
Identification and Functional Analysis of a Pseudo-Cysteine Protease from the Midgut Transcriptome of Sphenophorus levis

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Sphenophorus levis Midgut Transcriptome and Identification of Putative Pseudo-Cysteine Proteases
2.2. Identification of Putative Proteases
2.3. Relative Expression of Sl-CathL-CS in Different Parts of the Larvae
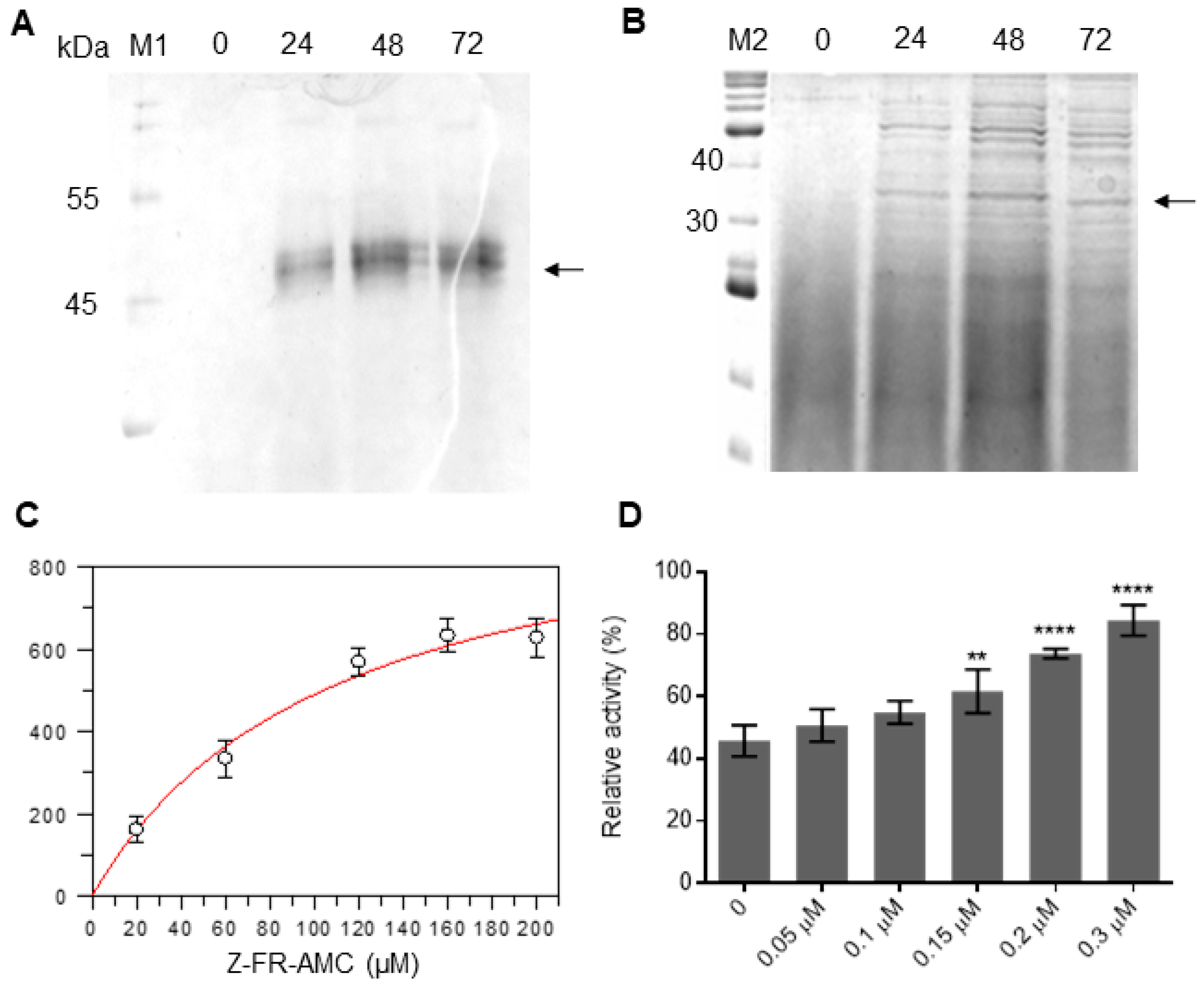
2.4. Heterologous Expression of Sl-CathL-CS and Sl-CathL-mutSC
2.5. Enzymatic Characterization of Sl-CathL-CS and Sl-CathL-mutSC
2.5.1. Sl-CathL-CS and Sl-CathL-mutSC Protease Activity
2.5.2. Competition Assay
2.6. Pull-Down
2.7. Static Protein Ligand Docking
3. Discussion
4. Materials and Methods
4.1. RNA Extraction
4.2. Library Preparation and Illumina Sequencing
4.3. Quality Control, Preprocessing and de Novo Assembly
4.4. Identification of Putative Pseudo-Cysteine Proteases
4.5. Relative Gene Expression of Sl-CathL-CS in Different Developmental Stages and Parts of the Larvae
4.6. Heterologous Expression of Sl-CathL-CS and Sl-CathLCS-mutSC
4.7. Enzymatic Characterization of Sl-CathL-CS and Sl-CathL-mutSC
4.7.1. Sl-CathL-CS and Sl-CathL-mutSC Protease Activity
4.7.2. Competition Assay
4.8. Pull-Down
4.9. Static Protein-Ligand Docking
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Degaspari, N.; Botelho, P.S.M.; Almeida, L.C.; Castilho, H.J. Biologia de Sphenophorus levis Vaurie, 1978 (col.: Curculionidae), em dieta artificial e no campo. Pesqui. Agropecu. Bras. 1987, 22, 553–558. [Google Scholar]
- Fonseca, F.P.; Soares-Costa, A.; Ribeiro, A.F.; Rosa, J.C.; Terra, W.R.; Henrique-Silva, F. Recombinant expression, localization and in vitro inhibition of midgut cysteine peptidase (Sl-CathL) from sugarcane weevil, Sphenophorus levis. Insect Biochem. Mol. Biol. 2012, 42, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Bown, D.P.; Wilkinson, H.S.; Jongsma, M.A.; Gatehouse, J.A. Characterisation of cysteine proteinases responsible for digestive proteolysis in guts of larval western corn rootworm (Diabrotica virgifera) by expression in the yeast Pichia pastoris. Insect Biochem. Mol. Biol. 2004, 34, 305–320. [Google Scholar] [CrossRef] [PubMed]
- Papisova, A.I.; Javadov, A.; Rudenskaya, Y.A.; Balandina, G.N.; Zhantiev, R.D.; Korsunovskaia, O.S.; Dunaevsky, Y.E.; Rudenskaya, G.N. Novel cathepsin L-like protease from dermestid beetle Dermestes frischii maggot. Biochimie 2011, 93, 141–148. [Google Scholar] [CrossRef]
- Soares-Costa, A.; Dias, A.B.; Dellamano, M.; de Paula, F.F.; Carmona, A.K.; Terra, W.R.; Henrique-Silva, F. Digestive physiology and characterization of digestive cathepsin L-like proteinase from the sugarcane weevil Sphenophorus levis. J. Insect Physiol. 2011, 57, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Vinokurov, K.S.; Elpidina, E.N.; Zhuzhikov, D.P.; Oppert, B.; Kodrik, D.; Sehnal, F. Digestive proteolysis organization in two closely related Tenebrionid beetles: Red flour beetle (Tribolium castaneum) and confused flour beetle (Tribolium confusum). Arch. Insect Biochem. Physiol. 2009, 70, 254–279. [Google Scholar] [CrossRef]
- Vinokurov, K.S.; Elpidina, E.N.; Oppert, B.; Prabhakar, S.; Zhuzhikov, D.P.; Dunaevsky, Y.E.; Belozersky, M.A. Diversity of digestive proteinases in Tenebrio molitor (Coleoptera: Tenebrionidae) larvae. Comp. Biochem. Physiol. Biochem. Mol. Biol. 2006, 145, 126–137. [Google Scholar] [CrossRef]
- Christeller, J.T.; Laing, W.A.; Markwick, N.P.; Burgess, E.P.J. Midgut protease activities in 12 phytophagous lepidopteran larvae: Dietary and protease inhibitor interactions. Insect Biochem. Mol. Biol. 1992, 22, 735–746. [Google Scholar] [CrossRef]
- Srinivasan, A.; Giri, A.P.; Gupta, V.S. Structural and functional diversities in lepidopteran serine proteases. Cell. Mol. Biol. Lett. 2006, 11, 132–154. [Google Scholar] [CrossRef]
- Pedezzi, R.; Fonseca, F.P.; Santos Junior, C.D.; Kishi, L.T.; Terra, W.R.; Henrique-Silva, F. A novel beta-fructofuranosidase in Coleoptera: Characterization of a beta-fructofuranosidase from the sugarcane weevil, Sphenophorus levis. Insect Biochem. Mol. Biol. 2014, 55, 31–38. [Google Scholar] [CrossRef]
- Habrylo, O.; Evangelista, D.E.; Castilho, P.V.; Pelloux, J.; Henrique-Silva, F. The pectinases from Sphenophorus levis: Potential for biotechnological applications. Int. J. Biol. Macromol. 2018, 112, 499–508. [Google Scholar] [CrossRef]
- Evangelista, D.E.; de Paula, F.F.; Rodrigues, A.; Henrique-Silva, F. Pectinases from Sphenophorus levis Vaurie, 1978 (Coleoptera: Curculionidae): Putative accessory digestive enzymes. J. Insect Sci. 2015, 15, 168. [Google Scholar] [CrossRef]
- Shibao, P.Y.T.; Santos-Júnior, C.D.; Santiago, A.C.; Mohan, C.; Miguel, M.C.; Toyama, D.; Vieira, M.A.S.; Narayanan, S.; Figueira, A.; Carmona, A.K.; et al. Sugarcane cystatins: From discovery to biotechnological applications. Int. J. Biol. Macromol. 2021, 167, 676–686. [Google Scholar] [CrossRef]
- Endara, M.J.; Coley, P.D.; Ghabash, G.; Nicholls, J.A.; Dexter, K.G.; Donoso, D.A.; Stone, G.N.; Pennington, R.T.; Kursar, T.A. Coevolutionary arms race versus host defense chase in a tropical herbivore-plant system. Proc. Natl. Acad. Sci. USA 2017, 114, E7499–E7505. [Google Scholar] [CrossRef] [Green Version]
- Eyers, P.A.; Murphy, J.M. The evolving world of pseudoenzymes: Proteins, prejudice and zombies. BMC Biol. 2016, 14, 98. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.M.; Farhan, H.; Eyers, P.A. Bio-Zombie: The rise of pseudoenzymes in biology. Biochem. Soc. Trans. 2017, 45, 537–544. [Google Scholar] [CrossRef]
- Ribeiro, A.J.M.; Das, S.; Dawson, N.; Zaru, R.; Orchard, S.; Thornton, J.M.; Orengo, C.; Zeqiraj, E.; Murphy, J.M.; Eyers, P.A. Emerging concepts in pseudoenzyme classification, evolution, and signaling. Sci. Signal. 2019, 12. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Parizi, L.F.; Torquato, R.J.S.; Vaz Junior, I.S.; Tanaka, A.S. Novel pseudo-aspartic peptidase from the midgut of the tick Rhipicephalus microplus. Sci. Rep. 2019, 9, 435. [Google Scholar] [CrossRef] [Green Version]
- Soares-Costa, A.; Beltramini, L.M.; Thiemann, O.H.; Henrique-Silva, F. A sugarcane cystatin: Recombinant expression, purification, and antifungal activity. Biochem. Biophys. Res. Commun. 2002, 296, 1194–1199. [Google Scholar] [CrossRef]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [Green Version]
- Hirata, I.Y.; Sedenho Cezari, M.H.; Nakaie, C.R.; Boschcov, P.; Ito, A.S.; Juliano, M.A.; Juliano, L. Internally quenched fluorogenic protease substrates: Solid-phase synthesis and fluorescence spectroscopy of peptides containing ortho-aminobenzoyl/dinitrophenyl groups as donor-acceptor pairs. Lett. Pept. Sci. 1995, 1, 299–308. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant. Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Zhu-Salzman, K.; Koiwa, H.; Salzman, R.A.; Shade, R.E.; Ahn, J.E. Cowpea bruchid Callosobruchus maculatus uses a three-component strategy to overcome a plant defensive cysteine protease inhibitor. Insect Mol. Biol. 2003, 12, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Benchabane, M.; Schluter, U.; Vorster, J.; Goulet, M.C.; Michaud, D. Plant cystatins. Biochimie 2010, 92, 1657–1666. [Google Scholar] [CrossRef]
- Martinez, M.; Santamaria, M.E.; Diaz-Mendoza, M.; Arnaiz, A.; Carrillo, L.; Ortego, F.; Diaz, I. Phytocystatins: Defense Proteins against Phytophagous Insects and Acari. Int. J. Mol. Sci. 2016, 17, 1747. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, L.; Martinez, M.; Ramessar, K.; Cambra, I.; Castanera, P.; Ortego, F.; Diaz, I. Expression of a barley cystatin gene in maize enhances resistance against phytophagous mites by altering their cysteine-proteases. Plant. Cell. Rep. 2011, 30, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Fabrick, J.; Behnke, C.; Czapla, T.; Bala, K.; Rao, A.G.; Kramer, K.J.; Reeck, G.R. Effects of a potato cysteine proteinase inhibitor on midgut proteolytic enzyme activity and growth of the southern corn rootworm, Diabrotica undecimpunctata howardi (Coleoptera: Chrysomelidae). Insect Biochem. Mol. Biol. 2002, 32, 405–415. [Google Scholar] [CrossRef]
- Valdés-Rodríguez, S.; Galván-Ramírez, J.P.; Guerrero-Rangel, A.; Cedro-Tanda, A. Multifunctional amaranth cystatin inhibits endogenous and digestive insect cysteine endopeptidases: A potential tool to prevent proteolysis and for the control of insect pests. Biotechnol. Appl. Biochem. 2015, 62, 634–641. [Google Scholar] [CrossRef]
- De Oliveira, C.F.R.; de Paula Souza, T.; Parra, J.R.P.; Marangoni, S.; de Castro Silva-Filho, M.; Macedo, M.L.R. Insensitive trypsins are differentially transcribed during Spodoptera frugiperda adaptation against plant protease inhibitors. Comp. Biochem. Physiol. Biochem. Mol. Biol. 2013, 165, 19–25. [Google Scholar] [CrossRef]
- Lopes, A.R.; Juliano, M.A.; Juliano, L.; Terra, W.R. Coevolution of insect trypsins and inhibitors. Arch. Insect Biochem. Physiol. 2004, 55, 140–152. [Google Scholar] [CrossRef]
- Brunelle, F.; Cloutier, C.; Michaud, D. Colorado potato beetles compensate for tomato cathepsin D inhibitor expressed in transgenic potato. Arch. Insect Biochem. Physiol. 2004, 55, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Cloutier, C.; Jean, C.; Fournier, M.; Yelle, S.; Michaud, D. Adult Colorado potato beetles, Leptinotarsa decemlineata compensate for nutritional stress on oryzacystatin I-transgenic potato plants by hypertrophic behavior and over-production of insensitive proteases. Arch. Insect Biochem. Physiol. 2000, 44, 69–81. [Google Scholar] [CrossRef]
- Vorster, J.; Rasoolizadeh, A.; Goulet, M.C.; Cloutier, C.; Sainsbury, F.; Michaud, D. Positive selection of digestive Cys proteases in herbivorous Coleoptera. Insect Biochem. Mol. Biol. 2015, 65, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruden, K.; Popovic, T.; Cimerman, N.; Krizaj, I.; Strukelj, B. Diverse enzymatic specificities of digestive proteases, ‘intestains’, enable Colorado potato beetle larvae to counteract the potato defence mechanism. Biol. Chem. 2003, 384, 305–310. [Google Scholar] [CrossRef]
- Souza, T.P.; Dias, R.O.; Castelhano, E.C.; Brandão, M.M.; Moura, D.S.; Silva-Filho, M.C. Comparative analysis of expression profiling of the trypsin and chymotrypsin genes from Lepidoptera species with different levels of sensitivity to soybean peptidase inhibitors. Comp. Biochem. Physiol. Biochem. Mol. Biol. 2016, 196–197, 67–73. [Google Scholar] [CrossRef]
- Zhu-Salzman, K.; Zeng, R. Insect response to plant defensive protease inhibitors. Annu. Rev. Entomol. 2015, 60, 233–252. [Google Scholar] [CrossRef]
- Barrett, A.J.; Kirschke, H. Cathepsin B, Cathepsin H, and cathepsin L. Methods Enzymol. 1981, 80, 535–561. [Google Scholar] [CrossRef]
- Barrett, A.J.; Kembhavi, A.A.; Brown, M.A.; Kirschke, H.; Knight, C.G.; Tamai, M.; Hanada, K. L-trans-Epoxysuccinyl-leucylamido(4-guanidino)butane (E-64) and its analogues as inhibitors of cysteine proteinases including cathepsins B, H and L. Biochem. J. 1982, 201, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Sekar, V.; Hageman, J.H. Specificity of the serine protease inhibitor, phenylmethylsulfonyl fluoride. Biochem. Biophys. Res. Commun. 1979, 89, 474–478. [Google Scholar] [CrossRef]
- Schneider, V.K.; Soares-Costa, A.; Chakravarthi, M.; Ribeiro, C.; Chabregas, S.M.; Falco, M.C.; Henrique-Silva, F. Transgenic sugarcane overexpressing CaneCPI-1 negatively affects the growth and development of the sugarcane weevil Sphenophorus levis. Plant. Cell Rep. 2017, 36, 193–201. [Google Scholar] [CrossRef]
- Yokoyama, T.; Silversides, D.W.; Waymire, K.G.; Kwon, B.S.; Takeuchi, T.; Overbeek, P.A. Conserved cysteine to serine mutation in tyrosinase is responsible for the classical albino mutation in laboratory mice. Nucleic Acids Res. 1990, 18, 7293–7298. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Quiocho, F. A nonconservative serine to cysteine mutation in the sulfate-binding protein, a transport receptor. Science 1991, 251, 1479–1481. [Google Scholar] [CrossRef]
- Chou, W.Y.; Matthews, K.S. Serine to cysteine mutations in trp repressor protein alter tryptophan and operator binding. J. Biol. Chem. 1989, 264, 18314–18319. [Google Scholar] [CrossRef]
- Santos, J.; Risso, V.A.; Sica, M.P.; Ermácora, M.R. Effects of Serine-to-Cysteine Mutations on β-Lactamase Folding. Biophys. J. 2007, 93, 1707–1718. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data [Online]. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 31 August 2021).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Seppey, M.; Manni, M.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness. Methods Mol. Biol. 2019, 1962, 227–245. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Morrison, J.F. The slow-binding and slow, tight-binding inhibition of enzyme-catalysed reactions. Trends Biochem. Sci. 1982, 7, 102–105. [Google Scholar] [CrossRef]
- Leatherbarrow, R. GraFit Version 5; Erithacus Software Ltd.: Horley, UK, 2001. [Google Scholar]
- Eswar, N.; Webb, B.; Marti-Renom, M.A.; Madhusudhan, M.S.; Eramian, D.; Shen, M.-Y.; Pieper, U.; Sali, A. Comparative protein structure modeling using Modeller. Curr. Protoc. Bioinform. 2006, 5, 56. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Lasker, K.; Schneidman-Duhovny, D.; Webb, B.; Huang, C.C.; Pettersen, E.F.; Goddard, T.D.; Meng, E.C.; Sali, A.; Ferrin, T.E. UCSF Chimera, MODELLER, and IMP: An integrated modeling system. J. Struct. Biol. 2012, 179, 269–278. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of raw reads | 70,720,844 |
| Number of readings after filtering | 146,145 |
| Average length of contigs (bp) | 1131.23 |
| N50 length (bp) | 2382 |
| GC Percent | 37.20 |
| Primer | Sequence (5′–3′) | Purpose | Amplicon (bp) |
|---|---|---|---|
| Sl-CathL-CS_fw: | CCGAATTCAGTTCGGAGCTGAACATGG | ORF cloning | 984 |
| Sl-CathL-CS_rv | ATTCTTATGCGGCCGCATCGATTTCGACGTAGGCAGC | ||
| Sl-CathL-mutSCforw | ACCAAGGAGAATGGGATACATGTTGGGCTTTCTCCACTATTGC | Site-directed mutagenesis | |
| Sl-CathL-mutSC_rev | GCAATAGTGGAGAAAGCCCAACATGTATCCCATTCTCCTTGGT | ||
| Sl_GADPH_qF | CAACTGGCGTTTTTACCACA | Real Time PCR | 104 |
| Sl_GADPH_qR | AACATACATTGGGGCGTCA | ||
| Sl_CathL-CS_qF: | ATACGACTGGAGGGAGCAGA | Real Time PCR | 108 |
| Sl_CathL-CS_qR | ATGGCGTAGGCACTTTCAAC | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shibao, P.Y.T.; Ferro, M.; de Paula, F.F.P.; Lima, B.S.; Henrique-Silva, F. Identification and Functional Analysis of a Pseudo-Cysteine Protease from the Midgut Transcriptome of Sphenophorus levis. Int. J. Mol. Sci. 2021, 22, 11476. https://doi.org/10.3390/ijms222111476
Shibao PYT, Ferro M, de Paula FFP, Lima BS, Henrique-Silva F. Identification and Functional Analysis of a Pseudo-Cysteine Protease from the Midgut Transcriptome of Sphenophorus levis. International Journal of Molecular Sciences. 2021; 22(21):11476. https://doi.org/10.3390/ijms222111476
Chicago/Turabian StyleShibao, Priscila Yumi Tanaka, Milene Ferro, Fernando Fonseca Pereira de Paula, Bruno Salata Lima, and Flávio Henrique-Silva. 2021. "Identification and Functional Analysis of a Pseudo-Cysteine Protease from the Midgut Transcriptome of Sphenophorus levis" International Journal of Molecular Sciences 22, no. 21: 11476. https://doi.org/10.3390/ijms222111476