
MAP/Microtubule Affinity Regulating Kinase 4 Inhibitory Potential of Irisin: A New Therapeutic Strategy to Combat Cancer and Alzheimer’s Disease



 ,
,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Expression and Purification of Irisin and MARK4
2.3. Fluorescence Measurements
2.4. Isothermal Titration Calorimetry
2.5. Kinase Assay
2.6. Protein Structure Modeling and Preparation
2.7. Protein–Protein Docking
2.8. Molecular Dynamics Simulations
2.9. Post-Dynamic Trajectories Analysis
2.10. Dynamic of the Cross-Correlation Matrix
2.11. Binding Energy Calculation
3. Result and Discussion
3.1. Fluorescence-Based Binding Analysis
3.2. Isothermal Titration Calorimetry
3.3. Kinase Assay
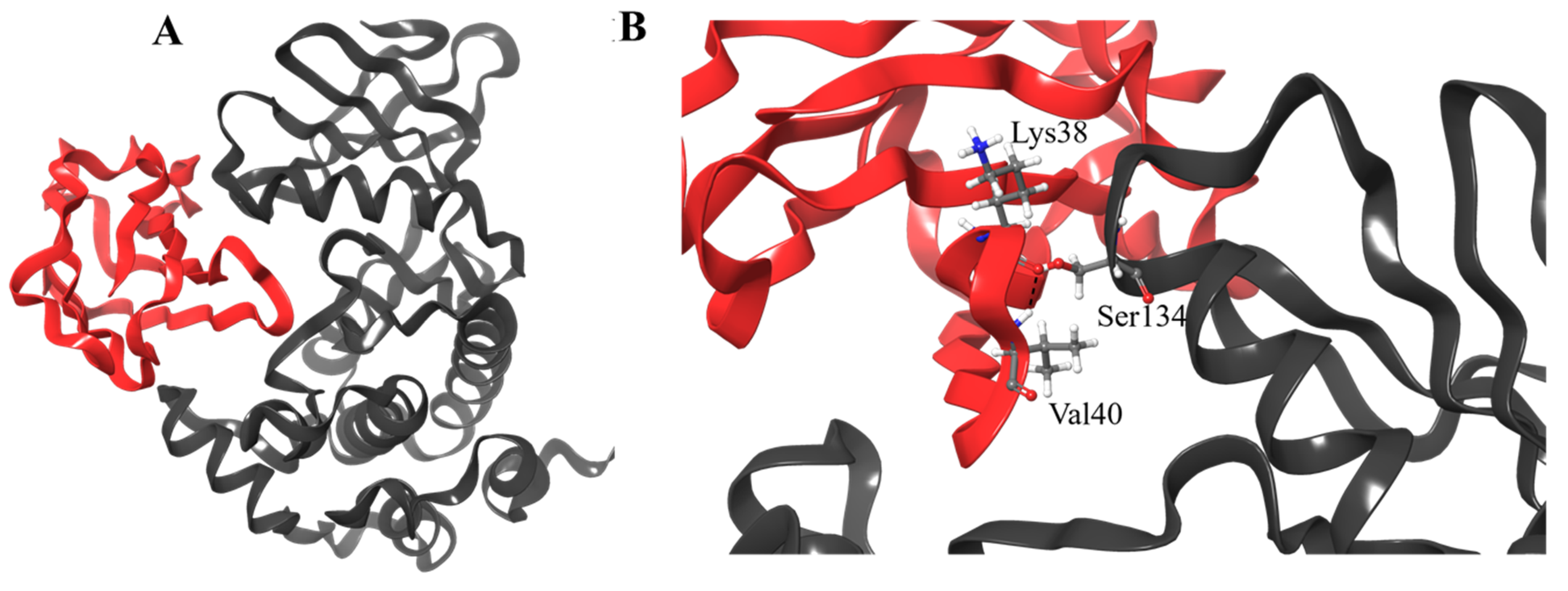
3.4. Molecular Docking
3.5. Post-Dynamics Trajectories Analysis
3.6. Hydrogen Bond Analysis
3.7. Distance Correlation Matrix
3.8. Secondary Structure Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Jedrychowski, M.P.; Wrann, C.D.; Paulo, J.A.; Gerber, K.K.; Szpyt, J.; Robinson, M.M.; Nair, K.S.; Gygi, S.P.; Spiegelman, B.M. Detection and quantitation of circulating human irisin by tandem mass spectrometry. Cell Metab. 2015, 22, 734–740. [Google Scholar] [CrossRef] [Green Version]
- Waseem, R.; Shamsi, A.; Mohammad, T.; Alhumaydhi, F.A.; Kazim, S.N.; Hassan, M.I.; Ahmad, F.; Islam, A. Multispectroscopic and Molecular Docking Insight into Elucidating the Interaction of Irisin with Rivastigmine Tartrate: A Combinational Therapy Approach to Fight Alzheimer’s Disease. ACS Omega 2021, 6, 7910–7921. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Pan, J. Irisin ameliorates depressive-like behaviors in rats by regulating energy metabolism. Biochem. Biophys. Res. Commun. 2016, 474, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.-Y.; Shi, C.-X.; Gao, R.; Sun, H.-J.; Xiong, X.-Q.; Ding, L.; Chen, Q.; Li, Y.-H.; Wang, J.-J.; Kang, Y.M.; et al. Irisin inhibits hepatic gluconeogenesis and increases glycogen synthesis via the PI3K/Akt pathway in type 2 diabetic mice and hepatocytes. Clin. Sci. 2015, 129, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-Q.; Huang, Y.-Y.; Gusdon, A.M.; Qu, S. Irisin: A new molecular marker and target in metabolic disorder. Lipids Health Dis. 2015, 14, 2. [Google Scholar] [CrossRef] [Green Version]
- Qiao, X.; Nie, Y.; Ma, Y.; Chen, Y.; Cheng, R.; Yin, W.; Hu, Y.; Xu, W.; Xu, L. Irisin promotes osteoblast proliferation and differentiation via activating the MAP kinase signaling pathways. Sci. Rep. 2016, 6, 18732. [Google Scholar] [CrossRef] [Green Version]
- Novelle, M.G.; Contreras, C.; Romero-Picó, A.; López, M.; Diéguez, C. Irisin, two years later. Int. J. Endocrinol. 2013, 2013, 746281. [Google Scholar] [CrossRef] [Green Version]
- Kim, O.Y.; Song, J. The role of irisin in Alzheimer’s disease. J. Clin. Med. 2018, 7, 407. [Google Scholar] [CrossRef] [Green Version]
- Lourenco, M.V.; Frozza, R.L.; de Freitas, G.B.; Zhang, H.; Kincheski, G.C.; Ribeiro, F.C.; Gonçalves, R.A.; Clarke, J.R.; Beckman, D.; Staniszewski, A.; et al. Exercise-linked FNDC5/irisin rescues synaptic plasticity and memory defects in Alzheimer’s models. Nat. Med. 2019, 25, 165–175. [Google Scholar] [CrossRef]
- Lund, H.; Gustafsson, E.; Svensson, A.; Nilsson, M.; Berg, M.; Sunnemark, D.; von Euler, G. MARK4 and MARK3 associate with early tau phosphorylation in Alzheimer’s disease granulovacuolar degeneration bodies. Acta Neuropathol. Commun. 2014, 2, 22. [Google Scholar] [CrossRef] [Green Version]
- Mandelkow, E.-M.; Thies, E.; Trinczek, B.; Biernat, J.; Mandelkow, E. MARK/PAR1 kinase is a regulator of microtubule-dependent transport in axons. J. Cell Biol. 2004, 167, 99–110. [Google Scholar] [CrossRef]
- Sun, W.; Lee, S.; Huang, X.; Liu, S.; Inayathullah, M.; Kim, K.-M.; Tang, H.; Ashford, J.W.; Rajadas, J. Attenuation of synaptic toxicity and MARK4/PAR1-mediated Tau phosphorylation by methylene blue for Alzheimer’s disease treatment. Sci. Rep. 2016, 6, 34784. [Google Scholar] [CrossRef]
- Jenardhanan, P.; Mannu, J.; Mathur, P.P. The structural analysis of MARK4 and the exploration of specific inhibitors for the MARK family: A computational approach to obstruct the role of MARK4 in prostate cancer progression. Mol. Biosyst. 2014, 10, 1845–1868. [Google Scholar] [CrossRef] [PubMed]
- Naz, F.; Khan, F.I.; Mohammad, T.; Khan, P.; Manzoor, S.; Hasan, G.M.; Lobb, K.A.; Luqman, S.; Islam, A.; Ahmad, F.; et al. Investigation of molecular mechanism of recognition between citral and MARK4: A newer therapeutic approach to attenuate cancer cell progression. Int. J. Biol. Macromol. 2018, 107, 2580–2589. [Google Scholar] [CrossRef] [PubMed]
- Naz, F.; Anjum, F.; Islam, A.; Ahmad, F.; Hassan, M.I. Microtubule affinity-regulating kinase 4: Structure, function, and regulation. Cell Biochem. Biophys. 2013, 67, 485–499. [Google Scholar] [CrossRef]
- Naz, F.; Shahbaaz, M.; Bisetty, K.; Islam, A.; Ahmad, F.; Hassan, M.I. Designing New Kinase Inhibitor Derivatives as Therapeutics Against Common Complex Diseases: Structural Basis of Microtubule Affinity-Regulating Kinase 4 (MARK4) Inhibition. OMICS 2015, 19, 700–711. [Google Scholar] [CrossRef]
- Tiwari, M.K.; Coghi, P.; Agrawal, P.; Yadav, D.K.; Yang, L.J.; Congling, Q.; Sahal, D.; Wong, V.K.W.; Chaudhary, S. Novel halogenated arylvinyl-1, 2, 4 trioxanes as potent antiplasmodial as well as anticancer agents: Synthesis, bioevaluation, structure-activity relationship and in-silico studies. Eur. J. Med. Chem. 2021, 224, 113685. [Google Scholar] [CrossRef] [PubMed]
- Kalani, K.; Yadav, D.K.; Alam, S.; Khan, F.; Kashyap, M.P.; Srivastava, S.K.; Pant, A.B. In-silico Studies and Wet-Lab Validation of Camptothecin Derivatives for Anti-Cancer Activity Against Liver (HepG2) and Lung (A549) Cancer Cell Lines. Curr. Top. Med. Chem. 2021, 21, 1–12. [Google Scholar] [CrossRef]
- Yadav, D.K. Recent Advances on Small Molecule Medicinal Chemistry to Treat Human Diseases-Part II. Curr. Top. Med. Chem. 2021, 21, 849. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Park, S.; Kim, S.Y.; Um, S.H.; Moon, E.Y. Curcumin hampers the antitumor effect of vinblastine via the inhibition of microtubule dynamics and mitochondrial membrane potential in HeLa cervical cancer cells. Phytomedicine 2016, 23, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Gadhe, C.G.; Kim, M.H. Insights into the binding modes of CC chemokine receptor 4 (CCR4) inhibitors: A combined approach involving homology modelling, docking, and molecular dynamics simulation studies. Mol. Biosyst. 2015, 11, 618–634. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yi, S.H.; Lee, J.; Kim, Y.S. Effects of clozapine on heart rate dynamics and their relationship with therapeutic response in treatment-resistant schizophrenia. J. Clin. Psychopharmacol. 2013, 33, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Ann, J.H.; Lee, J.; Kim, M.H.; Han, A.Y. Altered heart rate dynamics associated with antipsychotic-induced subjective restlessness in patients with schizophrenia. Neuropsychiatr. Dis. Treat. 2013, 9, 989–994. [Google Scholar] [CrossRef] [Green Version]
- Yadav, D.K.; Kumar, S.; Teli, M.K.; Kim, M.H. Ligand-based pharmacophore modeling and docking studies on vitamin D receptor inhibitors. J. Cell. Biochem. 2020, 121, 3570–3583. [Google Scholar] [CrossRef]
- Shyamlal, B.R.K.; Mathur, M.; Yadav, D.K.; Chaudhary, S. Microwave-assisted modified synthesis of C8-analogues of naturally occurring methylxanthines: Synthesis, biological evaluation and their practical applications. Fitoterapia 2020, 143, 104533. [Google Scholar] [CrossRef]
- Dileep, K.; Ashok, S.; Remya, C.; Dharmendra, K.; Pérez-Sánchez, H.; Sadasivan, C. Indole fragments for the design of lead molecules against pancreatitis. J. Biomol. Struct. Dyn. 2020, 38, 263–267. [Google Scholar] [CrossRef]
- Yadav, D.K.; Kumar, S.; Choi, E.-H.; Chaudhary, S.; Kim, M.-H. Computational Modeling on Aquaporin-3 as Skin Cancer Target: A Virtual Screening and Molecular Dynamic Simulation Study. Front. Chem. 2020, 8, 250. [Google Scholar] [CrossRef] [Green Version]
- Yadav, D.K.; Saloni, S.K.; Kim, M.-H. Proteomics and Molecular Medicine Bioinformatics. J. Proteom. 2017, 10. [Google Scholar] [CrossRef]
- Khan, P.; Rahman, S.; Queen, A.; Manzoor, S.; Naz, F.; Hasan, G.M.; Luqman, S.; Kim, J.; Islam, A.; Ahmad, F.; et al. Elucidation of Dietary Polyphenolics as Potential Inhibitor of Microtubule Affinity Regulating Kinase 4: In silico and In vitro Studies. Sci. Rep. 2017, 7, 9470. [Google Scholar] [CrossRef]
- Dimitrov, D.S. Therapeutic Proteins; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–26. [Google Scholar]
- Cheon, J.H.; Kim, K.S.; Yadav, D.K.; Kim, M.; Kim, H.S.; Yoon, S. The JAK2 inhibitors CEP-33779 and NVP-BSK805 have high P-gp inhibitory activity and sensitize drug-resistant cancer cells to vincristine. Biochem. Biophys. Res. Commun. 2017, 490, 1176–1182. [Google Scholar] [CrossRef]
- Venkanna, A.; Kwon, O.W.; Afzal, S.; Jang, C.; Cho, K.H.; Yadav, D.K.; Kim, K.; Park, H.-g.; Chun, K.-H.; Kim, S.Y. Pharmacological use of a novel scaffold, anomeric N, N-diarylamino tetrahydropyran: Molecular similarity search, chemocentric target profiling, and experimental evidence. Sci. Rep. 2017, 7, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Yadav, D.K.; Kumar, S.; Misra, S.; Yadav, L.; Teli, M.; Sharma, P.; Chaudhary, S.; Kumar, N.; Choi, E.H.; Kim, H.S. Molecular insights into the interaction of RONS and Thieno [3, 2-c] pyran analogs with SIRT6/COX-2: A molecular dynamics study. Sci. Rep. 2018, 8, 1–16. [Google Scholar] [CrossRef]
- Dwivedi, G.R.; Maurya, A.; Yadav, D.K.; Khan, F.; Darokar, M.P.; Srivastava, S.K. Drug resistance reversal potential of ursolic acid derivatives against nalidixic acid-and multidrug-resistant Escherichia coli. Chem. Biol. Drug Des. 2015, 86, 272–283. [Google Scholar] [CrossRef]
- Jaiswal, P.K.; Sharma, V.; Kumar, S.; Mathur, M.; Swami, A.K.; Yadav, D.K.; Chaudhary, S. Non-peptide-based new class of platelet aggregation inhibitors: Design, synthesis, bioevaluation, SAR, and in silico studies. Archiv. Der. Pharmazie 2018, 351, 1700349. [Google Scholar] [CrossRef]
- Yadav, D.K.; Khan, F.; Negi, A.S. QSAR and Docking studies of Gallic acid derivatives for anticancer and Immunomodulatory activity. Nat. Chem. 2011, 1. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.B.; Ah Lee, J.; Soobin, J.; Lim Lee, H. Herbal medicine (Suoquan) for treating nocturnal enuresis: A protocol for a systematic review of randomized controlled trials. Medicine (Baltimore) 2018, 97, e0391. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Cho, S.G.; Choi, Y.K.; Choi, Y.J.; Lee, G.R.; Jeon, C.Y.; Ko, S.G. Herbal prescription, Danggui-Sayuk-Ga-Osuyu-Senggang-Tang, inhibits TNF-alpha-induced epithelial-mesenchymal transition in HCT116 colorectal cancer cells. Int. J. Mol. Med. 2018, 41, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.L.; Lee, Y.B.; Choi, J.Y.; Lee, J.A. Herbal medicine for idiopathic central precocious puberty: A protocol for a systematic review of controlled trials. Medicine (Baltimore) 2018, 97, e0267. [Google Scholar] [CrossRef]
- Park, S.J.; Kim, M.O.; Kim, J.H.; Jeong, S.; Kim, M.H.; Yang, S.J.; Lee, J.; Lee, H.J. Antioxidant Activities of Functional Beverage Concentrates Containing Herbal Medicine Extracts. Prev. Nutr. Food. Sci. 2017, 22, 16–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ku, S.K.; Kim, H.; Kim, J.W.; Kang, K.S.; Lee, H.J. Ameliorating effects of herbal formula hemomine on experimental subacute hemorrhagic anemia in rats. J. Ethnopharmacol. 2017, 198, 205–213. [Google Scholar] [CrossRef]
- Hussain, A.; Yadav, M.K.; Bose, S.; Wang, J.H.; Lim, D.; Song, Y.K.; Ko, S.G.; Kim, H. Daesiho-Tang Is an Effective Herbal Formulation in Attenuation of Obesity in Mice through Alteration of Gene Expression and Modulation of Intestinal Microbiota. PLoS ONE 2016, 11, e0165483. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, M.P.; Singh, A.K.; Yadav, D.K.; Siddiqui, M.A.; Srivastava, R.K.; Chaturvedi, V.; Rai, N. 4-Hydroxy-trans-2-nonenal (4-HNE) induces neuronal SH-SY5Y cell death via hampering ATP binding at kinase domain of Akt1. Arch. Toxicol. 2015, 89, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Naz, F.; Asad, M.; Malhotra, P.; Islam, A.; Ahmad, F.; Hassan, M.I. Cloning, expression, purification and refolding of microtubule affinity-regulating kinase 4 expressed in Escherichia coli. Appl. Biochem. Biotechnol. 2014, 172, 2838–2848. [Google Scholar] [CrossRef]
- Khan, P.; Idrees, D.; Moxley, M.A.; Corbett, J.A.; Ahmad, F.; von Figura, G.; Sly, W.S.; Waheed, A.; Hassan, M.I. Luminol-based chemiluminescent signals: Clinical and non-clinical application and future uses. Appl. Biochem. Biotechnol. 2014, 173, 333–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamsi, A.; Anwar, S.; Mohammad, T.; Alajmi, M.F.; Hussain, A.; Rehman, M.; Hasan, G.M.; Islam, A.; Hassan, M.I. MARK4 inhibited by AChE inhibitors, donepezil and Rivastigmine tartrate: Insights into Alzheimer’s disease therapy. Biomolecules 2020, 10, 789. [Google Scholar] [CrossRef] [PubMed]
- Anwar, S.; Mohammad, T.; Shamsi, A.; Queen, A.; Parveen, S.; Luqman, S.; Hasan, G.M.; Alamry, K.A.; Azum, N.; Asiri, A.M.; et al. Discovery of Hordenine as a potential inhibitor of pyruvate dehydrogenase kinase 3: Implication in lung Cancer therapy. Biomedicines 2020, 8, 119. [Google Scholar] [CrossRef] [PubMed]
- Anwar, S.; Shamsi, A.; Shahbaaz, M.; Queen, A.; Khan, P.; Hasan, G.M.; Islam, A.; Alajmi, M.F.; Hussain, A.; Ahmad, F.; et al. Rosmarinic acid exhibits anticancer effects via MARK4 inhibition. Sci. Rep. 2020, 10, 10300. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Naz, F.; Sami, N.; Islam, A.; Ahmad, F.; Hassan, M.I. Ubiquitin-associated domain of MARK4 provides stability at physiological pH. Int. J. Biol. Macromol. 2016, 93, 1147–1154. [Google Scholar] [CrossRef]
- Naz, F.; Sami, N.; Naqvi, A.T.; Islam, A.; Ahmad, F.; Imtaiyaz Hassan, M. Evaluation of human microtubule affinity-regulating kinase 4 inhibitors: Fluorescence binding studies, enzyme, and cell assays. J. Biomol. Struct. Dyn. 2017, 35, 3194–3203. [Google Scholar] [CrossRef]
- Sastry, G.M.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput.-Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- Harder, E.; Damm, W.; Maple, J.; Wu, C.; Reboul, M.; Xiang, J.Y.; Wang, L.; Lupyan, D.; Dahlgren, M.K.; Knight, J.L.; et al. OPLS3: A force field providing broad coverage of drug-like small molecules and proteins. J. Chem. Theory Comput. 2016, 12, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Jaiswal, P.K.; Saran, M.; Yadav, D.K.; Saloni; Mathur, M.; Swami, A.K.; Misra, S.; Kim, M.H.; Chaudhary, S. Discovery of C-3 Tethered 2-oxo-benzo[1,4]oxazines as Potent Antioxidants: Bio-Inspired Based Design, Synthesis, Biological Evaluation, Cytotoxic, and in Silico Molecular Docking Studies. Front. Chem. 2018, 6, 56. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Jaiswal, P.K.; Kumar, S.; Mathur, M.; Swami, A.K.; Yadav, D.K.; Chaudhary, S. Discovery of Aporphine Analogues as Potential Antiplatelet and Antioxidant Agents: Design, Synthesis, Structure-Activity Relationships, Biological Evaluations, and in silico Molecular Docking Studies. Chem. Med. Chem. 2018, 13, 1817–1832. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.H.; Hyun, J.S.; Choi, J.; Choi, K.E.; Jee, J.G.; Park, S.J. Structural ensemble-based docking simulation and biophysical studies discovered new inhibitors of Hsp90 N-terminal domain. Sci. Rep. 2018, 8, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozakov, D.; Brenke, R.; Comeau, S.R.; Vajda, S. PIPER: An FFT-based protein docking program with pairwise potentials. Proteins Struct. Funct. Bioinform. 2006, 65, 392–406. [Google Scholar] [CrossRef] [Green Version]
- Hospital, A.; Goñi, J.R.; Orozco, M.; Gelpí, J.L. Molecular dynamics simulations: Advances and applications. Adv. Appl. Bioinform. Chem. AABC 2015, 8, 37. [Google Scholar]
- Lee, T.-S.; Cerutti, D.S.; Mermelstein, D.; Lin, C.; LeGrand, S.; Giese, T.J.; Roitberg, A.; Case, D.A.; Walker, R.C.; York, D.M. GPU-accelerated molecular dynamics and free energy methods in Amber18: Performance enhancements and new features. J. Chem. Inf. Modeling 2018, 58, 2043–2050. [Google Scholar] [CrossRef]
- Perez, A.; MacCallum, J.L.; Brini, E.; Simmerling, C.; Dill, K.A. Grid-based backbone correction to the ff12SB protein force field for implicit-solvent simulations. J. Chem. Theory Comput. 2015, 11, 4770–4779. [Google Scholar] [CrossRef] [Green Version]
- Harrach, M.F.; Drossel, B. Structure and dynamics of TIP3P, TIP4P, and TIP5P water near smooth and atomistic walls of different hydroaffinity. J. Chem. Phys. 2014, 140, 174501. [Google Scholar] [CrossRef]
- Wang, J.; Wang, W.; Kollman, P.A.; Case, D.A. Automatic atom type and bond type perception in molecular mechanical calculations. J. Mol. Graph. Model. 2006, 25, 247–260. [Google Scholar] [CrossRef]
- Lin, Y.; Pan, D.; Li, J.; Zhang, L.; Shao, X. Application of Berendsen barostat in dissipative particle dynamics for nonequilibrium dynamic simulation. J. Chem. Phys. 2017, 146, 124108. [Google Scholar] [CrossRef]
- Gonnet, P. P-SHAKE: A quadratically convergent SHAKE in O (n2). J. Comput. Phys. 2007, 220, 740–750. [Google Scholar] [CrossRef]
- Gupta, P.; Khan, S.; Fakhar, Z.; Hussain, A.; Rehman, M.; AlAjmi, M.F.; Islam, A.; Ahmad, F.; Hassan, M.I. Identification of potential inhibitors of calcium/calmodulin-dependent protein kinase IV from bioactive phytoconstituents. Oxidative Med. Cell. Longev. 2020, 2020, 2094635. [Google Scholar] [CrossRef]
- Roe, D.R.; Cheatham, T.E., III. PTRAJ and CPPTRAJ: Software for processing and analysis of molecular dynamics trajectory data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef] [PubMed]
- Seifert, E. OriginPro 9.1: Scientific data analysis and graphing software-software review. J. Chem. Inf. Modeling 2014, 54, 1552. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Anwar, S.; Shamsi, A.; Kar, R.K.; Queen, A.; Islam, A.; Ahmad, F.; Hassan, M.I. Structural and biochemical investigation of MARK4 inhibitory potential of cholic acid: Towards therapeutic implications in neurodegenerative diseases. Int. J. Biol. Macromol. 2020, 161, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, A.; Ahmed, A.; Khan, M.S.; Al Shahwan, M.; Husain, F.M.; Bano, B. Understanding the binding between Rosmarinic acid and serum albumin: In vitro and in silico insight. J. Mol. Liq. 2020, 311, 113348. [Google Scholar] [CrossRef]
- Soares, S.; Mateus, N.; De Freitas, V. Interaction of different polyphenols with bovine serum albumin (BSA) and human salivary α-amylase (HSA) by fluorescence quenching. J. Agric. Food Chem. 2007, 55, 6726–6735. [Google Scholar] [CrossRef] [PubMed]
- Klajnert, B.; Stanisławska, L.; Bryszewska, M.; Pałecz, B. Interactions between PAMAM dendrimers and bovine serum albumin. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2003, 1648, 115–126. [Google Scholar] [CrossRef]
- Shamsi, A.; Al Shahwan, M.; Ahamad, S.; Hassan, M.I.; Ahmad, F.; Islam, A. Spectroscopic, calorimetric and molecular docking insight into the interaction of Alzheimer’s drug donepezil with human transferrin: Implications of Alzheimer’s drug. J. Biomol. Struct. Dyn. 2020, 38, 1094–1102. [Google Scholar] [CrossRef]
- Banerjee, T.; Kishore, N. 2, 2, 2-trifluoroethanol-induced molten globule state of concanavalin A and energetics of 8-anilinonaphthalene sulfonate binding: Calorimetric and spectroscopic investigation. J. Phys. Chem. B 2005, 109, 22655–22662. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.S.; Khan, P.; Inam, A.; Ahmad, K.; Yousuf, M.; Islam, A.; Alib, S.; Azam, A.; Husain, M.; Hassan, M.I. Discovery of 4-(2-(dimethylamino) ethoxy) benzohydrazide derivatives as prospective microtubule affinity regulating kinase 4 inhibitors. RSC Adv. 2020, 10, 20129–20137. [Google Scholar] [CrossRef]
- Rao, V.S.; Srinivas, K.; Sujini, G.; Kumar, G. Protein-protein interaction detection: Methods and analysis. Int. J. Proteom. 2014, 2014, 47648. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Bjij, I.; Soliman, M.E. Selective covalent inhibition of “Allosteric Cys121” distort the binding of PTP1B enzyme: A novel therapeutic approach for cancer treatment. Cell Biochem. Biophys. 2019, 77, 203–211. [Google Scholar] [CrossRef]
- Khan, S.; Bjij, I.; Betz, R.M.; Soliman, M.E. Reversible versus irreversible inhibition modes of ERK2: A comparative analysis for ERK2 protein kinase in cancer therapy. Future Med. Chem. 2018, 10, 1003–1015. [Google Scholar] [CrossRef]
- Bornot, A.; Etchebest, C.; De Brevern, A.G. Predicting protein flexibility through the prediction of local structures. Proteins Struct. Funct. Bioinform. 2011, 79, 839–852. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Fakhar, Z.; Hussain, A.; Ahmad, A.; Jairajpuri, D.S.; Alajmi, M.F.; Hassan, M.I. Structure-based identification of potential SARS-CoV-2 main protease inhibitors. J. Biomol. Struct. Dyn. 2020, 1–14. [Google Scholar] [CrossRef]
- Chen, H.; Panagiotopoulos, A.Z. Molecular modeling of surfactant micellization using solvent-accessible surface area. Langmuir 2019, 35, 2443–2450. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ka (Association Constant) M−1 | ∆H (Enthalpy Change) cal/mol | ∆S (cal/mol/deg) |
|---|---|---|
| Ka1 = 2.59 × 103 ± 2.10 × 104 | ∆H1 = −1.549 × 106 7617 ± 6.75 × 107 | ∆S1 = −5.18 × 103 |
| Ka2 = 1.19 × 106 ± 9.73 | ∆H2 = 1.898 × 105 ± 1.04 × 106 | ∆S2 = 664 |
| Protein–Protein Complex | α | β | 310-Helix | Turn | Bend | Other |
|---|---|---|---|---|---|---|
| Apo MARK4 | 28 | 25 | 4 | 13 | 7 | 21 |
| Irisin-MARK4 | 26 | 27 | 6 | 16 | 11 | 23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Waseem, R.; Anwar, S.; Khan, S.; Shamsi, A.; Hassan, M.I.; Anjum, F.; Shafie, A.; Islam, A.; Yadav, D.K. MAP/Microtubule Affinity Regulating Kinase 4 Inhibitory Potential of Irisin: A New Therapeutic Strategy to Combat Cancer and Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 10986. https://doi.org/10.3390/ijms222010986
Waseem R, Anwar S, Khan S, Shamsi A, Hassan MI, Anjum F, Shafie A, Islam A, Yadav DK. MAP/Microtubule Affinity Regulating Kinase 4 Inhibitory Potential of Irisin: A New Therapeutic Strategy to Combat Cancer and Alzheimer’s Disease. International Journal of Molecular Sciences. 2021; 22(20):10986. https://doi.org/10.3390/ijms222010986
Chicago/Turabian StyleWaseem, Rashid, Saleha Anwar, Shama Khan, Anas Shamsi, Md. Imtaiyaz Hassan, Farah Anjum, Alaa Shafie, Asimul Islam, and Dharmendra Kumar Yadav. 2021. "MAP/Microtubule Affinity Regulating Kinase 4 Inhibitory Potential of Irisin: A New Therapeutic Strategy to Combat Cancer and Alzheimer’s Disease" International Journal of Molecular Sciences 22, no. 20: 10986. https://doi.org/10.3390/ijms222010986