is another exciting avenue of investigation. Phytosterols could cross the BBB through a less-known mechanism and become accumulated in the brain. The BBB is one of the most important and largest barriers among the three CNS barriers for the exchange of constituents between the blood and the CNS [105]. Any impairment in the brain endothelial cells may alter the normal function of adhesion molecules, chemotactic proteins, and angiogenic factors; thus, increased ROS production, infiltration of immune cells, and neuroinflammation could be the consequences in CNS disorders [106,107,108,109]. Moreover, normal functioning of various signaling pathways occurring at lipid rafts (cholesterol-rich domains), such as γ-aminobutyric acid and glutamate signaling, may perturb the synaptic vesicle turnover, protection of motor neurons by brain-derived neurotrophic factor (BDNF), functioning of the calcium channel, and calcium-dependent neurotransmitter release [110,111,112,113].
AD patients are shown to have compromised cholesterol turnover [114,115], where suppressed cholesterol biosynthesis may reduce the production of Aβ both in vitro and in vivo [68]. Likewise, a cholesterol-rich diet increased the generation of Aβ in mice [116,117,118]. The increased cellular cholesterol turnovers were in association with increased expression of the LXR-activated genes, which significantly amended cognitive performance in AD animal models [119,120,121].
Because the early stage of AD is linked to BBB dysfunction, the decreased concentrations of phytosterols in CSF could be used as a promising prognostic biomarker. Importantly, cholesterol is highly amyloidogenic, whereas the phytosterols are not. In fact, phytosterols significantly reduced brain Aβ levels and β- and γ-secretase activities in vivo, suggesting that a diet enriched in plant sterols might be beneficial for neurodegenerative diseases. Research on the optimal doses of phytosterols through oral administration would be essential. Because the consumed phytosterols would have limited intestinal absorption and crossing the BBB may take longer periods of ~6 weeks, higher doses may be required to capture the benefits [122]. Researchers have resorted to nanoencapsulation for improving the bioavailability of phytosterols in the food industry [123,124,125], but only for research purposes as a methodology of drug delivery. Thus, future studies should be carried out to explore the therapeutic and disease-specific mechanisms of phytosterols for their neuroprotective role in neurodegenerative diseases.
Author Contributions
Conceptualization, M.A.T. and S.S.A.A.; writing—original draft preparation, N.S. and M.A.T.; writing—review and editing, N.S., M.A.T. and S.S.A.A.; funding acquisition, S.S.A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Research Foundation of Korea and by the Korean Government (2020R1A2B5B01002463 and 2021R1A6A1A03038996).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- LaFerla, F.M.; Green, K.N.; Oddo, S. Intracellular amyloid-β in Alzheimer’s disease. Nat. Rev. Neurosci. 2007, 8, 499–509. [Google Scholar] [CrossRef]
- Minati, L.; Edginton, T.; Bruzzone, M.G.; Giaccone, G. Current concepts in Alzheimer’s disease: A multidisciplinary review. Am. J. Alzheimers Dis. Other Dement. 2009, 24, 95–121. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J. Alzheimer’s disease: Genes, proteins, and therapy. Physiol. Rev. 2001, 81, 741–766. [Google Scholar] [CrossRef] [PubMed]
- Armand, E.F.; Shantaram, M.; Nico, N.F.; Simon, F.N.; Paul, M.F. Potential of medicinal plant compounds to targeting Tau protein in the therapy of Alzheimer’s disease—A review. Biomedicine 2019, 39, 217–227. [Google Scholar] [CrossRef]
- Cui, X.; Lin, Q.; Liang, Y. Plant-derived antioxidants protect the nervous system from aging by inhibiting oxidative stress. Front. Aging Neurosci. 2020, 12, 209. [Google Scholar] [CrossRef] [PubMed]
- Murray, A.P.; Faraoni, M.B.; Castro, M.J.; Alza, N.P.; Cavallaro, V. Natural AChE inhibitors from plants and their contribution to Alzheimer’s disease therapy. Curr. Neuropharmacol. 2013, 11, 388–413. [Google Scholar] [CrossRef] [Green Version]
- Witter, S.; Witter, R.; Vilu, R.; Samoson, A. Medical plants and nutraceuticals for amyloid-β fibrillation inhibition. J. Alzheimers Dis. Rep. 2018, 2, 239–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, L.; Zhang, F.; Shen, M.; Jia, S.; Xie, J. Phytosterols suppress phagocytosis and inhibit inflammatory mediators via ERK pathway on LPS-triggered inflammatory responses in RAW264. 7 macrophages and the correlation with their structure. Foods 2019, 8, 582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piironen, V.; Lindsay, D.G.; Miettinen, T.A.; Toivo, J.; Lampi, A.M. Plant sterols: Biosynthesis, biological function and their importance to human nutrition. J. Sci. Food Agric. 2000, 80, 939–966. [Google Scholar] [CrossRef]
- Weihrauch, J.L.; Gardner, J.M. Sterol content of foods of plant origin. J. Am. Diet. Assoc. 1978, 73, 39–47. [Google Scholar] [CrossRef]
- Othman, R.A.; Myrie, S.B.; Jones, P.J. Non-cholesterol sterols and cholesterol metabolism in sitosterolemia. Atherosclerosis 2013, 231, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Ostlund Jr, R.E.; McGill, J.B.; Zeng, C.M.; Covey, D.F.; Stearns, J.; Stenson, W.F.; Spilburg, C.A. Gastrointestinal absorption and plasma kinetics of soy Δ5-phytosterols and phytostanols in humans. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E911–E916. [Google Scholar] [CrossRef]
- Von Bergmann, K.; Sudhop, T.; Lütjohann, D. Cholesterol and plant sterol absorption: Recent insights. Am. J. Cardiol. 2005, 96, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Escurriol, V.; Cofán, M.; Moreno-Iribas, C.; Larrañaga, N.; Martínez, C.; Navarro, C.; Rodriguez, L.; Gonzalez, C.A.; Corella, D.; Ros, E. Phytosterol plasma concentrations and coronary heart disease in the prospective Spanish EPIC cohort. J. Lipid Res. 2010, 51, 618–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morand, C.; Tomás Barberán, F.A. Contribution of plant food bioactives in promoting health effects of plant foods: Why look at interindividual variability? Eur. J. Nutr. 2019, 58, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Nashed, B.; Yeganeh, B.; HayGlass, K.T.; Moghadasian, M.H. Antiatherogenic effects of dietary plant sterols are associated with inhibition of proinflammatory cytokine production in Apo E-KO mice. J. Nutr. 2005, 135, 2438–2444. [Google Scholar] [CrossRef] [Green Version]
- Trautwein, E.A.; Vermeer, M.A.; Hiemstra, H.; Ras, R.T. LDL-cholesterol lowering of plant sterols and stanols—Which factors influence their efficacy? Nutrients 2018, 10, 1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopylov, A.T.; Malsagova, K.A.; Stepanov, A.A.; Kaysheva, A.L. Diversity of plant sterols metabolism: The impact on human health, sport, and accumulation of contaminating sterols. Nutrients 2021, 13, 1623. [Google Scholar] [CrossRef] [PubMed]
- Dutta, P.C. Phytosterols as Functional Food Components and Nutraceuticals, 1st ed.; CRC Press: Boca Raton, FL, USA, 2003; p. 96. [Google Scholar]
- Vanmierlo, T.; Weingärtner, O.; van der Pol, S.; Husche, C.; Kerksiek, A.; Friedrichs, S.; Sijbrands, S.; Steinbusch, H.; Grimm, M.; Hartmann, T.; et al. Dietary intake of plant sterols stably increases plant sterol levels in the murine brain. J. Lipid Res. 2012, 53, 726–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, R.A.; Nystrom, L.; Whitaker, B.D.; Winkler-Moser, J.K.; Baer, D.J.; Gebauer, S.K.; Hicks, K.B. Phytosterols and their derivatives: Structural diversity, distribution, metabolism, analysis, and health-promoting uses. Prog. Lipid Res. 2018, 70, 35–61. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lin, K.; Li, Y. Highlights to phytosterols accumulation and equilibrium in plants: Biosynthetic pathway and feedback regulation. Plant. Physiol. Biochem. 2020, 155, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Fricke, C.B.; Schrøder, M.; Poulsen, M.; von Bergmann, K.; Wester, I.; Knudsen, I.; Mortensen, A.; Lütjohann, D. Increased plant sterol and stanol levels in brain of Watanabe rabbits fed rapeseed oil derived plant sterol or stanol esters. Br. J. Nutr. 2007, 98, 890–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, P.J.; Lutjohann, D.; Abildayeva, K.; Vanmierlo, T.; Plosch, T.; Plat, J.; von Bergmann, K.; Groen, A.; Ramaekers, F.; Kuipers, F.; et al. Dietary plant sterols accumulate in the brain. Biochim. Biophys. Acta 2006, 1761, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Dietschy, J.M.; Turley, S.D. Thematic review series: Brain lipids. Cholesterol metabolism in the central nervous system during early development and in the mature animal. J. Lipid Res. 2004, 45, 1375–1397. [Google Scholar] [CrossRef] [Green Version]
- Björkhem, I. Crossing the barrier: Oxysterols as cholesterol transporters and metabolic modulators in the brain. J. Intern. Med. 2006, 260, 493–508. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Muneton, S.; Sjövall, J.; Jovanovic, J.N.; Griffiths, W.J. The effect of 24S-hydroxycholesterol on cholesterol homeostasis in neurons: Quantitative changes to the cortical neuron proteome. J. Proteome Res. 2008, 7, 1606–1614. [Google Scholar] [CrossRef] [Green Version]
- Van Kampen, J.M.; Robertson, H.A. The BSSG rat model of Parkinson’s disease: Progressing towards a valid, predictive model of disease. EPMA J. 2017, 8, 261–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mongrand, S.; Stanislas, T.; Bayer, E.M.; Lherminier, J.; Simon-Plas, F. Membrane rafts in plant cells. Trends Plant. Sci. 2010, 15, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Klaikeaw, N.; Wongphoom, J.; Werawatganon, D.; Chayanupatkul, M.; Siriviriyakul, P. Anti-inflammatory and anti-oxidant effects of Aloe vera in rats with non-alcoholic steatohepatitis. World J. Hepatol. 2020, 12, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Burg, V.K.; Grimm, H.S.; Rothhaar, T.L.; Grösgen, S.; Hundsdörfer, B.; Haupenthal, V.J.; Zimmer, V.; Mett, J.; Weingartner, O.; Laufs, U.; et al. Plant sterols the better cholesterol in Alzheimer’s disease? A mechanistical study. J. Neurosci. 2013, 33, 16072–16087. [Google Scholar] [CrossRef] [PubMed]
- Panchal, M.; Loeper, J.; Cossec, J.; Perruchini, C.; Lazar, A.; Pompon, D.; Duyckaerts, C. Enrichment of cholesterol in microdissected Alzheimer’s disease senile plaques as assessed by mass spectrometry. J. Lipid Res. 2010, 51, 598–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandiah, N.; Feldman, H. Therapeutic potential of statins in Alzheimer’s disease. J. Neurol. Sci. 2009, 283, 230–234. [Google Scholar] [CrossRef]
- Fonseca, A.; Resende, R.; Oliveira, C.; Pereira, C. Cholesterol and statins in Alzheimer’s disease: Current controversies. Exp. Neurol. 2010, 223, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Puglielli, L.; Tanzi, R.; Kovacs, D. Alzheimer’s disease: The cholesterol connection. Nat. Neurosci. 2003, 6, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.; Emmerling, M.; Bisgaier, C.; Essenburg, A.; Lampert, H.; Drumm, D.; Roher, A. Elevated low-density lipoprotein in Alzheimer’s disease correlates with brain Aβ1-42 levels. Biochem. Biophys. Res. Commun. 1998, 252, 711–715. [Google Scholar] [CrossRef] [PubMed]
- Knebl, J.; DeFazio, P.; Clearfield, M.; Little, L.; McConathy, W.; McPherson, R.; Lacko, A. Plasma lipids and cholesterol esterification in Alzheimer’s disease. Mech. Ageing Dev. 1994, 73, 69–77. [Google Scholar] [CrossRef]
- Tang, B.L. Neuronal protein trafficking associated with Alzheimer disease: From APP and BACE1 to glutamate receptors. Cell Adh. Migr. 2009, 3, 118–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalvodova, L.; Kahya, N.; Schwille, P.; Ehehalt, R.; Verkade, P.; Drechsel, D.; Simons, K. Lipids as modulators of proteolytic activity of BACE: Involvement of cholesterol, glycosphingolipids, and anionic phospholipids in vitro. J. Biol. Chem. 2005, 280, 36815–36823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beel, A.J.; Sakakura, M.; Barrett, P.J.; Sanders, C.R. Direct binding of cholesterol to the amyloid precursor protein: An important interaction in lipid–Alzheimer’s disease relationships? Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2010, 1801, 975–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wu, F.; Shi, C. Substitution of membrane cholesterol with β-sitosterol promotes nonamyloidogenic cleavage of endogenous amyloid precursor protein. Neuroscience 2013, 247, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Valerio, M.; Liu, H.B.; Heffner, R.; Zivadinov, R.; Ramanathan, M.; Weinstock-Guttman, B.; Awad, A.B. Phytosterols ameliorate clinical manifestations and inflammation in experimental autoimmune encephalomyelitis. Inflamm. Res. 2011, 60, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Berghoff, S.A.; Gerndt, N.; Winchenbach, J.; Stumpf, S.K.; Hosang, L.; Odoardi, F. Dietary cholesterol promotes repair of demy-elinated lesions in the adult brain. Nat. Commun. 2017, 8, 14241. [Google Scholar] [CrossRef] [PubMed]
- Saher, G.; Rudolphi, F.; Corthals, K.; Ruhwedel, T.; Schmidt, K.F.; Lowel, S.; Dibaj, P.; Barrette, B.; Mobius, W.; Nave, K. Therapy of Pelizaeus-Merzbacher disease in mice by feeding a cholesterol-enriched diet. Nat. Med. 2012, 18, 1130–1135. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, M.; Junaid, M.; Ullah, F.; Subhan, F.; Sadiq, A.; Ali, G.; Ovais, M.; Shahid, M.; Ahmad, A.; Wadood, A.; et al. Anti-Alzheimer’s studies on β-sitosterol isolated from Polygonum hydropiper L. Front. Pharmacol. 2017, 8, 697. [Google Scholar] [CrossRef] [Green Version]
- Bari, W.U.; Zahoor, M.; Zeb, A.; Khan, I.; Nazir, Y.; Khan, A.; Ur Rehman, N.; Ullah, R.; Shahat, A.; Mahmood, H.M. Anticholinesterase, antioxidant potentials, and molecular docking studies of isolated bioactive compounds from Grewia optiva. Int. J. Food Prop. 2019, 22, 1386–1396. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Wu, F.; Zhu, X.; Xu, J. Incorporation of β-sitosterol into the membrane increases resistance to oxidative stress and lipid peroxidation via estrogen receptor-mediated PI3K/GSK3β signalling. Biochim. Biophys. Acta 2013, 1830, 2538–2544. [Google Scholar] [CrossRef] [PubMed]
- Vivancos, M.; Moreno, J.J. β-Sitosterol modulates antioxidant enzyme response in RAW 264.7 macrophages. Free Radic. Biol. Med. 2005, 39, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Fan, X.; Gabbi, C.; Yakimchuk, K.; Parini, P.; Warner, M.; Gustafsson, J.A. Liver X receptor β (LXRβ): A link between β-sitosterol and amyotrophic lateral sclerosis-Parkinson’s dementia. Proc. Natl. Acad. Sci. USA 2008, 105, 2094–2099. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.Y.; Li, L.; Hao, Q.M.; Qin, Y.; Ma, C.S. β-Sitosterol treatment attenuates cognitive deficits and prevents amyloid plaque deposition in amyloid protein precursor/presenilin 1 mice. Korean J. Physiol. Pharmacol. 2020, 24, 39–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Runz, H.; Rietdorf, J.; Tomic, I.; de Bernard, M.; Beyreuther, K.; Pepperkok, R.; Hartmann, T. Inhibition of intracellular cholesterol transport alters presenilin localization and amyloid precursor protein processing in neuronal cells. J. Neurosci. 2002, 22, 1679–1689. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Kulas, J.A.; Wang, C.; Holtzman, D.M.; Ferris, H.A.; Hansen, S.B. Regulation of beta-amyloid production in neurons by astrocyte-derived cholesterol. Proc. Nat. Acad. Sci. USA 2021, 118, e2102191118. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Callaghan, D.; Jones, A.; Walker, D.G.; Lue, L.F.; Beach, T.G.; Sue, L.; Woulfe, J.; Xu, H.; Stanimirovic, D.; et al. Cholesterol retention in Alzheimer’s brain is responsible for high β-and γ-secretase activities and Aβ production. Neurobiol. Dis. 2008, 29, 422–437. [Google Scholar] [CrossRef] [Green Version]
- Clifton, P. Lowering cholesterol: A review on the role of plant sterols. Aust. Fam. Physician 2009, 38, 218–221. [Google Scholar] [PubMed]
- Shi, C.; Liu, J.; Wu, F.; Zhu, X.; Yew, D.T.; Xu, J. β-sitosterol inhibits high cholesterol-induced platelet β-amyloid release. J. Bioenerg. Biomembr. 2011, 43, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Schon, E.A.; Manfredi, G. Neuronal degeneration and mitochondrial dysfunction. J. Clin. Investig. 2003, 111, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.H. Role of mitochondria in neurodegenerative diseases: Mitochondria as a therapeutic target in Alzheimer’s disease. CNS Spect. 2009, 8 (Suppl. 7), 8–13. [Google Scholar]
- Shi, C.; Wu, F.; Xu, J. Incorporation of β-sitosterol into mitochondrial membrane enhances mitochondrial function by promoting inner mitochondrial membrane fluidity. J. Bioenerg. Biomembr. 2013, 45, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Gao, L.; Hou, W.; Wu, J. β-Sitosterol alleviates inflammatory response via inhibiting the activation of ERK/p38 and NF-κB pathways in LPS-exposed BV2 cells. BioMed Res. Int. 2020, 2020, 7532306. [Google Scholar] [CrossRef]
- Sultana, N.; Khalid, A. Phytochemical and enzyme inhibitory studies on indigenous medicinal plant Rhazya stricta. Nat. Prod. Res. 2010, 24, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Weon, J.B.; Ma, C.J. Neuroprotective activity of phytosterols isolated from Artemisia apiacea. Korean J. Pharmacogn. 2014, 45, 214–219. [Google Scholar]
- Pratiwi, R.; Nantasenamat, C.; Ruankham, W.; Suwanjang, W.; Prachayasittikul, V.; Prachayasittikul, S.; Phopin, K. Mechanisms and neuroprotective activities of stigmasterol against oxidative stress-induced neuronal cell death via sirtuin family. Front. Nutr. 2021, 8, 648995. [Google Scholar] [CrossRef]
- Donmez, G. The neurobiology of sirtuins and their role in neurodegeneration. Trends Pharmacol. Sci. 2012, 33, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Braidy, N.; Jayasena, T.; Poljak, A.; Sachdev, P.S. Sirtuins in cognitive ageing and Alzheimer’s disease. Curr. Opin. Psychiatry 2012, 25, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Hanson, P.S.; Morris, C.M. SIRT1 ameliorates oxidative stress induced neural cell death and is down-regulated in Parkinson’s disease. BMC Neurosci. 2017, 18, 46. [Google Scholar] [CrossRef] [PubMed]
- Gabay, O.; Sanchez, C.; Salvat, C.; Chevy, F.; Breton, M.; Nourissat, G.; Wolf, C.; Jacques, C.; Berenbaum, F. Stigmasterol: A phytosterol with potential anti-osteoarthritic properties. Osteoarthr. Cartil. 2010, 18, 106–116. [Google Scholar] [CrossRef]
- Haque, M.N.; Moon, I.S. Stigmasterol upregulates immediate early genes and promotes neuronal cytoarchitecture in primary hippocampal neurons as revealed by transcriptome analysis. Phytomedicine 2018, 46, 164–175. [Google Scholar] [CrossRef]
- Shafaati, M.; Marutle, A.; Pettersson, H.; Lövgren-Sandblom, A.; Olin, M.; Pikuleva, I. Marked accumulation of 27- hydroxycholesterol in the brains of Alzheimer’s patients with the Swedish APP 670/671 mutation. J. Lipid Res. 2011, 52, 1004–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanmierlo, T.; Popp, J.; Kölsch, H.; Friedrichs, S.; Jessen, F.; Stoffel-Wagner, B.; Bertsch, T.; Hartmann, T.; Maier, W.; von Bergmann, K.; et al. The plant sterol brassicasterol as additional CSF biomarker in Alzheimer’s disease. Acta Psychiatr. Scand. 2011, 124, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Pei, H.; Ma, X.; Pan, Y.; Han, T.; Lu, Z.; Wu, R.; Cao, X.; Zheng, J. Separation and purification of lanosterol, dihydrolanosterol, and cholesterol from lanolin by high-performance counter-current chromatography dual-mode elution method. J. Sep. Sci. 2019, 42, 2171–2178. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Yang, Z.; Tian, X.; Chen, L.; Lee, S.; Huynh, T.; Ge, C.; Zhou, R. Lanosterol disrupts the aggregation of amyloid-β peptides. ACS Chem. Neurosci. 2019, 10, 4051–4060. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.-D.; Wang, J.; Chen, X.-J.; Yan, Y.-B. Lanosterol modulates proteostasis via dissolving cytosolic sequestosomes/aggresome-like induced structures. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118617. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, A.; Amanullah, A.; Mishra, R.; Kumar, A.; Mishra, A. Lanosterol suppresses the aggregation and cytotoxicity of misfolded proteins linked with neurodegenerative diseases. Mol. Neurobiol. 2018, 55, 1169–1182. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.; Jackson-Lewis, V.; Wong, L.C.; Shui, G.H.; Goh, A.X.; Kesavapany, S.; Jenner, A.M.; Fivaz, M.; Przedborski, S.; Wenk, M.R. Lanosterol induces mitochondrial uncoupling and protects dopaminergic neurons from cell death in a model for Parkinson’s disease. Cell Death Differ. 2012, 19, 416–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Liu, J.; Fu, Z.; Ye, C.; Zhang, R.; Song, Y.; Zhang, Y.; Li, H.; Ying, H.; Liu, H. 24(S)-Saringosterol from dibble marine seaweed Sargassum fusiforme is a novel selectve LXRβ agonist. J. Agric. Food Chem. 2014, 62, 6130–6137. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Niu, Z.; Wang, B.; Zhao, S.; Sun, C.; Wu, Y.; Li, Y.; Ying, H.; Liu, H. Saringosterol from sargassum fusiforme modulates cholesterol metabolism and alleviates atherosclerosis in ApoE-deficient mice. Mar. Drugs 2021, 19, 485. [Google Scholar] [CrossRef] [PubMed]
- Martens, N.; Schepers, M.; Zhan, N.; Leijten, F.; Voortman, G.; Tiane, A.; Rombaut, B.; Poisquet, J.; van de Sande, N.; Kerksiek, A.; et al. 24(S)-Saringosterol prevents cognitive decline in a mouse model for Alzheimer’s disease. Mar. Drugs 2021, 19, 190. [Google Scholar] [CrossRef]
- Bogie, J.; Hoeks, C.; Schepers, M.; Tiane, A.; Cuypers, A.; Leijten, F.; Chintapakorn, Y.; Suttiyut, T.; Pornpakakul, S.; Struik, D.; et al. Dietary Sargassum fusiforme imprives memory and reduces amyloid plaque load in an Alzheimer’s disease mouse model. Sci. Rep. 2019, 9, 4908. [Google Scholar]
- Park, S.J.; Ahn, J.Y.; Oh, S.R.; Lee, Y.; Kwon, G.; Woo, H.; Lee, H.E.; Jang, D.S.; Ryu, J.H. Amyrin attenuates scopolamine-induced cognitive impairment in mice. Biol. Pharm. Bull. 2014, 37, 1207–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, S.; Loesche, A.; Lucas, S.D.; Sommerwerk, S.; Serbian, I.; Siewert, B.; Pianowski, E.; Csuk, R. Converting maslinic acid into an effective inhibitor of acetylcholinesterases. Eur. J. Med. Chem. 2015, 103, 438–445. [Google Scholar] [CrossRef]
- Berté, T.E.; Dalmagro, A.P.; Zimath, P.L.; Gonçalves, A.E.; Meyre-Silva, C.; Bürger, C.; Weber, C.J.; Dos Santos, D.A.; Cechinel-Filho, V.; de Souza, M.M. Taraxerol as a possible therapeutic agent on memory impairments and Alzheimer’s disease: Effects against scopolamine and streptozotocin-induced cognitive dysfunctions. Steroids 2018, 132, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Ngo, S.T.; Li, M.S. Top-leads from natural products for treatment of Alzheimer’s disease: Docking and molecular dynamics study. Mol. Simul. 2013, 39, 279. [Google Scholar] [CrossRef]
- Hosen, S.M.; Rubayed, M.; Dash, R.; Junaid, M.; Mitra, S.; Alam, M.; Dey, R. Prospecting and structural insight into the binding of novel plant-derived molecules of Leea indica as inhibitors of BACE1. Curr. Pharm. Des. 2018, 24, 3972–3979. [Google Scholar] [CrossRef] [Green Version]
- Koirala, P.; Su, H.S.; Jung, H.A.; Choi, J.S. Comparative molecular docking studies of lupeol and lupenone isolated from Pueraria lobata that inhibits BACE1: Probable remedies for Alzheimer’s disease. Asian Pac. J. Trop. Med. 2017, 10, 1117–1122. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Chen, H.; Miller, W.C.; Mailman, R.B.; Woodard, J.L.; Chen, P.C.; Dong, X.; Murrow, R.W.; Wang, Y.Z.; Poole, C. Lower low density lipid cholesterol levels are associated with Parkinson’s disease. Mov. Disord. 2007, 22, 377–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Liu, R.; Chang, M.; Jin, Q.; Zhang, H.; Wang, X. Health benefits of 4, 4-dimethyl phytosterols: An exploration beyond 4-desmethyl phytosterols. Food Funct. 2020, 11, 93–110. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.-C.; Chang, C.-H.; Liao, H.-C. Anti-Parkinsonian effects of β-amyrin are regulated via LGG-1 involved autophagy pathway in Caenorhabditis elegans. Phytomedicine 2017, 36, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.L. The multifunctional fungal ergosterol. mBio 2018, 9, e01755-18. [Google Scholar] [CrossRef] [Green Version]
- Brufau, G.; Canela, M.A.; Rafecas, M. Phytosterols: Physiologic and metabolic aspects related to cholesterol-lowering properties. Nutr. Res. 2008, 28, 217–225. [Google Scholar] [CrossRef]
- Miettinen, T.A.; Vuoristo, M.; Nissinen, M.; Jarvinen, H.J.; Gylling, H. Serum, biliary, and fecal cholesterol and plant sterols in colectomized patients before and during consumption of stanol ester margarine. Am. J. Clin. Nutr. 2000, 71, 1095–1102. [Google Scholar] [CrossRef]
- Xinmei, X.; Ning, X.; Jianbin, H. Physiological function of phytosterol and its application. Anim. Husb. Feed Sci. 2015, 7, 67–69. [Google Scholar]
- Katan, M.B.; Grundy, S.M.; Jones, P.; Law, M.; Miettinen, T.A.; Paoletti, R.; Stresa Workshop Participants. Efficacy and safety of plant stanols and sterols in the management of blood cholesterol levels. Mayo Clin. Proc. 2003, 78, 965–978. [Google Scholar] [CrossRef] [Green Version]
- Sierksma, A.; Weststrate, J.A.; Meijer, G.W. Spreads enriched with plant sterols, either esterified 4,4 dimethyl-sterols or free 4-desmethylsterols, and plasma total- and LDL-cholesterol concentrations. Br. J. Nutr. 1999, 82, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, H.F.; Weststrate, J.A.; van Vliet, T.; Meijer, G.W. Spreads enriched with three different levels of vegetable oil sterols and the degree of cholesterol lowering in normocholesterolaemic and mildly hypercholesterolaemic subjects. Eur. J. Clin. Nutr. 1999, 53, 319–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallikainen, M.A.; Sarkkinen, E.S.; Uusitupa, M.I. Effects of low-fat stanol ester enriched margarine on concentrations of serum carotenoids in subjects with elevated serum cholesterol concentrations. Eur. J. Clin. Nutr. 1999, 53, 966–969. [Google Scholar] [CrossRef]
- Salen, G.; Shefer, S.; Nguyen, L.; Ness, G.C.; Tint, G.S.; Shore, V. Sitosterolemia. J. Lipid Res. 1992, 33, 945–955. [Google Scholar] [CrossRef]
- Lütjohann, D.; Björkhem, I.; Ose, L. Phytosterolemia in a Norwegian family: Diagnosis and characterization of the first Scandinavian case. Scand. J. Clin. Lab. Investig. 1996, 56, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Assmann, G.; Cullen, P.; Erbey, J.; Ramey, D.R.; Kannenberg, F.; Schulte, H. Plasma sitosterol elevations are associated with an increased incidence of coronary events in men: Results of a nested case control analysis of the prospective cardiovascular munster (PROCAM) study. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Panov, A.; Kubalik, N.; Brooks, B.R.; Shaw, C.A. In vitro effects of cholesterol β-D-glucoside, cholesterol and cycad phytosterol glucosides on respiration and reactive oxygen species generation in brain mitochondria. J. Membr. Biol. 2010, 237, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Tabata, R.C.; Wilson, J.M.; Ly, P.; Zwiegers, P.; Kwok, D.; Van Kampen, J.M.; Cashman, N.; Shaw, C. Chronic exposure to dietary sterol glucosides is neurotoxic to motor neurons and induces an ALS-PDC phenotype. Neuromolecular Med. 2008, 10, 24–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weingartner, O.; Lutjohann, D.; Ji, S.; Weisshoff, N.; List, F.; Sudhop, T.; von Bergmann, K.; Gertz, K.; Konig, J.; Schafers, H.J.; et al. Vascular effects of diet supplementation with plant sterols. J. Am. Coll. Cardiol. 2008, 51, 1553–1561. [Google Scholar] [CrossRef] [Green Version]
- Weingartner, O.; Ulrich, C.; Lutjohann, D.; Ismail, K.; Schirmer, S.H.; Vanmierlo, T.; Bohm, M.; Laufs, U. Differential effects on inhibition of cholesterol absorption by plant stanol and plant sterol esters in apoE-/- mice. Cardiovasc. Res. 2011, 90, 484–492. [Google Scholar] [CrossRef] [Green Version]
- Nam, Y.; Lee, D. Ameliorating effects of constituents from Cortex Acanthopanacis Radicis on memory impairment in mice induced by scopolamine. J. Tradit. Chin. Med. 2014, 34, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Chen, Z.-Y.; Wong, S.-L.; Liu, J.; Liang, Y.-T.; Lau, C.-W.; Lee, H.-K.; Huang, Y.; Tsang, S.-Y. β-Sitosterol oxidation products attenuate vasorelaxation by increasing reactive oxygen species and cyclooxygenase-2. Cardiovasc. Res. 2013, 97, 520–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanmierlo, T.; Bogie, J.F.; Mailleux, J.; Vanmol, J.; Lütjohann, D.; Mulder, M.; Hendriks, J.J. Plant sterols: Friend or foe in CNS disorders? Prog. Lipid Res. 2015, 58, 26–39. [Google Scholar] [CrossRef]
- Abbott, N.J.; Patabendige, A.A.; Dolman, D.E.; Yusof, S.R.; Begley, D.J. Structure and function of the blood–brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Zlokovic, B.V. The blood–brain barrier in health and chronic neurodegenerative disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, D.A.; Jin, K. From angiogenesis to neuropathology. Nature 2005, 438, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Uttara, B.; Singh, A.V.; Zamboni, P.; Mahajan, R.T. Oxidative stress and neurodegenerative diseases: A review of upstream and downstream antioxidant therapeutic options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grammas, P.; Martinez, J.; Miller, B. Cerebral microvascular endothelium and the pathogenesis of neurodegenerative diseases. Expert Rev. Mol. Med. 2011, 13, e19. [Google Scholar] [CrossRef]
- Davies, A.; Douglas, L.; Hendrich, J.; Wratten, J.; Tran Van Minh, A.; Foucault, I.; Koch, D.; Pratt, W.; Saibil, H.; Dolphin, A. The calcium channel α2δ-2 subunit partitions with CaV2.1 into lipid rafts in cerebellum: Implications for localization and function. J. Neurosci. 2006, 26, 8748–8757. [Google Scholar] [CrossRef] [PubMed]
- Mojsilovic-Petrovic, J.; Jeong, G.B.; Crocker, A.; Arneja, A.; David, S.; Russell, D.; Kalb, R. Protecting motor neurons from toxic insult by antagonism of adenosine A2a and Trk receptors. J. Neurosci. 2006, 26, 9250–9263. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Serwanski, D.R.; Miralles, C.P.; Bahr, B.A.; De Blas, A.L. Two pools of Triton X-100-insoluble GABA(A) receptors are present in the brain, one associated to lipid rafts and another one to the postsynaptic GABAergic complex. J. Neurochem. 2007, 102, 1329–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasser, C.R.; Ertunc, M.; Liu, X.; Kavalali, E.T. Cholesterol-dependent balance between evoked and spontaneous synaptic vesicle recycling. J. Physiol. 2007, 579, 413–429. [Google Scholar] [CrossRef] [PubMed]
- Kölsch, H.; Heun, R.; Jessen, F.; Popp, J.; Hentschel, F.; Maier, W.; Lütjohann, D. Alterations of cholesterol precursor levels in Alzheimer’s disease. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2010, 1801, 945–950. [Google Scholar] [CrossRef]
- Fassbender, K.; Simons, M.; Bergmann, C.; Stroick, M.; Lutjohann, D.; Keller, P.; Runz, H.; Kuhl, S.; Bertsch, T.; von Bergmann, K.; et al. Simvastatin strongly reduces levels of Alzheimer’s disease beta-amyloid peptides Abeta 42 and Abeta 40 in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2001, 98, 5856–5861. [Google Scholar] [CrossRef] [Green Version]
- Thirumangalakudi, L.; Prakasam, A.; Zhang, R.; Bimonte-Nelson, H.; Sambamurti, K.; Kindy, M.S.; Bhat, N.R. High cholesterol-induced neuroinflammation and amyloid precursor protein processing correlate with loss of working memory in mice. J. Neurochem. 2008, 106, 475–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julien, C.; Tremblay, C.; Phivilay, A.; Berthiaume, L.; Emond, V.; Julien, P.; Calon, F. Highfat diet aggravates amyloid-beta and tau pathologies in the 3xTg-AD mouse model. Neurobiol. Aging 2010, 31, 1516–1531. [Google Scholar] [CrossRef]
- Knight, E.M.; Martins, I.V.; Gumusgoz, S.; Allan, S.M.; Lawrence, C.B. High-fat diet induced memory impairment in triple-transgenic Alzheimer’s disease (3xTgAD) mice is independent of changes in amyloid and tau pathology. Neurobiol. Aging 2014, 35, 1821–1832. [Google Scholar] [CrossRef]
- Riddell, D.R.; Zhou, H.; Comery, T.A.; Kouranova, E.; Lo, C.F.; Warwick, H.K.; Ring, R.; Kirksey, Y.; Aschmies, S.; Xu, J.; et al. The LXR agonist TO901317 selectively lowers hippocampal Abeta42 and improves memory in the Tg2576 mouse model of Alzheimer’s disease. Mol. Cell. Neurosci. 2007, 34, 621–628. [Google Scholar] [CrossRef]
- Jiang, Q.; Lee, C.Y.; Mandrekar, S.; Wilkinson, B.; Cramer, P.; Zelcer, N.; Mann, K.; Lamb, B.; Wilson, T.; Collins, J.; et al. ApoE promotes the proteolytic degradation of Abeta. Neuron 2008, 58, 681–693. [Google Scholar] [CrossRef] [Green Version]
- Vanmierlo, T.; Rutten, K.; Dederen, J.; Bloks, V.W.; van Vark-van der Zee, L.C.; Kuipers, F.; Kiliaan, A.; Blokland, A.; Sijbrands, E.; Steinbusch, H.; et al. Liver X receptor activation restores memory in aged AD mice without reducing amyloid. Neurobiol. Aging 2011, 32, 1262–1272. [Google Scholar] [CrossRef]
- Mohammadi, M.; Jafari, S.M.; Hamishehkar, H.; Ghanbarzadeh, B. Phytosterols as the core or stabilizing agent in different nanocarriers. Trends Food Sci. Technol. 2020, 101, 73–88. [Google Scholar] [CrossRef]
- Tolve, R.; Cela, N.; Condelli, N.; Di Cairano, M.; Caruso, M.C.; Galgano, F. Microencapsulation as a tool for the formulation of functional foods: The phytosterols’ case study. Foods 2020, 9, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joye, I.J.; Davidov-Pardo, G.; McClements, D.J. Nanotechnology for increased micronutrient bioavailability. Trends Food Sci. Technol. 2014, 40, 168–182. [Google Scholar] [CrossRef]
Figure 1.
A simplified overview of the biosynthetic pathway of the major phytosterols in plants.

Figure 2.
Pathway of dietary phytosterol assimilation in humans. Phytosterols and dietary cholesterol are mainly absorbed in the intestine through NPC1L1 transporter. After absorption, sterols are esterified by ACAT and transported to the liver in the form of chylomicrons. The unesterified sterols are pumped out by ABCG5/ABCG8 transporters. The plasma efflux of sterols is regulated by ABCA1, which is involved in the assembly of HDL-like particles with the assimilated sterols. In liver, phytosterols stimulate LXR receptors regulating ApoE expression required for HDL and LDL assembly and uptake. Stimulated LXR receptors upregulate ABCG5/G8 transporters and augment sterol absorption. The exported HDL-like particles containing phytosterols are taken up by SR-IB receptors expressed on the liver, adrenal, and brain surface. SR-B1 has a crucial role in the flux of phytosterols across the BBB. ABCA1/ABCG1 transporters present on astrocytes and the basolateral side of the cerebral endothelium also assist in the transfer of phytosterols. Inside the brain, PS exert a positive effect on brain function by escalating A-β plaque clearance, increasing re-myelination besides reducing neuro inflammation and BACE 1 activity. Abbreviations: ABC, ATP-binding cassette transporters; A-β, Amyloid-β; ACAT, Acyl-coenzyme A cholesterol acyl transferase; ApoE, Apolipoprotein E; BACE1, β-Secretase 1; HDL, High-density lipoprotein; SR-B1, Scavenger receptor class B type 1; LDL, Low-density lipoprotein; LXR, Liver X receptor; NPC1L1, Niemann-Pick C1 like 1 protein; PS, Phytosterols.
Figure 2.
Pathway of dietary phytosterol assimilation in humans. Phytosterols and dietary cholesterol are mainly absorbed in the intestine through NPC1L1 transporter. After absorption, sterols are esterified by ACAT and transported to the liver in the form of chylomicrons. The unesterified sterols are pumped out by ABCG5/ABCG8 transporters. The plasma efflux of sterols is regulated by ABCA1, which is involved in the assembly of HDL-like particles with the assimilated sterols. In liver, phytosterols stimulate LXR receptors regulating ApoE expression required for HDL and LDL assembly and uptake. Stimulated LXR receptors upregulate ABCG5/G8 transporters and augment sterol absorption. The exported HDL-like particles containing phytosterols are taken up by SR-IB receptors expressed on the liver, adrenal, and brain surface. SR-B1 has a crucial role in the flux of phytosterols across the BBB. ABCA1/ABCG1 transporters present on astrocytes and the basolateral side of the cerebral endothelium also assist in the transfer of phytosterols. Inside the brain, PS exert a positive effect on brain function by escalating A-β plaque clearance, increasing re-myelination besides reducing neuro inflammation and BACE 1 activity. Abbreviations: ABC, ATP-binding cassette transporters; A-β, Amyloid-β; ACAT, Acyl-coenzyme A cholesterol acyl transferase; ApoE, Apolipoprotein E; BACE1, β-Secretase 1; HDL, High-density lipoprotein; SR-B1, Scavenger receptor class B type 1; LDL, Low-density lipoprotein; LXR, Liver X receptor; NPC1L1, Niemann-Pick C1 like 1 protein; PS, Phytosterols.

Figure 3.
(A) Membrane cholesterol favors β-secretase cleavage of APP by direct binding to the C-terminal transmembrane domain of APP, generating neurotoxic Aβ and less sAPP α. (B) Substitution of membrane cholesterol with β-sitosterol promotes the re-distribution of APP in non-raft region and non-amyloidogenic processing, generating less Aβ and more of neuroprotective sAPP α. Abbreviations: Aβ: beta amyloid; APP: amyloid precursor protein; sAPP α: soluble alpha-amyloid precursor protein.
Figure 3.
(A) Membrane cholesterol favors β-secretase cleavage of APP by direct binding to the C-terminal transmembrane domain of APP, generating neurotoxic Aβ and less sAPP α. (B) Substitution of membrane cholesterol with β-sitosterol promotes the re-distribution of APP in non-raft region and non-amyloidogenic processing, generating less Aβ and more of neuroprotective sAPP α. Abbreviations: Aβ: beta amyloid; APP: amyloid precursor protein; sAPP α: soluble alpha-amyloid precursor protein.

Figure 4.
Structure of the phytosterols with potential activities against neurodegenerative diseases.
Figure 4.
Structure of the phytosterols with potential activities against neurodegenerative diseases.

Figure 5.
Phytosterols exert a multitarget approach to ameliorate symptoms of AD. (A) Some prevent amyloid-beta aggregation by inhibiting cleavage of the amyloid precursor protein (APP) by β-secretase (BACE-I). This causes a shift in the non-amyloidogenic pathway and reduces the levels of Aβ produced. (B) Aβ can self-aggregate to form oligomers and eventually amyloid plaques. Some phytosterols are able to inhibit the formation of amyloid plaques by binding to Aβ, inhibiting aggregation, and thereby promoting the formation of nontoxic oligomers. Toxic Aβ monomers and oligomers have been shown to induce microglial activation and proliferation. Activated microglia secrete pro-inflammatory cytokines such as IL-1β and IL-6. (C) Some phytosterols have been shown to reduce the levels of these cytokines. Some phytosterols reduce oxidative stress by increasing the levels of antioxidant enzymes and reducing lipid peroxidation. (D) Acetylcholine (ACh), a neurotransmitter essential for processing memory and learning, is decreased in both concentration and function in AD. Decreased levels of ACh can be restored by anticholinesterase activity of various phytosterols. (E) ROS irreversibly oxidize DNA and are important mediators of Aβ-induced neuronal cell death in the development of AD. Abbreviations: APP: Amyloid Precursor Protein; AChE: Acetyl Cholinesterase Enzyme; BACE 1: Beta-Secretase 1; BChE: Butyl Cholinesterase Enzyme; Bcl-2: B-Cell Lymphoma 2; CAT: Catalase; GPX: Glutathione Peroxidase; HO 1: Heme Oxygenase; ROS: Reactive Oxygen Species; SOD: Superoxide Dismutase.
Figure 5.
Phytosterols exert a multitarget approach to ameliorate symptoms of AD. (A) Some prevent amyloid-beta aggregation by inhibiting cleavage of the amyloid precursor protein (APP) by β-secretase (BACE-I). This causes a shift in the non-amyloidogenic pathway and reduces the levels of Aβ produced. (B) Aβ can self-aggregate to form oligomers and eventually amyloid plaques. Some phytosterols are able to inhibit the formation of amyloid plaques by binding to Aβ, inhibiting aggregation, and thereby promoting the formation of nontoxic oligomers. Toxic Aβ monomers and oligomers have been shown to induce microglial activation and proliferation. Activated microglia secrete pro-inflammatory cytokines such as IL-1β and IL-6. (C) Some phytosterols have been shown to reduce the levels of these cytokines. Some phytosterols reduce oxidative stress by increasing the levels of antioxidant enzymes and reducing lipid peroxidation. (D) Acetylcholine (ACh), a neurotransmitter essential for processing memory and learning, is decreased in both concentration and function in AD. Decreased levels of ACh can be restored by anticholinesterase activity of various phytosterols. (E) ROS irreversibly oxidize DNA and are important mediators of Aβ-induced neuronal cell death in the development of AD. Abbreviations: APP: Amyloid Precursor Protein; AChE: Acetyl Cholinesterase Enzyme; BACE 1: Beta-Secretase 1; BChE: Butyl Cholinesterase Enzyme; Bcl-2: B-Cell Lymphoma 2; CAT: Catalase; GPX: Glutathione Peroxidase; HO 1: Heme Oxygenase; ROS: Reactive Oxygen Species; SOD: Superoxide Dismutase.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sharma, N.; Tan, M.A.; An, S.S.A. Phytosterols: Potential Metabolic Modulators in Neurodegenerative Diseases. Int. J. Mol. Sci. 2021, 22, 12255. https://doi.org/10.3390/ijms222212255
AMA Style
Sharma N, Tan MA, An SSA. Phytosterols: Potential Metabolic Modulators in Neurodegenerative Diseases. International Journal of Molecular Sciences. 2021; 22(22):12255. https://doi.org/10.3390/ijms222212255
Chicago/Turabian StyleSharma, Niti, Mario A. Tan, and Seong Soo A. An. 2021. "Phytosterols: Potential Metabolic Modulators in Neurodegenerative Diseases" International Journal of Molecular Sciences 22, no. 22: 12255. https://doi.org/10.3390/ijms222212255
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.