
BCL-2 Inhibitor ABT-737 Effectively Targets Leukemia-Initiating Cells with Differential Regulation of Relevant Genes Leading to Extended Survival in a NRAS/BCL-2 Mouse Model of High Risk-Myelodysplastic Syndrome

 , , , , , , , , , , add
Show full author list
, , , , , , , , , , add
Show full author list
Abastract
1. Introduction
2. Results
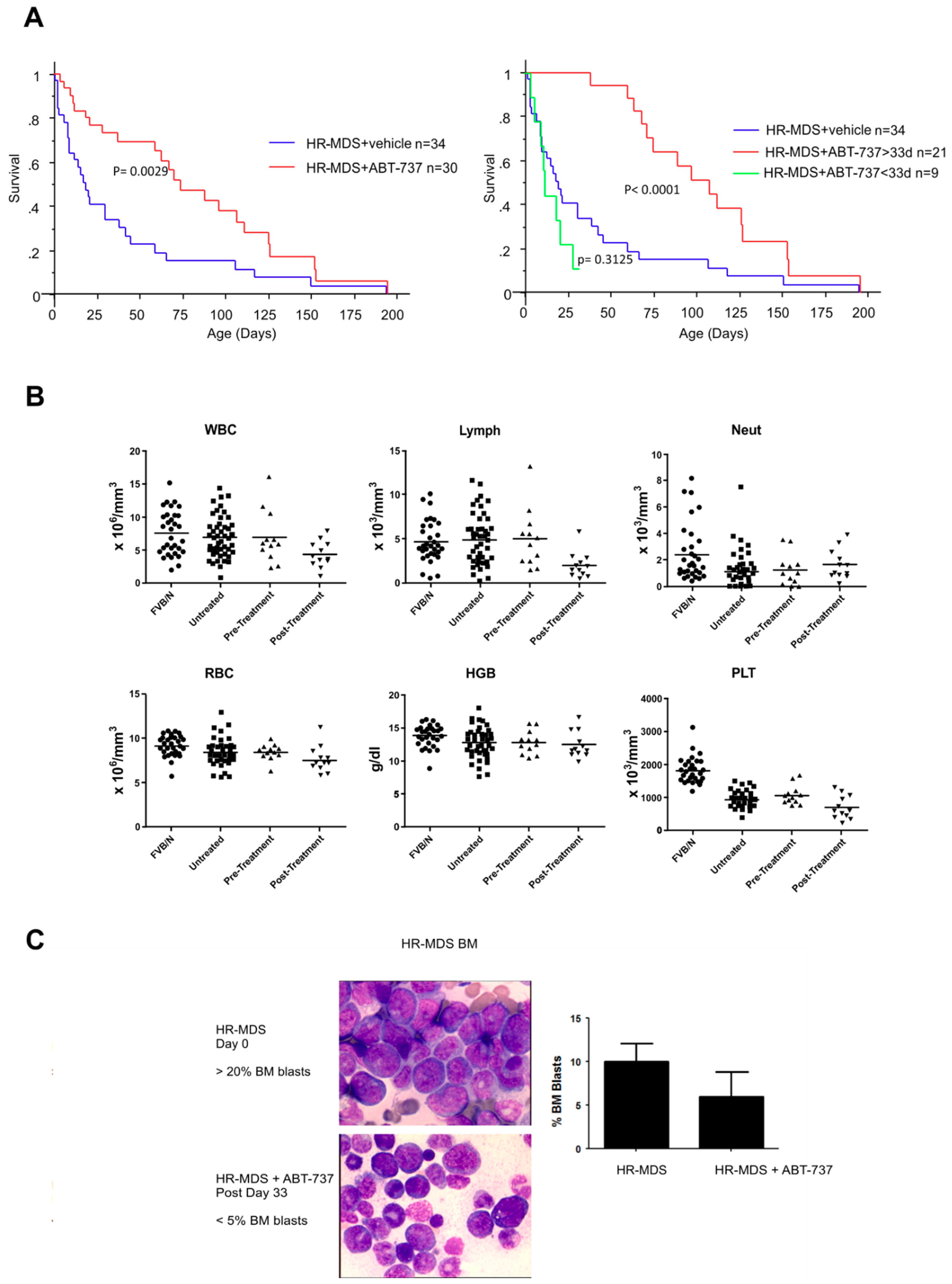
2.1. ABT-737 Treatment Prolongs Survival in HR-MDS Transgenic Mice
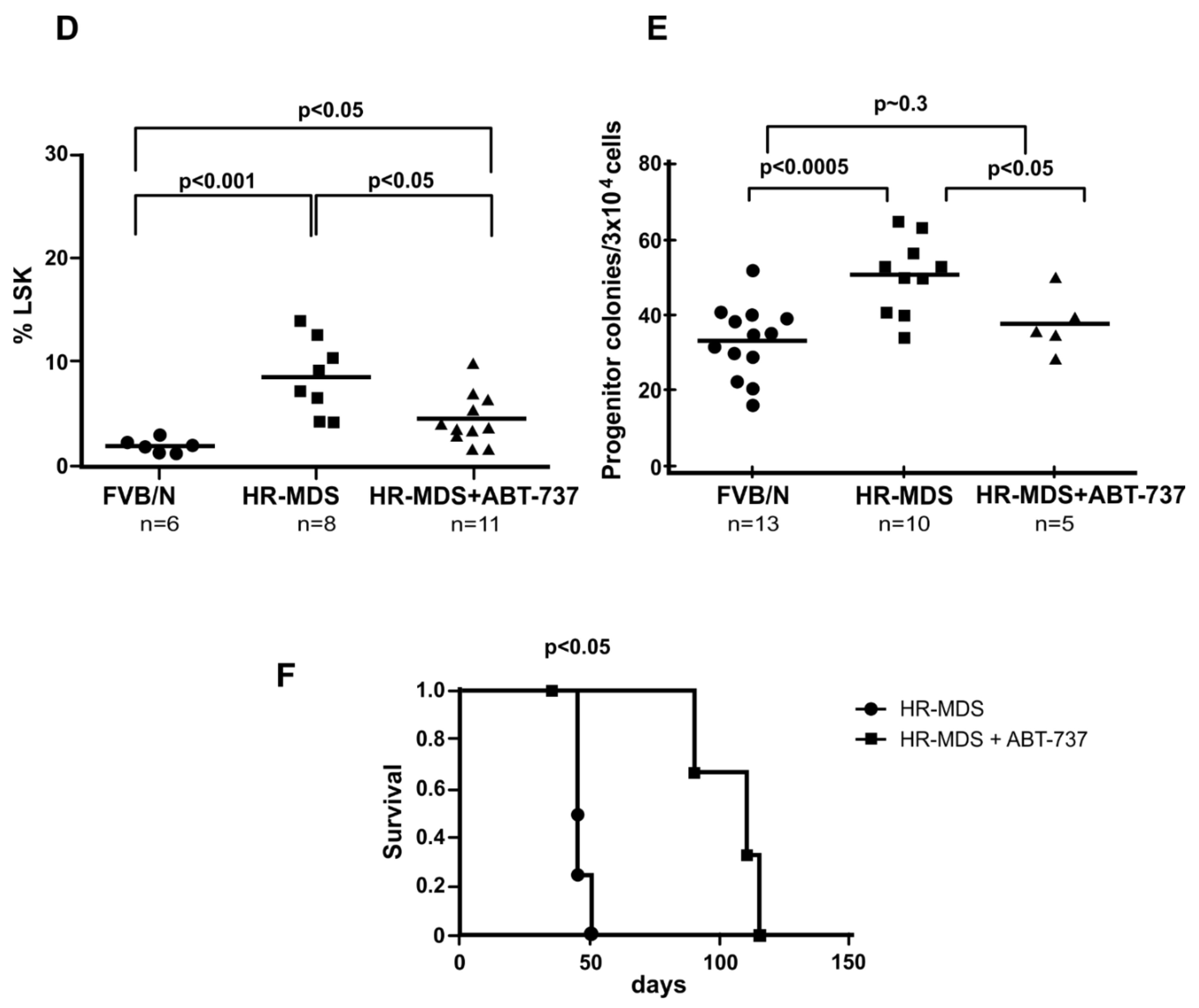
2.2. ABT-737 Treatment Targets Leukemia Initiating Cells (LICs) and Primitive Progenitors
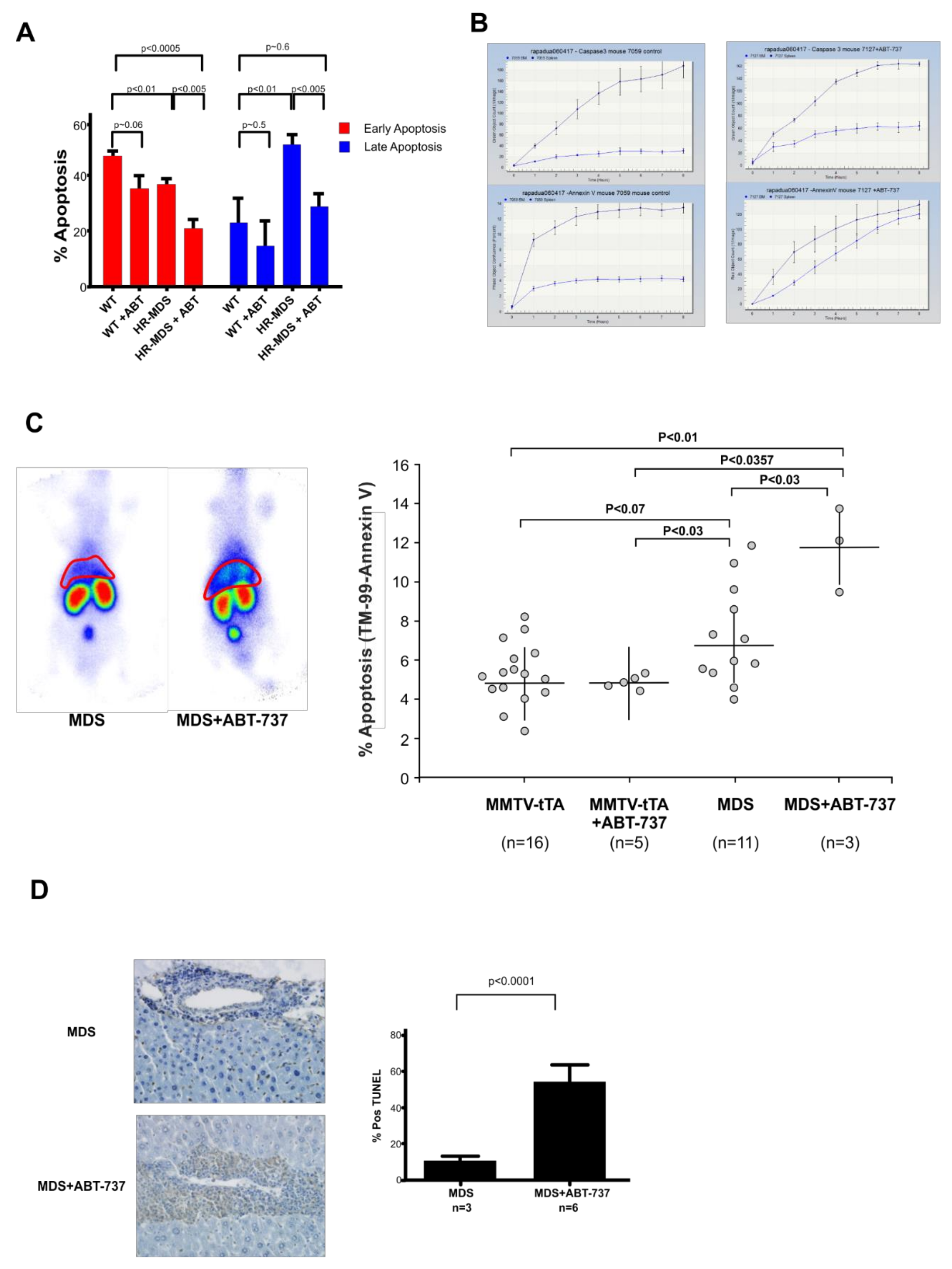
2.3. ABT-737 Treatment Induced Reduced Apoptosis in the BM, Increased Apoptosis and Inhibition of Cell Proliferation in the Liver and Spleen of HR-MDS Transgenic Mice
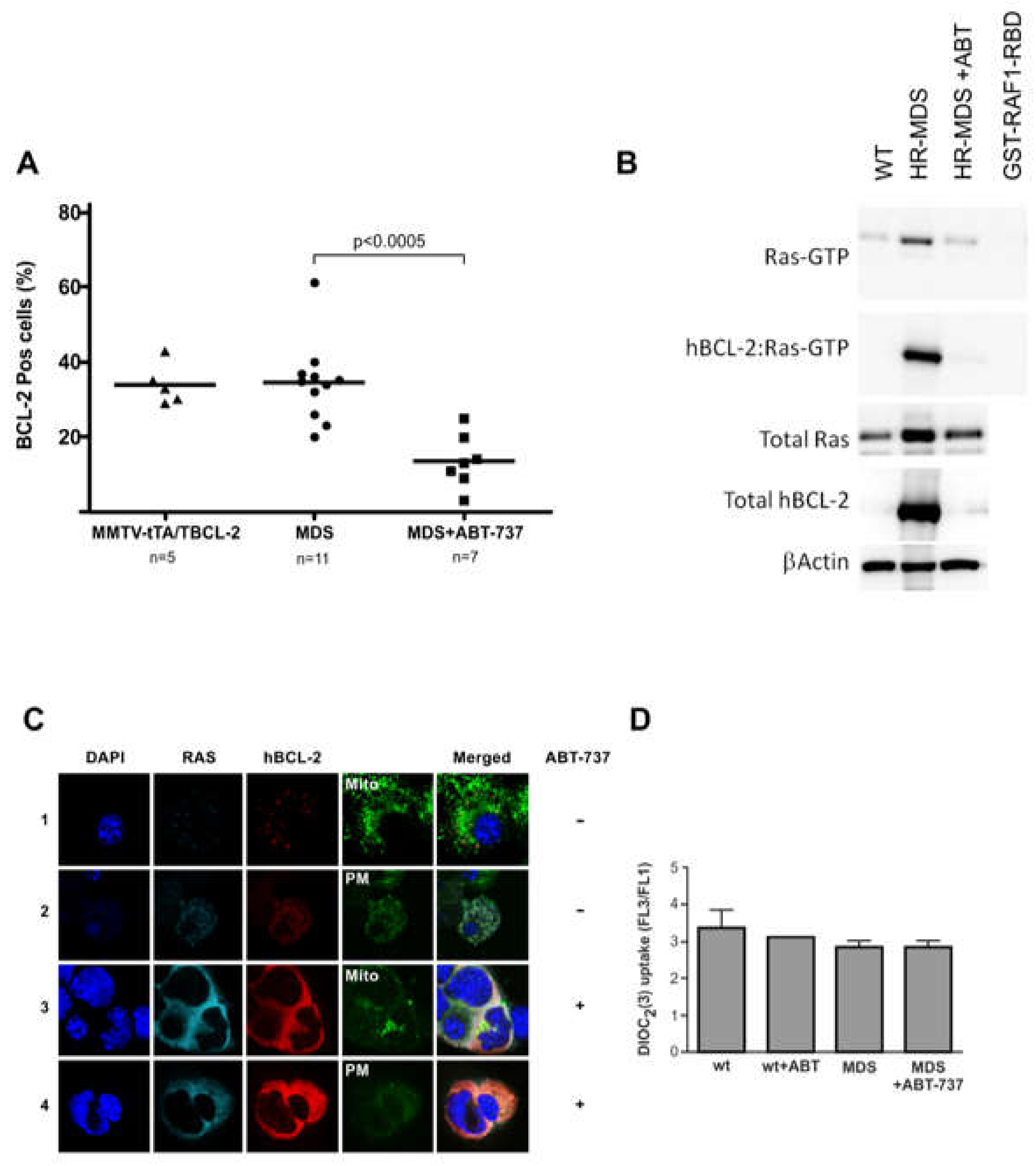
2.4. ABT-737 Induced Inhibition of BCL-2 Reduces RAS Activity in Sca1+ Cells
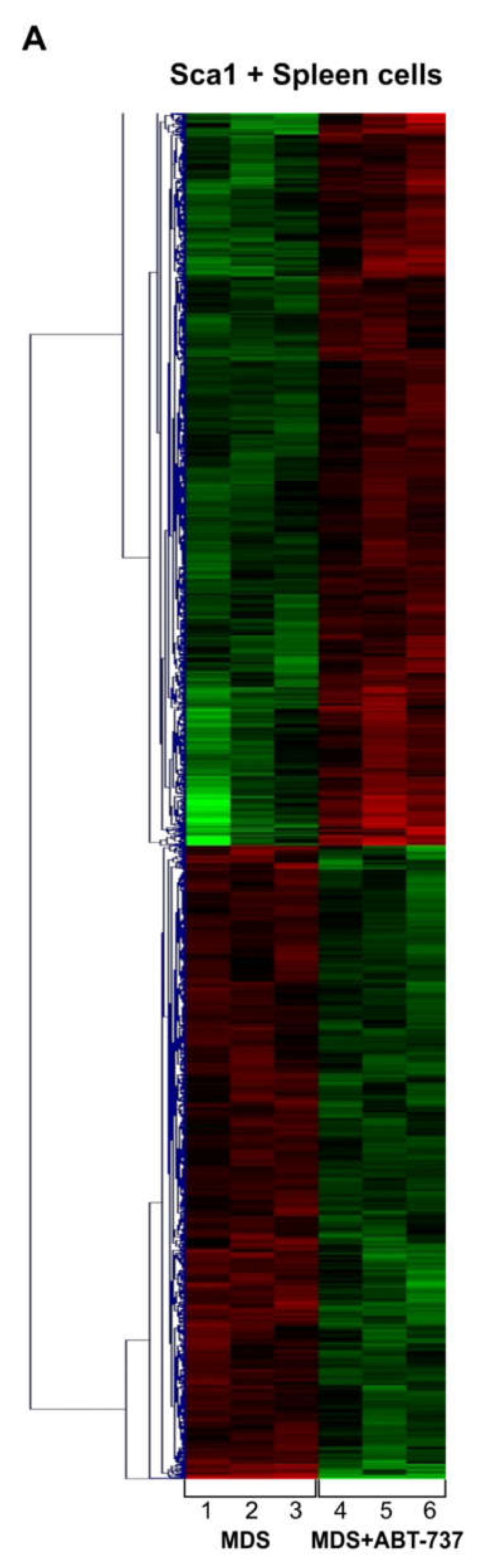
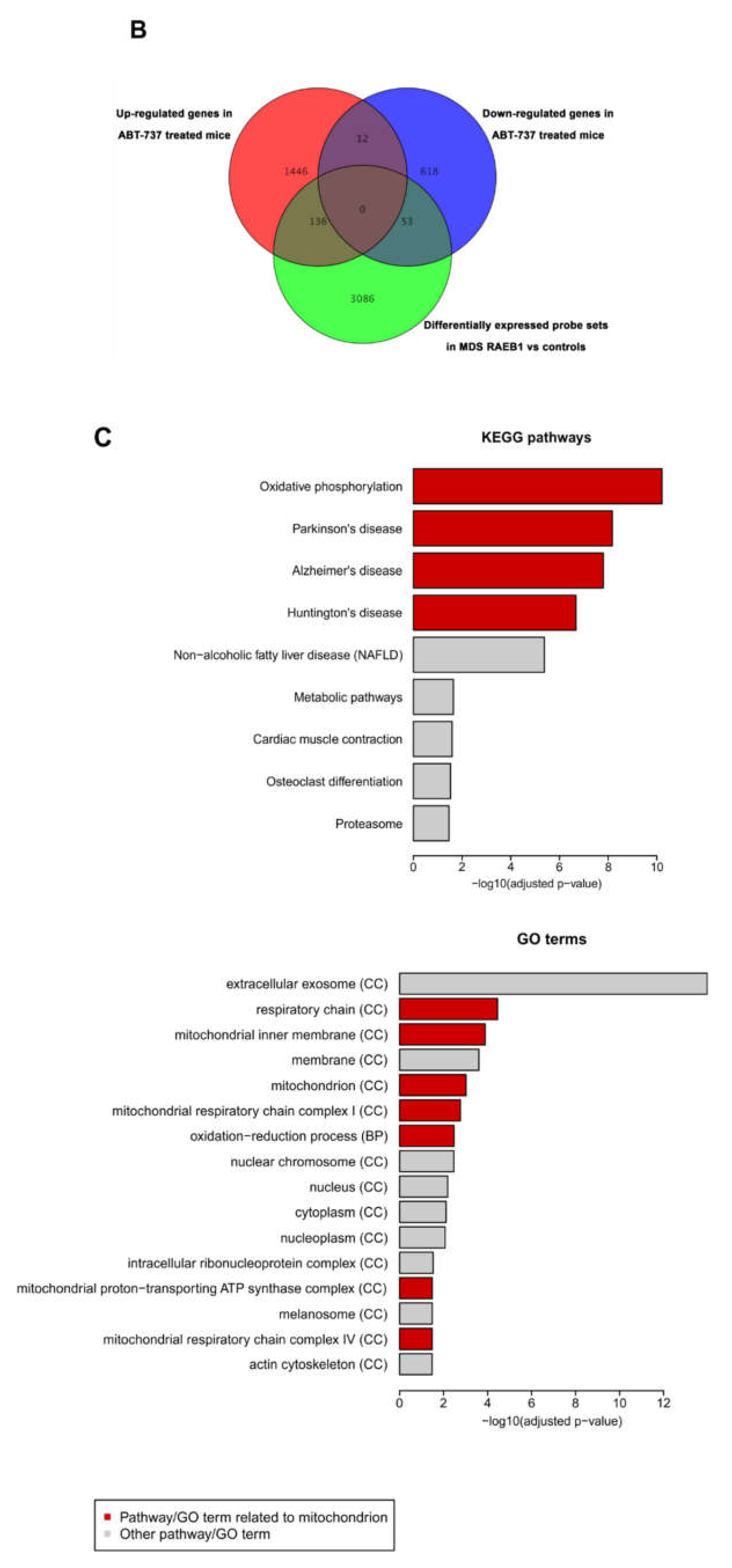
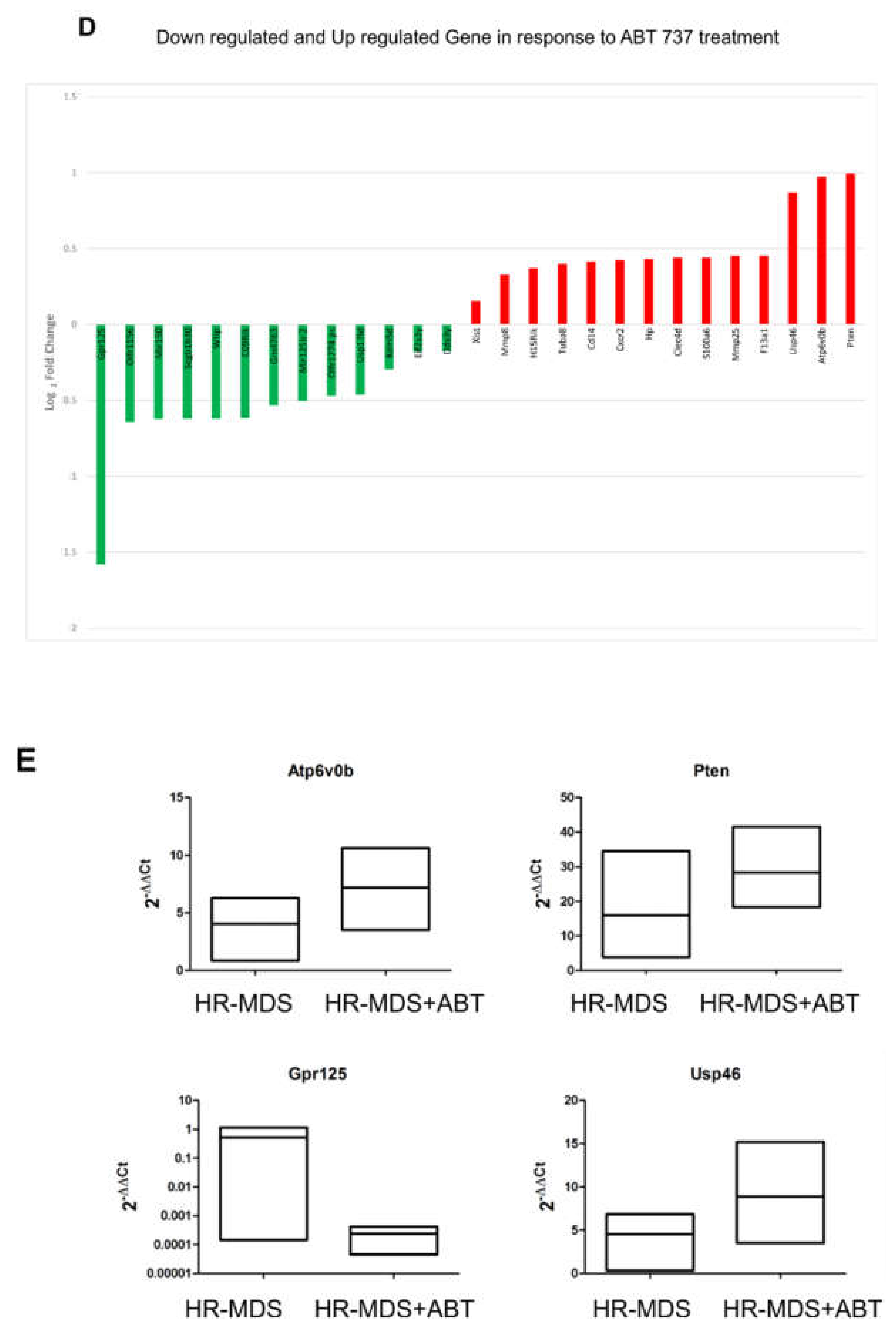
2.5. ABT-737 Treatment Induces Regulation of Pathways Implicated in Cell Survival, Proliferation and Stem Cell Regulation in HR-MDS Mice
3. Discussion
4. Materials and Methods
4.1. Transgenic Mice
4.2. ABT-737
4.3. Tissue and Cell Preparation, Flow Cytometry, Incucyte
4.4. Secondary Transplantation
4.5. Progenitor Colony Assay
4.6. Immunofluorescence and Confocal Microscopy
4.7. Mitochondrial Membrane Potential (MMP)
4.8. SPECT
4.8.1. ANX-Labeling
4.8.2. ANX-Scintigraphy
4.9. TUNEL
4.10. RAS Activation Assays and Western Blotting
4.11. Cell Preparation and RNA Extraction
4.12. Affymetrix Exon Array Hybridization
4.13. Array Data
4.14. RQ-PCR
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Greenberg, P.; Cox, C.; LeBeau, M.M.; Fenaux, P.; Morel, P.; Sanz, G.; Sanz, M.; Vallespi, T.; Hamblin, T.; Oscier, D.; et al. International scoring system for evaluating prognosis in myelodysplastic syndromes. Blood 1997, 89, 2079–2088. [Google Scholar] [CrossRef] [PubMed]
- Padua, R.A.; Guinn, B.A.; Al-Sabah, A.; Smith, M.; Taylor, C.; Pettersson, T.; Ridge, S.; Carter, G.; White, D.; Oscier, D.; et al. RAS, FMS and p53 mutations and poor clinical outcome in myelodysplasias: A 10-year follow-up. Leukemia 1998, 12, 887–892. [Google Scholar] [CrossRef] [Green Version]
- Papaemmanuil, E.; Gerstung, M.; Malcovati, L.; Tauro, S.; Gundem, G.; Loo, P.V.; Yoon, C.J.; Ellis, P.; Wedge, D.; Pellagatti, A.; et al. Clinical and biological implications of driver mutations in myelodysplastic syndromes. Blood 2013, 122, 3616–3627. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.E.; Mufti, G.J.; Rasool, F.; Mijovic, A.; Devereux, S.; Pagliuca, A. The role of apoptosis, proliferation, and the Bcl-2-related proteins in the myelodysplastic syndromes and acute myeloid leukemia secondary to MDS. Blood 2000, 96, 3932–3938. [Google Scholar] [CrossRef]
- Karakas, T.; Maurer, U.; Weidmann, E.; Miething, C.C.; Hoelzer, D.; Bergmann, L. High expression of bcl-2 mRNA as a determinant of poor prognosis in acute myeloid leukemia. Ann. Oncol. 1998, 9, 159–165. [Google Scholar] [CrossRef]
- Karakas, T.; Miething, C.C.; Maurer, U.; Weidmann, E.; Ackermann, H.; Hoelzer, D.; Bergmann, L. The coexpression of the apoptosis-related genes bcl-2 and wt1 in predicting survival in adult acute myeloid leukemia. Leukemia 2002, 16, 846–854. [Google Scholar] [CrossRef] [Green Version]
- Omidvar, N.; Kogan, S.; Beurlet, S.; Le Pogam, C.; Janin, A.; West, R.; Noguera, M.-E.; Reboul, M.; Soulié, A.; Leboeuf, C.; et al. BCL-2 and mutant NRAS interact physically and functionally in a mouse model of progressive myelodysplasia. Cancer Res. 2007, 67, 11657–11667. [Google Scholar] [CrossRef] [Green Version]
- Le Pogam, C.; Krief, P.; Beurlet, S.; Soulie, A.; Balitrand, N.; Cassinat, B.; Cavé, H.; Kosmider, O.; Setterblad, N.; Setterblad, N.; et al. Localization of the NRAS:BCL-2 complex determines anti-apoptotic features associated with progressive disease in myelodysplastic syndromes. Leuk. Res. 2013, 37, 312–319. [Google Scholar] [CrossRef]
- Beurlet, S.; Omidvar, N.; Gorombei, P.; Krief, P.; Le Pogam, C.; Setterblad, N.; De La Grange, P.; Leboeuf, C.; Janin, A.; Noguera, M.-E.; et al. BCL-2 inhibition with ABT-737 prolongs survival in an NRAS/BCL-2 mouse model of AML by targeting primitive LSK and progenitor cells. Blood 2013, 122, 2864–2876. [Google Scholar] [CrossRef]
- Roberts, A.W.; Seymour, J.F.; Brown, J.R.; Wierda, W.G.; Kipps, T.J.; Khaw, S.L.; Carney, D.A.; He, S.Z.; Huang, D.C.; Xiong, H.; et al. Substantial susceptibility of chronic lymphocytic leukemia to BCL-2 inhibition: Results of a phase I study of navitoclax in patients with relapsed or refractory disease. J. Clin. Oncol. 2012, 30, 488–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, W.H.; O’Connor, O.A.; Czuczman, M.S.; LaCasce, A.S.; Gerecitano, J.F.; Leonard, J.P.; Tulpule, A.; Dunleavy, K.; Xiong, H.; Chiu, Y.-L.; et al. Navitoclax, a targeted high-affinity inhibitor of BCL-2, in lymphoid malignancies: A phase 1 dose-escalation study of safety, pharmacokinetics, pharmacodynamics, and antitumour activity. Lancet Oncol. 2010, 11, 1149–1159. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Nimmer, P.M.; Tahir, S.K.; Chen, J.; Fryer, R.M.; Hahn, K.R.; A Iciek, L.; Morgan, S.J.; Nasarre, M.C.; Nelson, R.J.; et al. Bcl-2 family proteins are essential for platelet survival. Cell Death Differ. 2007, 14, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Pellagatti, A.; Cazzola, M.; Giagounidis, A.; Perry, J.; Malcovati, L.; Della Porta, M.G.; Jädersten, M.; Killick, S.; Verma, A.; Norbury, C.J.; et al. Deregulated gene expression pathways in myelodysplastic syndrome hematopoietic stem cells. Leukemia 2010, 24, 756–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Delft, M.F.; Wei, A.H.; Mason, K.D.; Vandenberg, C.J.; Chen, L.; Czabotar, P.E.; Willis, S.N.; Scott, C.L.; Day, C.; Cory, S.; et al. The BH3 mimetic ABT-737 targets selective Bcl-2 proteins and efficiently induces apoptosis via Bak/Bax if Mcl-1 is neutralized. Cancer Cell 2006, 10, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Konopleva, M.; Contractor, R.; Tsao, T.; Samudio, I.; Ruvolo, P.P.; Kitada, S.; Deng, X.; Zhai, D.; Shi, Y.-X.; Sneed, T.; et al. Mechanisms of apoptosis sensitivity and resistance to the BH3 mimetic ABT-737 in acute myeloid leukemia. Cancer Cell 2006, 10, 375–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, K.D.; Vandenberg, C.J.; Scott, C.L.; Wei, A.H.; Cory, S.; Huang, D.C.; Roberts, A.W. In vivo efficacy of the Bcl-2 antagonist ABT-737 against aggressive Myc-driven lymphomas. Proc. Natl. Acad. Sci. USA 2008, 105, 17961–17966. [Google Scholar] [CrossRef] [Green Version]
- Hann, C.L.; Daniel, V.C.; Sugar, E.A.; Dobromilskaya, I.; Murphy, S.C.; Cope, L.; Lin, X.; Hierman, J.S.; Wilburn, D.L.; Watkins, D.N.; et al. Therapeutic Efficacy of ABT-737, a Selective Inhibitor of BCL-2, in Small Cell Lung Cancer. Cancer Res. 2008, 68, 2321–2328. [Google Scholar] [CrossRef] [Green Version]
- Andreu-Fernandez, V.; Genoves, A.; Messeguer, A.; Orzaez, M.; Sancho, M.; Perez-Paya, E. BH3-mimetics- and cisplatin-induced cell death proceeds through different pathways depending on the availability of death-related cellular components. PLoS ONE 2013, 8, e56881. [Google Scholar] [CrossRef] [Green Version]
- Gersuk, G.M.; Lee, J.W.; Beckham, C.A.; Anderson, J.; Deeg, H.J. Fas (CD95) receptor and Fas-ligand expression in bone marrow cells from patients with myelodysplastic syndrome. Blood 1996, 88, 1122–1123. [Google Scholar] [CrossRef]
- Hyde, R.K.; Kamikubo, Y.; Anderson, S.; Kirby, M.; Alemu, L.; Zhao, L.; Liu, P.P. Cbfb/Runx1 repression-independent blockage of differentiation and accumulation of Csf2rb-expressing cells by Cbfb-MYH11. Blood 2010, 115, 1433–1443. [Google Scholar] [CrossRef] [Green Version]
- Fleischman, A.G. ALDH marks leukemia stem cell. Blood 2012, 119, 3376–3377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, Y.P.; Bultynck, G.; Aromolaran, A.S.; Zhong, F.; Parys, J.B.; Smedt, D.H.; Mignery, G.A.; Roderick, H.L.; Bootman, M.T.; Distelhorst, C.W. The BH4 domain of Bcl-2 inhibits ER calcium release and apoptosis by binding the regulatory and coupling domain of the IP3 receptor. Proc. Natl. Acad. Sci. USA 2009, 106, 14397–14402. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, Y.; Kikuchi, J.; Nakamura, M.; Iwase, S.; Yamada, H.; Matsuda, M. Lineage-specific regulation of cell cycle control gene expression during haematopoietic cell differentiation. Br. J. Haematol. 2000, 110, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Shen, H.; Jiang, H.; Hu, D.; Wang, J.; Wu, X. External Qi of Yan Xin Qigong inhibits activation of Akt, Erk1/2 and NF-kB and induces cell cycle arrest and apoptosis in colorectal cancer cells. Cell Physiol. Biochem. 2013, 31, 113–122. [Google Scholar] [CrossRef]
- Janku, F.; Lee, J.J.; Tsimberidou, A.M.; Hong, D.S.; Naing, A.; Falchook, G.S.; Fu, S.; Luthra, R.; Garrido-Laguna, I.; Kurzrock, R. PIK3CA Mutations Frequently Coexist with RAS and BRAF Mutations in Patients with Advanced Cancers. PLoS ONE 2011, 6, e22769. [Google Scholar] [CrossRef] [Green Version]
- Talab, F.; Allen, J.C.; Thompson, V.; Lin, K.; Slupsky, J.R. LCK is an important mediator of B-cell receptor signaling in chronic lymphocytic leukemia cells. Mol. Cancer Res. 2013, 11, 541–554. [Google Scholar] [CrossRef] [Green Version]
- Gandhi, L.; Camidge, D.R.; Ribeiro De Oliveira, M.M.R.; Bonomi, P.; Gandara, D.; Khaira, D.; Hann, C.L.; McKeegan, E.M.; Litvinovich, E.; Hemken, P.M.; et al. Phase I Study of Navitoclax (ABT-263), a Novel Bcl-2 Family Inhibitor, in Patients With Small-Cell Lung Cancer and Other Solid Tumors. J. Clin. Oncol. 2011, 29, 909–916. [Google Scholar] [CrossRef] [Green Version]
- Hallek, M.; Pflug, N. State of the art treatment of chronic lymphocytic leukaemia. Blood Rev. 2011, 25, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tse, C.; Shoemaker, A.R.; Adickes, J.; Anderson, M.G.; Chen, J.; Jin, S.; Johnson, E.F.; Marsh, K.C.; Mitten, M.J.; Nimmer, P.; et al. ABT-263: A Potent and Orally Bioavailable Bcl-2 Family Inhibitor. Cancer Res. 2008, 68, 3421–3428. [Google Scholar] [CrossRef] [Green Version]
- Kallam, A.; Armitage, J.O. Venetoclax in chronic lymphocytic leukaemia: A possible cure? Lancet Oncol. 2018, 19, 1143–1144. [Google Scholar] [CrossRef]
- Valentin, R.; Grabow, S.; Davids, M.S. The rise of apoptosis: Targeting apoptosis in hematologic malignancies. Blood 2018, 132, 1248–1264. [Google Scholar] [CrossRef] [Green Version]
- Souers, A.J.; Leverson, J.D.; Boghaert, E.R.; Ackler, S.L.; Catron, N.D.; Chen, J.; Dayton, B.D.; Ding, H.; Enschede, S.H.; Fairbrother, W.J.; et al. ABT-199, a potent and selective BCL-2 inhibitor, achieves antitumor activity while sparing platelets. Nat. Med. 2013, 19, 202–208. [Google Scholar] [CrossRef] [PubMed]
- DiNardo, C.D.; Pratz, K.W.; Letai, A.; Jonas, B.; Wei, A.H.; Thirman, M.; Arellano, M.; Frattini, M.G.; Kantarjian, H.; Popovic, R.; et al. Safety and preliminary efficacy of venetoclax with decitabine or azacitidine in elderly patients with previously untreated acute myeloid leukaemia: A non-randomised, open-label, phase 1b study. Lancet Oncol. 2018, 19, 216–228. [Google Scholar] [CrossRef]
- Hennighausen, L.; Wall, R.J.; Tillmann, U.; Li, M.; Furth, P.A. Conditional gene expression in secretory tissues and skin of transgenic mice using the MMTV-LTR and the tetracycline responsive system. J. Cell Biochem. 1995, 59, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Kogan, S.C.; Ward, J.M.; Anver, M.R.; Berman, J.J.; Brayton, C.; Cardiff, R.D.; Carter, J.S.; de Coronado, S.; Downing, J.R.; Fredrickson, T.N.; et al. Bethesda proposals for classification of nonlymphoid hematopoietic neoplasms in mice. Blood 2002, 100, 238–245. [Google Scholar] [CrossRef] [Green Version]
- Guerenne, L.; Beurlet, S.; Said, M.; Gorombei, P.; Le Pogam, C.; Guidez, F.; De La Grange, P.; Omidvar, N.; Vanneaux, V.; Mills, K.; et al. GEP analysis validates high risk MDS and acute myeloid leukemia post MDS mice models and highlights novel dysregulated pathways. J. Hematol. Oncol. 2016, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Novo, D.; Perlmutter, N.G.; Hunt, R.H.; Shapiro, H.M. Accurate flow cytometric membrane potential measurement in bacteria using diethyloxacarbocyanine and a ratiometric technique. Cytometry 1999, 35, 55–63. [Google Scholar] [CrossRef]
- De la Grange, P.; Gratadou, L.; Delord, M.; Dutertre, M.; Auboeuf, D. Splicing factor and exon profiling across human tissues. Nucleic. Acids. Res. 2010, 38, 2825–2838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Grange, P.; Dutertre, M.; Martin, N.; Auboeuf, D. FAST DB: A website resource for the study of the expression regulation of human gene products. Nucleic. Acids. Res. 2005, 33, 4276–4284. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S.; Sato, Y.; Furumichi, M.; Tanabe, M. KEGG for integration and interpretation of large-scale molecular data sets. Nucleic. Acids. Res. 2012, 40, D109–D114. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- High, L.M.; Szymanska, B.; Wilczynska-Kalak, U.; Barber, N.; O’Brien, R.; Khaw, S.L.; Vikstrom, I.B.; Roberts, A.W.; Lock, R.B. The Bcl-2 Homology Domain 3 Mimetic ABT-737 Targets the Apoptotic Machinery in Acute Lymphoblastic Leukemia Resulting in Synergistic in Vitro and in Vivo Interactions with Established Drugs. Mol. Pharmacol. 2009, 77, 483–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olberding, K.E.; Wang, X.; Zhu, Y.; Pan, J.; Rai, S.N.; Li, C. Actinomycin D synergistically enhances the efficacy of the BH3 mimetic ABT-737 by downregulating Mcl-1 expression. Cancer Biol. Ther. 2010, 10, 918–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Cai, T.-Y.; Zhu, H.; Yang, L.-Q.; Jiang, H.; Dong, X.-W.; Hu, Y.-Z.; Lin, N.-M.; He, Q.-J.; Yang, B. Synergistic Antitumor Activity of Gemcitabine and ABT-737 In Vitro and In Vivo through Disrupting the Interaction of USP9X and Mcl-1. Mol. Cancer Ther. 2011, 10, 1264–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, L.A.; Goldstein, N.; Johannes, W.U.; Walton, C.H.; Fujita, M.; Norris, D.A.; Shellman, Y.G. BH3 Mimetic ABT-737 and a Proteasome Inhibitor Synergistically Kill Melanomas through Noxa-Dependent Apoptosis. J. Investig. Dermatol. 2009, 129, 964–971. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Konopleva, M.; Ruvolo, V.R.; McQueen, T.; Evans, R.L.; Bornmann, W.G.; McCubrey, J.; Cortes, J.; Andreeff, M. Sorafenib induces apoptosis of AML cells via Bim-mediated activation of the intrinsic apoptotic pathway. Leukemia 2008, 22, 808–818. [Google Scholar] [CrossRef]
- Konopleva, M.; Letai, A. BCL-2 inhibition in AML: An unexpected bonus? Blood 2018, 132, 1007–1012. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Apoptosis related genes regulated | |||
|---|---|---|---|
| Gene symbol | Regulation | Fold change | p value |
| Sgk1 | up | 1.67 | 2.05 × 10−2 |
| E2f2 | up | 2.06 | 2.12 × 10−2 |
| Osm | up | 2.26 | 3.18 × 10−2 |
| Rybp | up | 1.85 | 3.48 × 10−2 |
| Ripk3 | up | 1.68 | 3.62 × 10−2 |
| Rnf130 | up | 1.57 | 4.72 × 10−3 |
| Gpx1 | up | 1.76 | 7.96 × 10−3 |
| Bcl2l12 | up | 1.79 | 1.63 × 10−2 |
| App | up | 2.46 | 3.79 × 10−2 |
| Tnfrsf21 | up | 2.02 | 4.42 × 10−2 |
| Pdcd5 | up | 1.65 | 1.82 × 10−2 |
| Apaf1 | up | 1.62 | 3.56 × 10−2 |
| Mfsd10 | up | 1.53 | 1.33 × 10−2 |
| Pdcl3 | up | 1.50 | 2.26 × 10−3 |
| Ckap2 | up | 1.91 | 2.50 × 10−2 |
| Casp1 | up | 1.77 | 2.25 × 10−3 |
| Bcl2a1a | up | 2.00 | 2.85 × 10−2 |
| G2e3 | up | 1.89 | 1.74 × 10−3 |
| Birc5 | up | 2.45 | 2.10 × 10−2 |
| C1d | up | 1.52 | 1.07 × 10−2 |
| Sgpl1 | up | 1.56 | 3.36 × 10−2 |
| Bak1 | up | 1.97 | 1.49 × 10−2 |
| Hipk2 | up | 1.51 | 3.38 × 10−2 |
| Naip2 | up | 1.81 | 2.77 × 10−3 |
| Rnf144b | up | 1.51 | 1.28 × 10−2 |
| Pten | up | 1.98 | 4.07 × 10−3 |
| Csf2rb | up | 2.48 | 1.18 × 10−2 |
| Pik3cg | up | 1.62 | 3.48 × 10−3 |
| Chp1 | up | 1.85 | 5.80 × × 10−4 |
| Csf2rb2 | up | 1.73 | 2.07 × 10−2 |
| Cxcr2 | up | 4.98 | 1.37 × 10−2 |
| Wwox | down | 1.60 | 5.05 × 10−3 |
| Eif2ak3 | down | 1.54 | 9.38 × 10−3 |
| Fasl | down | 2.33 | 3.27 × 10−2 |
| Lck | down | 1.68 | 3.46 × 10−2 |
| Dyrk2 | down | 1.63 | 8.96 × × 10−4 |
| Trib3 | down | 1.51 | 1.59 × 10−2 |
| Sod1 | down | 1.54 | 4.83 × × 10−4 |
| Fas | down | 1.54 | 2.01 × 10−2 |
| Csrnp2 | down | 1.7 | 2.16 × 10−2 |
| Splice related genes regulated | |||
| Gene Symbol | Regulation | Fold-Change | p-Value |
| Rsrc1 | up | 1.55 | 3.59 × 10−2 |
| Gemin6 | up | 1.97 | 2.50 × 10−2 |
| Gemin7 | up | 1.66 | 5.08 × 10−3 |
| Lgals3 | up | 4.29 | 1.75 × 10−2 |
| Lsm6 | up | 1.62 | 3.69 × 10−2 |
| Zrsr2 | up | 1.71 | 4.37 × 10−2 |
| Snrnp27 | up | 1.59 | 3.39 × 10−2 |
| Snrpd2 | up | 1.76 | 4.59 × 10−2 |
| Lsm10 | up | 1.90 | 1.28 × 10−3 |
| Wbp4 | up | 1.85 | 1.67 × 10−2 |
| Isy1 | down | 1.62 | 3.51 × 10−2 |
| Pnn | down | 1.91 | 1.19 × 10−2 |
| Prpf38b | down | 1.54 | 1.56 × 10−2 |
| Rbm20 | down | 1.73 | 1.36 × 10−2 |
| Rbfox1 | down | 1.50 | 6.72 × 10−3 |
| Snrnp48 | down | 1.50 | 3.85 × 10−2 |
| Tut1 | down | 1.73 | 2.19 × 10−2 |
| Cell cycle related genes regulated | |||
| Gene symbol | Regulation | Fold change | p value |
| Cdk2 | up | 1.72 | 1.10 × 10−2 |
| Ccne2 | up | 1.76 | 2.18 × 10−2 |
| Cdc6 | up | 1.51 | 4.07 × 10−2 |
| E2f2 | up | 2.06 | 2.12 × 10−2 |
| Mapk13 | up | 3.06 | 3.22 × 10−2 |
| Ccnb2 | up | 2.25 | 3.62 × 10−2 |
| Cdca8 | up | 1.90 | 4.16 × 10−2 |
| Ccna2 | up | 2.51 | 2.04 × 10−2 |
| Cdk1 | up | 2.27 | 9.20 × 10−3 |
| Cdkn2d | up | 2.01 | 5.12 × 10−3 |
| Cdkn2c | up | 2.08 | 1.20 × 10−2 |
| Ccnd3 | up | 1.88 | 2.09 × 10−2 |
| Cdca3 | up | 2.24 | 1.91 × 10−2 |
| Cdc25a | up | 1.92 | 1.95 × 10−2 |
| Cdkn3 | up | 2.43 | 2.91 × 10−2 |
| Mapk3 | up | 1.89 | 2.36 × 10−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorombei, P.; Guidez, F.; Ganesan, S.; Chiquet, M.; Pellagatti, A.; Goursaud, L.; Tekin, N.; Beurlet, S.; Patel, S.; Guerenne, L.; et al. BCL-2 Inhibitor ABT-737 Effectively Targets Leukemia-Initiating Cells with Differential Regulation of Relevant Genes Leading to Extended Survival in a NRAS/BCL-2 Mouse Model of High Risk-Myelodysplastic Syndrome. Int. J. Mol. Sci. 2021, 22, 10658. https://doi.org/10.3390/ijms221910658
Gorombei P, Guidez F, Ganesan S, Chiquet M, Pellagatti A, Goursaud L, Tekin N, Beurlet S, Patel S, Guerenne L, et al. BCL-2 Inhibitor ABT-737 Effectively Targets Leukemia-Initiating Cells with Differential Regulation of Relevant Genes Leading to Extended Survival in a NRAS/BCL-2 Mouse Model of High Risk-Myelodysplastic Syndrome. International Journal of Molecular Sciences. 2021; 22(19):10658. https://doi.org/10.3390/ijms221910658
Chicago/Turabian StyleGorombei, Petra, Fabien Guidez, Saravanan Ganesan, Mathieu Chiquet, Andrea Pellagatti, Laure Goursaud, Nilgun Tekin, Stephanie Beurlet, Satyananda Patel, Laura Guerenne, and et al. 2021. "BCL-2 Inhibitor ABT-737 Effectively Targets Leukemia-Initiating Cells with Differential Regulation of Relevant Genes Leading to Extended Survival in a NRAS/BCL-2 Mouse Model of High Risk-Myelodysplastic Syndrome" International Journal of Molecular Sciences 22, no. 19: 10658. https://doi.org/10.3390/ijms221910658