
Non-Canonical Helical Structure of Nucleic Acids Containing Base-Modified Nucleotides

 ,
,
Abstract
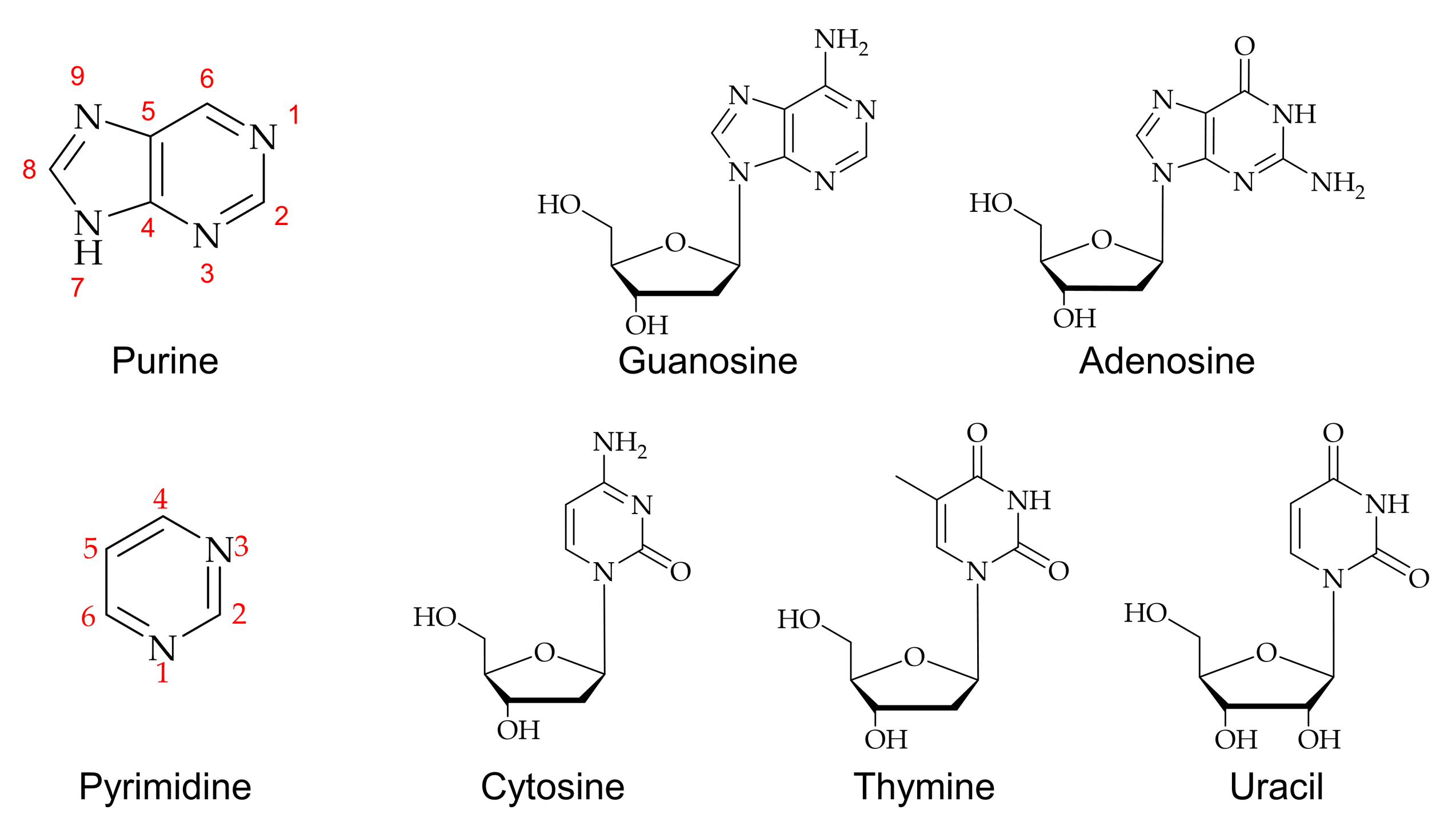
:1. Introduction
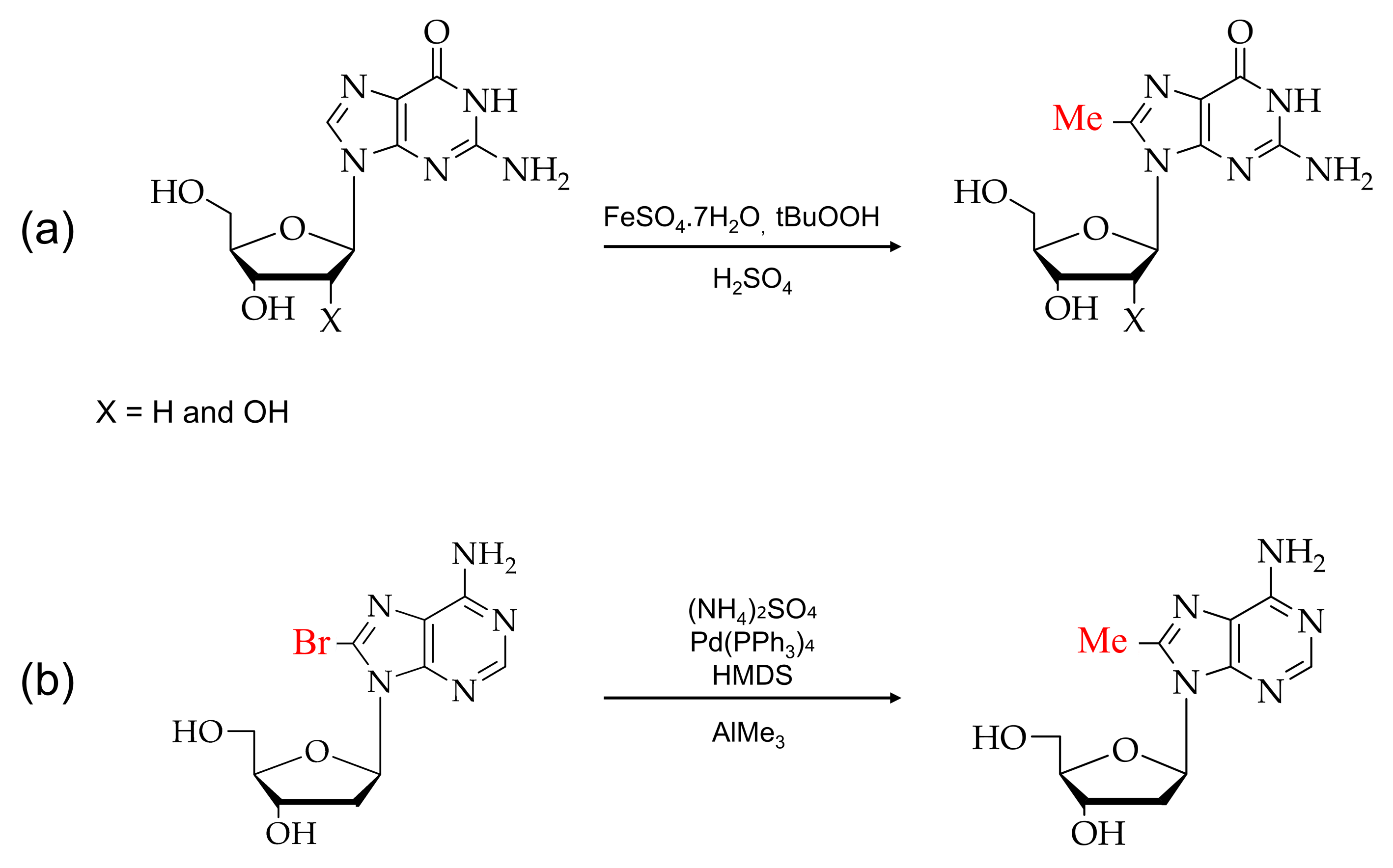
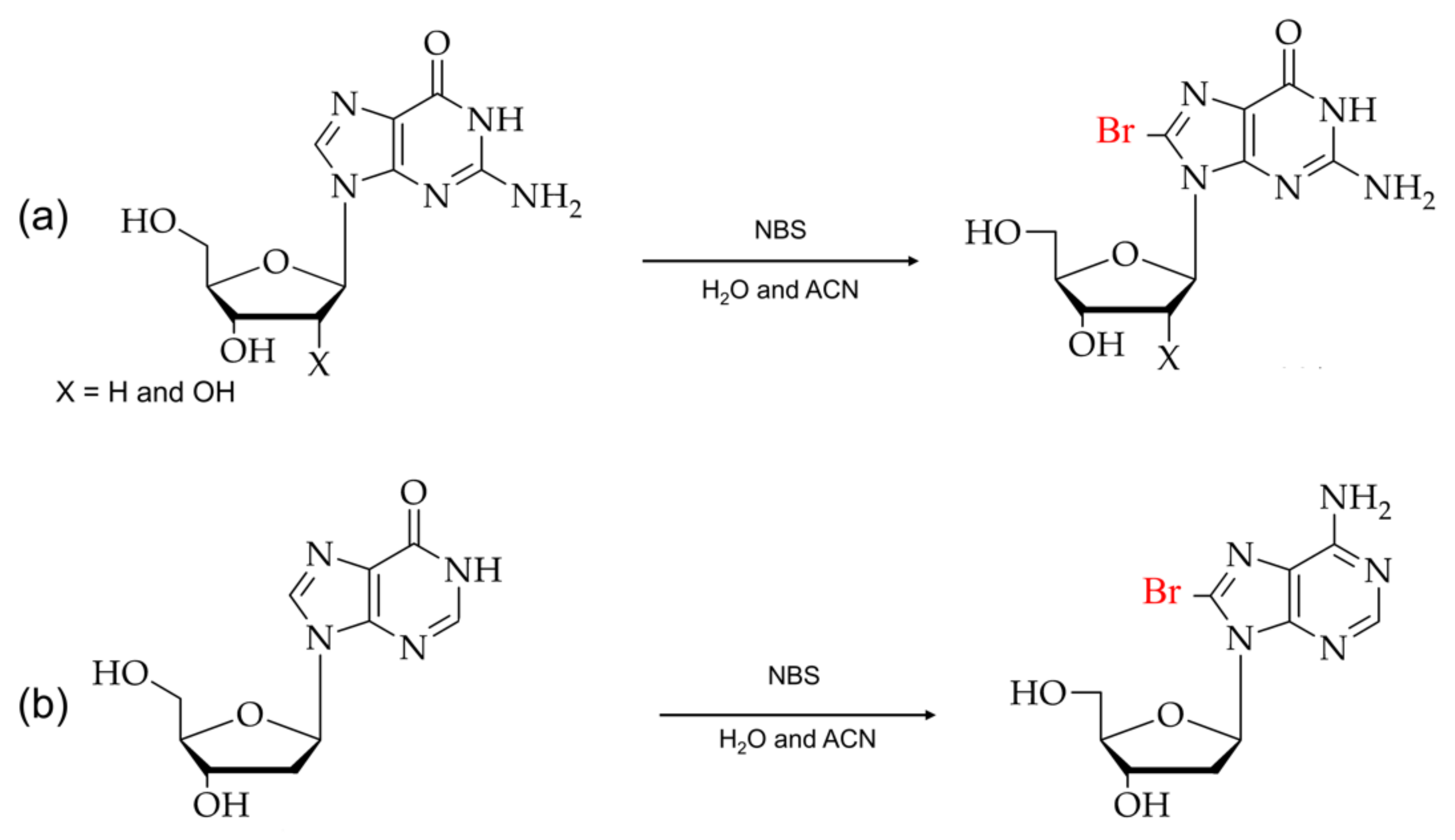
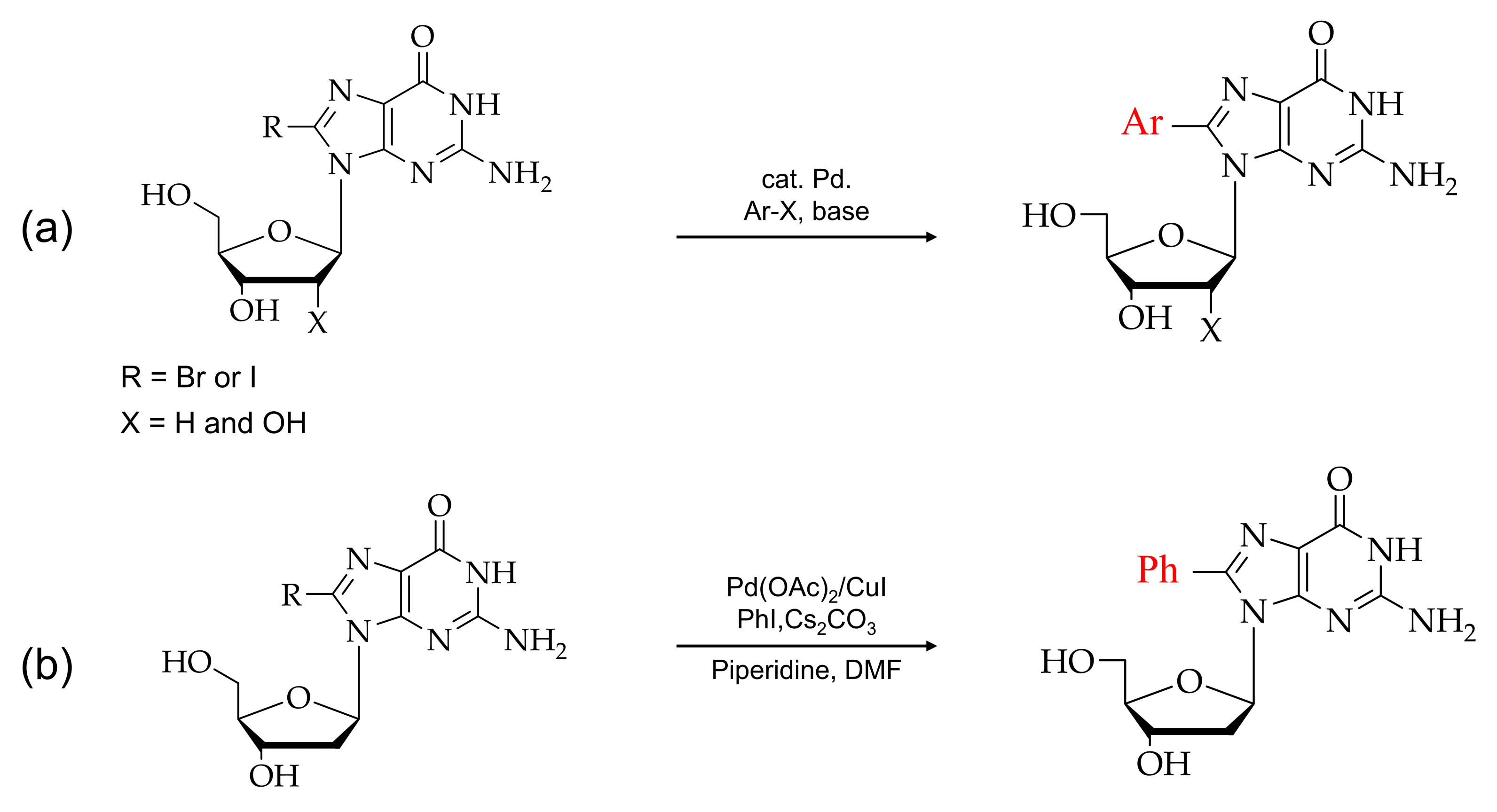
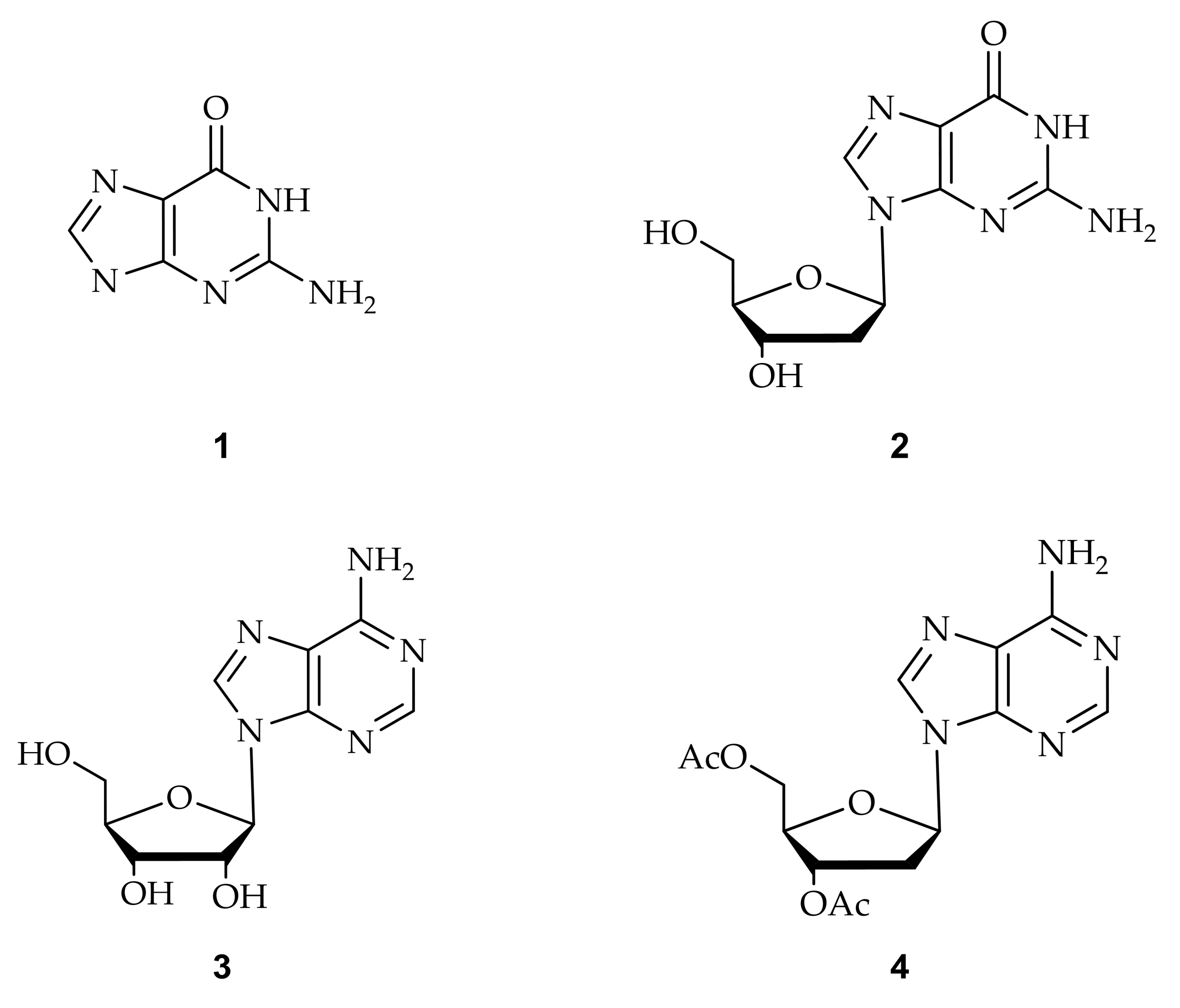
2. Synthesis of C8-Modified Purine
2.1. Synthesis of C8-Methylpurine
2.2. Synthesis of C8-Bromopurine
2.3. Synthesis of C8-Arylpurine
3. Non-Canonical Structure Containing C8-Modified Guanosine
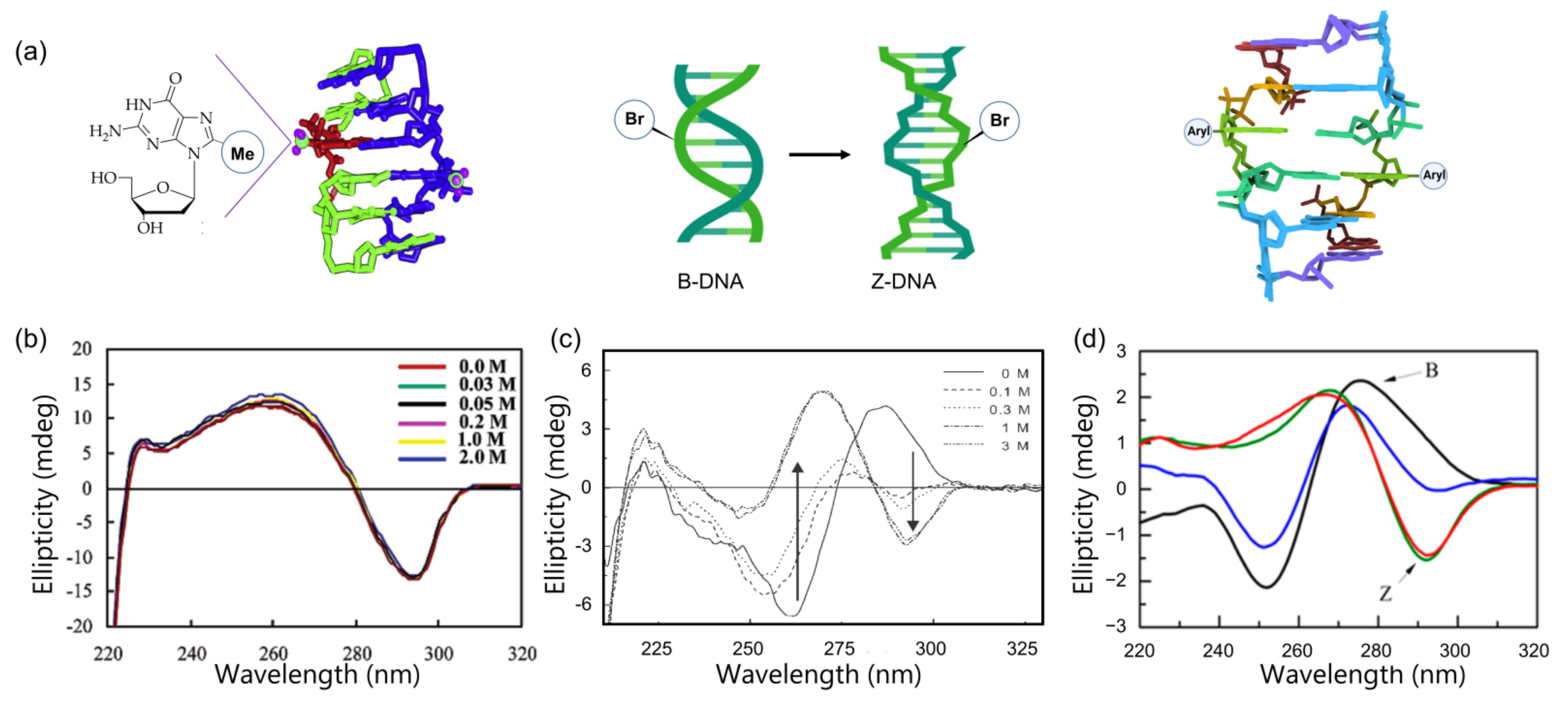
3.1. Z-DNA
3.1.1. C8-Methylation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Sequence | Modification | Midpoint of (B–Z Transition) NaCl Concentration (mM) | Reference |
|---|---|---|---|---|
| 1 | d(CGCm8rGCG)2 | m8rG | 0 | [79] |
| 2 | d(CGCFGCG)2 | FG | 20 | [88] |
| 3 | d(CGCm8mGCG)2 | m8mG | 0 | [86] |
| 4 | d(CGCm8GCG)2 | m8G | 30 | [90] |
| 5 | Poly(dG-dC) | Br-Poly(dG-dC) | 1000 | [91] |
3.1.2. C8-Bromination
3.1.3. C8-Arylation
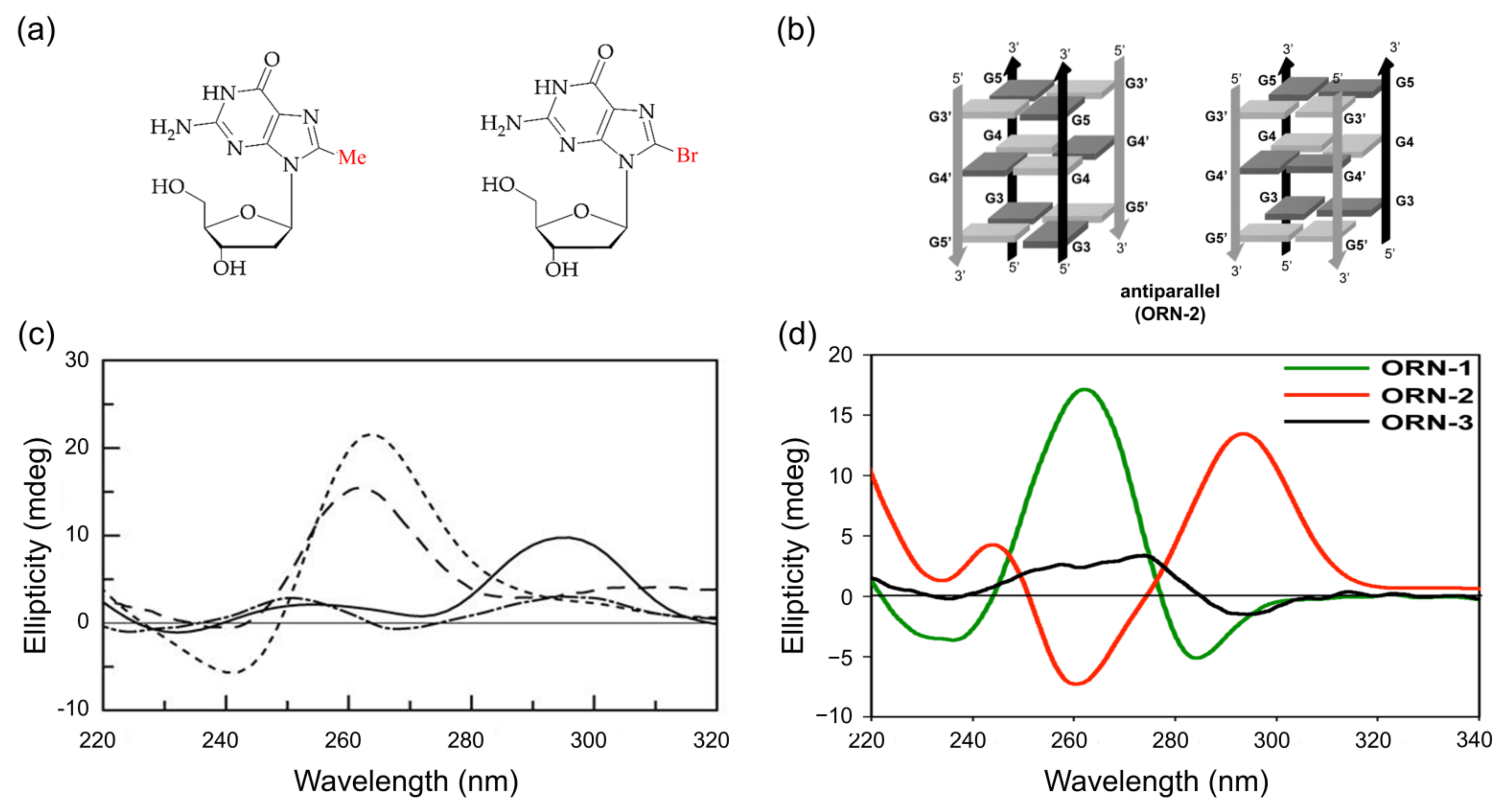
3.2. G-Quadruplex
3.2.1. C8-Methylation
3.2.2. C8-Bromination
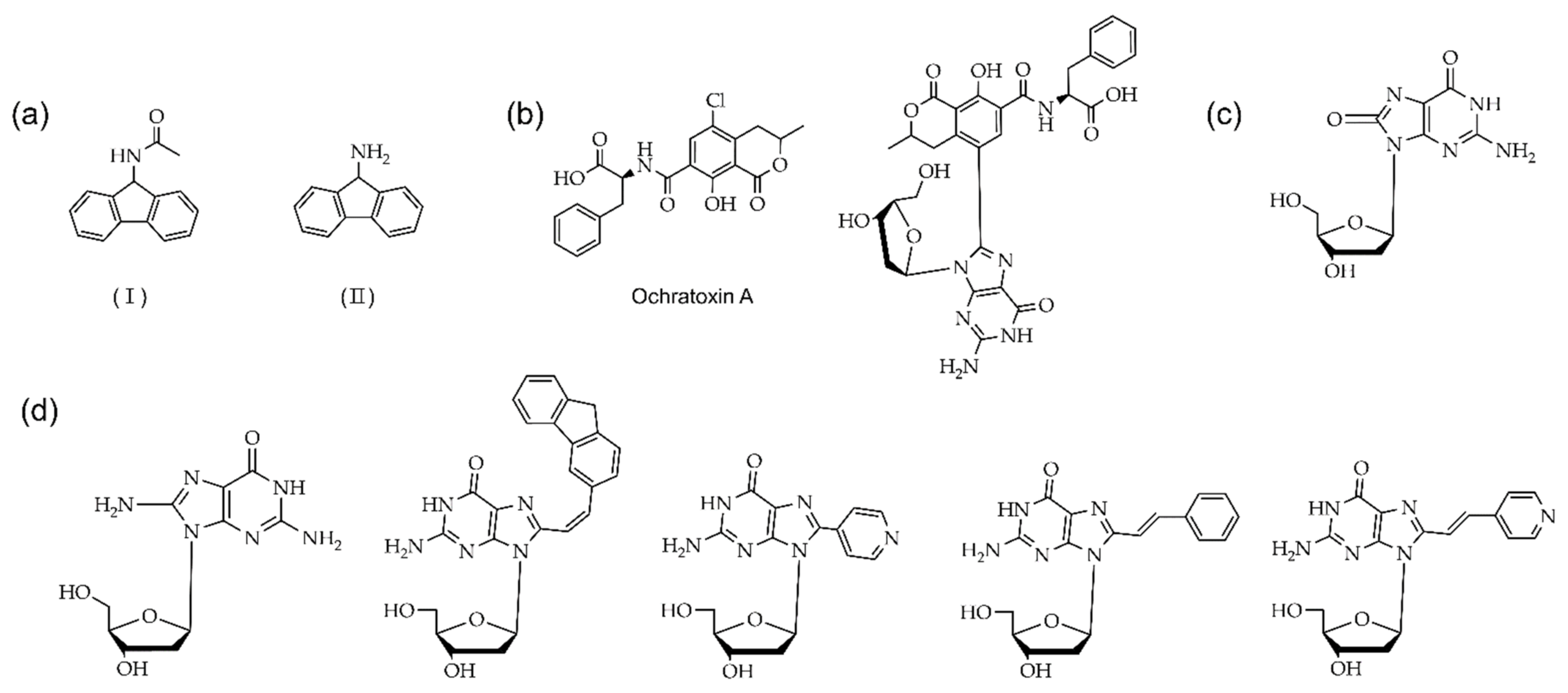
3.2.3. Other Modifications at the C8 Position
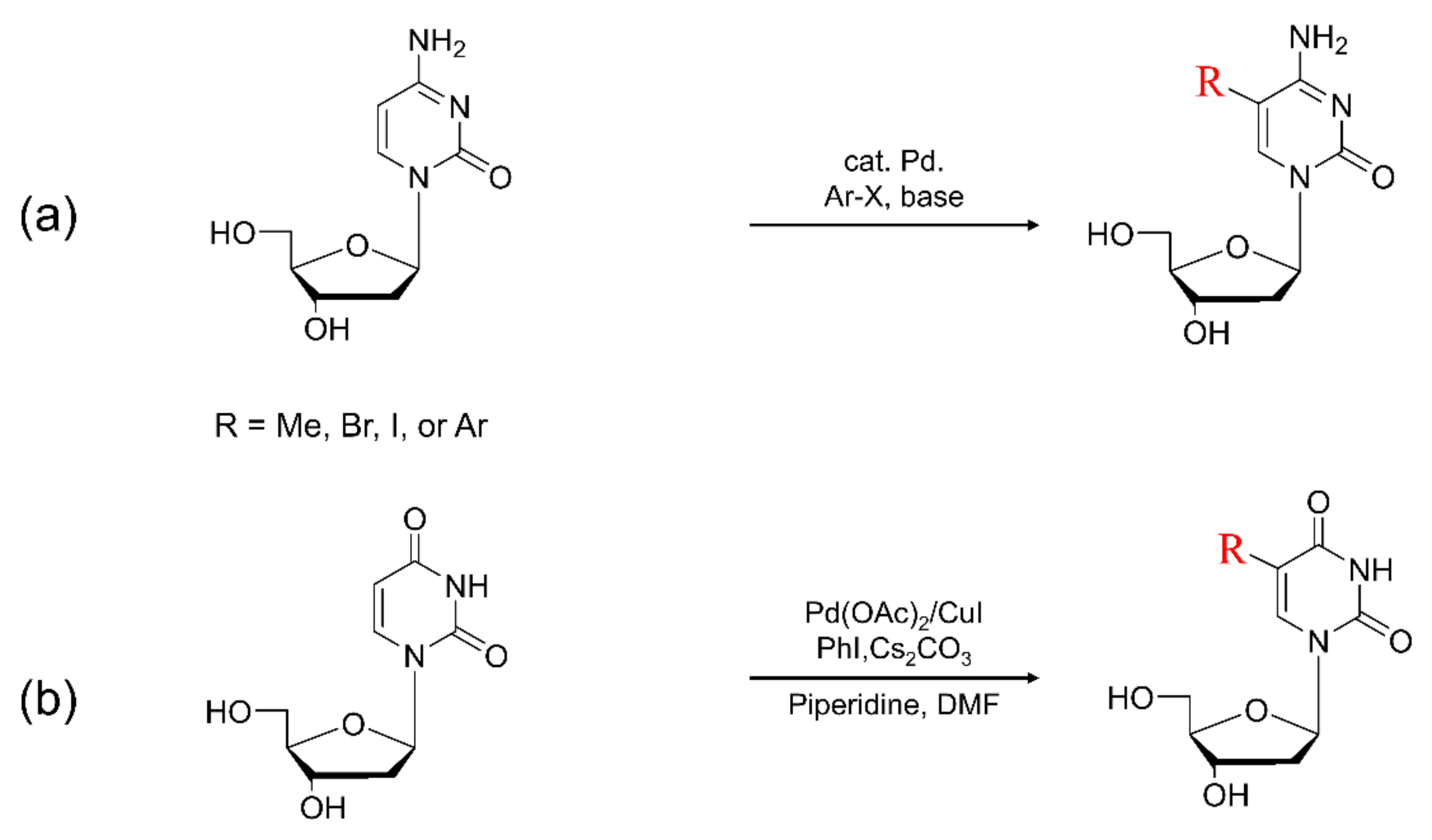
4. Synthesis of C5-Modified Pyrimidine
4.1. Synthesis of C5-Methylpyrimidine
4.2. Synthesis of C5-Bromopyrimidine
4.3. Synthesis of C5-Arylpyrimidine
5. Non-Canonical Structure Containing C5-Modified Pyrimidine
5.1. Z-DNA
5.1.1. C5-Methylation
5.1.2. C5-Bromination
5.2. G-Quadruplex
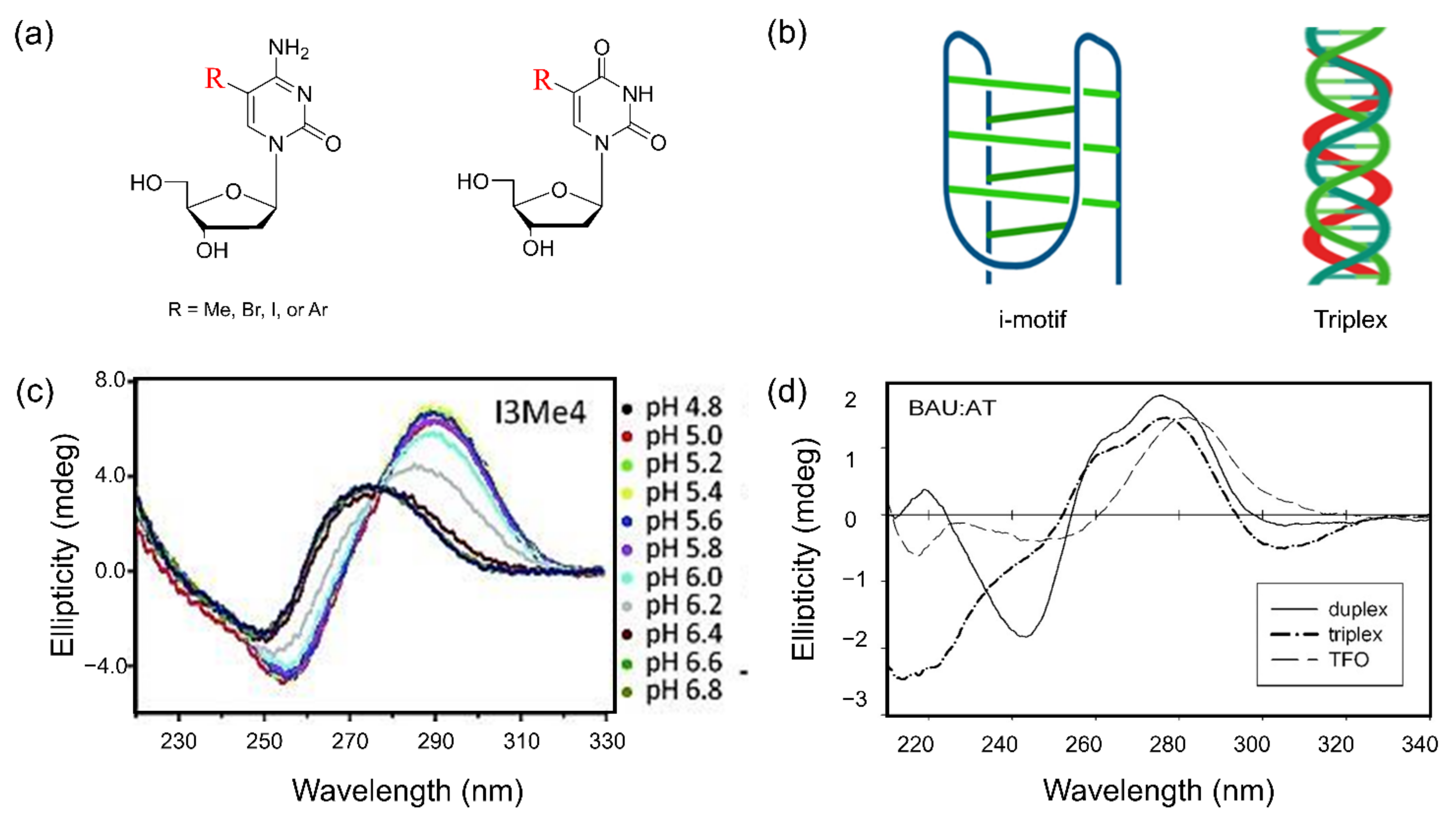
5.3. i-Motif Structure
5.4. Triple Helix
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Freier, S.M.; Altmann, K.-H. The ups and downs of nucleic acid duplex stability: Structure-stability studies on chemically-modified DNA:RNA duplexes. Nucleic Acids Res. 1997, 25, 4429–4443. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, V.B.; Taylor, A.I.; Cozens, C.; Abramov, M.; Renders, M.; Zhang, S.; Chaput, J.C.; Wengel, J.; Peak-Chew, S.-Y.; McLaughlin, S.H.; et al. Synthetic Genetic Polymers Capable of Heredity and Evolution. Science 2012, 336, 341. [Google Scholar] [CrossRef] [Green Version]
- Ni, S.; Yao, H.; Wang, L.; Lu, J.; Jiang, F.; Lu, A.; Zhang, G. Chemical Modifications of Nucleic Acid Aptamers for Therapeutic Purposes. Int. J. Mol. Sci. 2017, 18, 1683. [Google Scholar] [CrossRef]
- Goodchild, J. Conjugates of oligonucleotides and modified oligonucleotides: A review of their synthesis and properties. Bioconjugate Chem. 1990, 1, 165–187. [Google Scholar] [CrossRef] [PubMed]
- Doluca, O.; Withers, J.M.; Filichev, V.V. Molecular engineering of guanine-rich sequences: Z-DNA, DNA triplexes, and G-quadruplexes. Chem. Rev. 2013, 113, 3044–3083. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Hudson, R.H.E. Base-modified fluorescent purine nucleosides and nucleotides for use in oligonucleotide probes. J. Photochem. Photobiol. C 2018, 36, 48–73. [Google Scholar] [CrossRef]
- McKenzie, L.K.; El-Khoury, R.; Thorpe, J.D.; Damha, M.J.; Hollenstein, M. Recent progress in non-native nucleic acid modifications. Chem. Soc. Rev. 2021, 50, 5126–5164. [Google Scholar] [CrossRef]
- Cline, R.E.; Fink, R.M.; Fink, K. Synthesis of 5-Substituted Pyrimidines via Formaldehyde Addition1. J. Am. Chem. Soc. 1959, 81, 2521–2527. [Google Scholar] [CrossRef]
- Frisch, D.M.; Visser, D.W. 5-Bromodeoxycytidine and 5-Chlorodeoxycytidine1. J. Am. Chem. Soc. 1959, 81, 1756–1758. [Google Scholar] [CrossRef]
- Uesugi, S.; Ikehara, M. Carbon-13 magnetic resonance spectra of 8-substituted purine nucleosides. Characteristic shifts for the syn conformation. J. Am. Chem. Soc. 1977, 99, 3250–3253. [Google Scholar] [CrossRef]
- Vongsutilers, V.; Phillips, D.J.; Train, B.C.; McKelvey, G.R.; Thomsen, N.M.; Shaughnessy, K.H.; Lewis, J.P.; Gannett, P.M. The conformational effect of para-substituted C8-arylguanine adducts on the B/Z-DNA equilibrium. Biophys. Chem. 2011, 154, 41–48. [Google Scholar] [CrossRef]
- Watson, J.D.; Crick, F.H.C. Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid. Nature 1953, 171, 737–738. [Google Scholar] [CrossRef] [PubMed]
- Rich, A.; Zhang, S. Timeline: Z-DNA: The long road to biological function. Nat. Rev. Genet. 2003, 4, 566–572. [Google Scholar] [CrossRef]
- Wang, A.H.; Quigley, G.J.; Kolpak, F.J.; Crawford, J.L.; van Boom, J.H.; van der Marel, G.; Rich, A. Molecular structure of a left-handed double helical DNA fragment at atomic resolution. Nature 1979, 282, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.C.; Lowenhaupt, K.; Rich, A.; Kim, Y.G.; Kim, K.K. Crystal structure of a junction between B-DNA and Z-DNA reveals two extruded bases. Nature 2005, 437, 1183–1186. [Google Scholar] [CrossRef]
- Gros, J.; Rosu, F.; Amrane, S.; De Cian, A.; Gabelica, V.; Lacroix, L.; Mergny, J.L. Guanines are a quartet’s best friend: Impact of base substitutions on the kinetics and stability of tetramolecular quadruplexes. Nucleic Acids Res. 2007, 35, 3064–3075. [Google Scholar] [CrossRef]
- Davis, J.T. G-quartets 40 years later: From 5′-GMP to molecular biology and supramolecular chemistry. Angew. Chem. Int. Ed. Engl. 2004, 43, 668–698. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.Y.; Wang, X.N.; Cheng, S.Q.; Su, X.X.; Ou, T.M. Developing Novel G-Quadruplex Ligands: From Interaction with Nucleic Acids to Interfering with Nucleic Acid(-)Protein Interaction. Molecules 2019, 24, 396. [Google Scholar] [CrossRef] [Green Version]
- Wittig, B.; Wölfl, S.; Dorbic, T.; Vahrson, W.; Rich, A. Transcription of human c-myc in permeabilized nuclei is associated with formation of Z-DNA in three discrete regions of the gene. EMBO J. 1992, 11, 4653–4663. [Google Scholar] [CrossRef]
- Maruyama, A.; Mimura, J.; Harada, N.; Itoh, K. Nrf2 activation is associated with Z-DNA formation in the human HO-1 promoter. Nucleic Acids Res. 2013, 41, 5223–5234. [Google Scholar] [CrossRef] [Green Version]
- Manderville, R.A.; Wetmore, S.D. C-Linked 8-aryl guanine nucleobase adducts: Biological outcomes and utility as fluorescent probes. Chem. Sci. 2016, 7, 3482–3493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xodo, L.E.; Manzini, G.; Quadrifoglio, F.; van der Marel, G.A.; van Boom, J.H. Effect pf 5-methylcytosine on the stability of triple-stranded DNA—a thermodynamic study. Nucleic Acids Res. 1991, 19, 5625–5631. [Google Scholar] [CrossRef] [PubMed]
- Ontiveros, R.J.; Stoute, J.; Liu, K.F. The chemical diversity of RNA modifications. Biochem. J. 2019, 476, 1227–1245. [Google Scholar] [CrossRef]
- Tateishi-Karimata, H.; Sugimoto, N. Chemical biology of non-canonical structures of nucleic acids for therapeutic applications. Chem. Comm. 2020, 56, 2379–2390. [Google Scholar] [CrossRef] [PubMed]
- Ngassa, F.N.; Dekorver, K.A.; Melistas, T.S.; Yeh, E.A.; Lakshman, M.K. Pd-Xantphos-catalyzed direct arylation of nucleosides. Org. Lett. 2006, 8, 4613–4616. [Google Scholar] [CrossRef] [PubMed]
- Bhilare, S.; Gayakhe, V.; Ardhapure, A.V.; Sanghvi, Y.S.; Schulzke, C.; Borozdina, Y.; Kapdi, A.R. Novel water-soluble phosphatriazenes: Versatile ligands for Suzuki-Miyaura, Sonogashira and Heck reactions of nucleosides. RSC Adv. 2016, 6, 83820–83830. [Google Scholar] [CrossRef]
- Thomsen, N.M.; Vongsutilers, V.; Gannett, P.M. The synthesis of C8-aryl purines, nucleosides and phosphoramidites. Crit. Rev. Eukaryot. Gene Expr. 2011, 21, 155–176. [Google Scholar] [CrossRef]
- Hingerty, B.; Broyde, S. Conformation of the deoxydinucleoside monophosphate dCpdG modified at carbon 8 of guanine with 2-(acetylamino)fluorene. Biochemistry 1982, 21, 3243–3252. [Google Scholar] [CrossRef] [PubMed]
- Araki, M.; Maeda, M.; Kawazoe, Y. Chemical alteration of nucleic acids and their components—XIII. Tetrahedron 1976, 32, 337–340. [Google Scholar] [CrossRef]
- Humphreys, W.G.; Kadlubar, F.F.; Guengerich, F.P. Mechanism of C8 alkylation of guanine residues by activated arylamines: Evidence for initial adduct formation at the N7 position. Proc. Natl. Acad. Sci. USA 1992, 89, 8278–8282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masiero, S.; Trotta, R.; Pieraccini, S.; De Tito, S.; Perone, R.; Randazzo, A.; Spada, G.P. A non-empirical chromophoric interpretation of CD spectra of DNA G-quadruplex structures. Org. Biomol. Chem. 2010, 8, 2683–2692. [Google Scholar] [CrossRef]
- Gannett, P.M.; Heavner, S.; Daft, J.R.; Shaughnessy, K.H.; Epperson, J.D.; Greenbaum, N.L. Synthesis, properties, and NMR studies of a C8-phenylguanine modified oligonucleotide that preferentially adopts the Z DNA conformation. Chem. Res. Toxicol. 2003, 16, 1385–1394. [Google Scholar] [CrossRef]
- Rice, J.M.; Dudek, G.O. Mass spectra of nucleic acid derivatives. II. Guanine, adenine, and related compounds. J. Am. Chem. Soc. 1967, 89, 2719–2725. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.W.; Robins, R.K. Purine Nucleosides. III. Methylation Studies of Certain Naturally Occurring Purine Nucleosides. J. Am. Chem. Soc. 1963, 85, 193–201. [Google Scholar] [CrossRef]
- Rogers, K.J.; Pegg, A.E. Formation of O6-methylguanine by alkylation of rat liver, colon, and kidney DNA following administration of 1,2-dimethylhydrazine. Cancer Res. 1977, 37, 4082–4087. [Google Scholar] [PubMed]
- Araie, Y.; Ohtsuki, S.; Park, S.; Nagaoka, M.; Umemura, K.; Sugiyama, H.; Kusamori, K.; Takahashi, Y.; Takakura, Y.; Nishikawa, M. Combined use of chemically modified nucleobases and nanostructured DNA for enhanced immunostimulatory activity of CpG oligodeoxynucleotide. Bioorgan. Med. Chem. 2021, 29, 115864. [Google Scholar] [CrossRef] [PubMed]
- Virgilio, A.; Esposito, V.; Citarella, G.; Pepe, A.; Mayol, L.; Galeone, A. The insertion of two 8-methyl-2′-deoxyguanosine residues in tetramolecular quadruplex structures: Trying to orientate the strands. Nucleic Acids Res. 2012, 40, 461–475. [Google Scholar] [CrossRef] [Green Version]
- Thomson, J.M.; Lamont, I.L. Nucleoside Analogues as Antibacterial Agents. Front. Microbiol. 2019, 10, 952. [Google Scholar] [CrossRef] [Green Version]
- Maeda, M.; Nushi, K.; Kawazoe, Y. Studies on chemical alterations of nucleic acids and their components—VII. Tetrahedron 1974, 30, 2677–2682. [Google Scholar] [CrossRef]
- Kohda, K.; Tsunomoto, H.; Minoura, Y.; Tanabe, K.; Shibutani, S. Synthesis, miscoding specificity, and thermodynamic stability of oligodeoxynucleotide containing 8-methyl-2′-deoxyguanosine. Chem. Res. Toxicol. 1996, 9, 1278–1284. [Google Scholar] [CrossRef]
- Martinez, G.R.; Gasparutto, D.; Ravanat, J.L.; Cadet, J.; Medeiros, M.H.; Di Mascio, P. Identification of the main oxidation products of 8-methoxy-2′-deoxyguanosine by singlet molecular oxygen. Free Radic. Biol. Med. 2005, 38, 1491–1500. [Google Scholar] [CrossRef]
- Crean, C.; Geacintov, N.E.; Shafirovich, V. Methylation of 2′-deoxyguanosine by a free radical mechanism. J. Phys. Chem. B 2009, 113, 12773–12781. [Google Scholar] [CrossRef] [Green Version]
- Zady, M.F.; Wong, J.L. Kinetics and mechanism of carbon-8 methylation of purine bases and nucleosides by methyl radical. J. Am. Chem. Soc. 1977, 99, 5096–5101. [Google Scholar] [CrossRef] [PubMed]
- Elskens, J.; Manicardi, A.; Costi, V.; Madder, A.; Corradini, R. Synthesis and Improved Cross-Linking Properties of C5-Modified Furan Bearing PNAs. Molecules 2017, 22, 2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, H.-Y.; Su, L.-Y.; Bai, S.-X.; Li, J.-P.; Feng, X.-L.; Guo, H.-M. Synthesis of C8-alkyl-substituted purine analogues by direct alkylation of 8- H purines with tetrahydrofuran catalyzed by CoCl2·6H2O. Chin. Chem. Lett. 2017, 28, 105–108. [Google Scholar] [CrossRef]
- Yamazaki, A.; Kumashiro, I.; Takenishi, T. Synthesis of guanosine and its derivatives from 5-amino-1-beta-D-ribofuranosyl-4-imidazolecarboxamide. II. Ring closure with sodium methylxanthate. J. Org. Chem. 1967, 32, 3032–3038. [Google Scholar] [CrossRef]
- Pfleiderer, W.; Shanshal, M.; Eistetter, K. Nucleosides, V. Synthesis and structure of 8-methyl- and 8-(trifluoromethyl) guanine nucleosides. Chem. Ber. 1972, 105, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Ikehara, M.; Limn, W.; Fukui, T. Studies of nucleosides and nucleotides. LXXXI. Synthesis and characterization of 8-methyladenosine. Chem. Pharm. Bull. 1977, 25, 2702–2707. [Google Scholar] [CrossRef] [Green Version]
- Hirota, K.; Kitade, Y.; Kanbe, Y.; Maki, Y. Convenient method for the synthesis of C-alkylated purine nucleosides: Palladium-catalyzed cross-coupling reaction of halogenopurine nucleosides with trialkylaluminums. J. Org. Chem. 1992, 57, 5268–5270. [Google Scholar] [CrossRef]
- Henderson, J.P.; Byun, J.; Williams, M.V.; McCormick, M.L.; Parks, W.C.; Ridnour, L.A.; Heinecke, J.W. Bromination of deoxycytidine by eosinophil peroxidase: A mechanism for mutagenesis by oxidative damage of nucleotide precursors. Proc. Natl. Acad. Sci. USA 2001, 98, 1631–1636. [Google Scholar] [CrossRef]
- Volpini, R.; Costanzi, S.; Lambertucci, C.; Vittori, S.; Klotz, K.; Lorenzen, A.; Cristalli, G. Introduction of alkynyl chains on C-8 of adenosine led to very selective antagonists of the A(3) adenosine receptor. Bioorgan. Med. Chem. Lett. 2001, 11, 1931–1934. [Google Scholar] [CrossRef]
- El Safadi, Y.; Marquet, R.; Aubertin, A.M.; Vivet-Boudou, V. Synthesis and primary evaluation of novel HIV-1 inhibitors. Nucleosides Nucleotides Nucleic Acids 2007, 26, 1161–1165. [Google Scholar] [CrossRef]
- Asahi, T.; Kondo, H.; Masuda, M.; Nishino, H.; Aratani, Y.; Naito, Y.; Yoshikawa, T.; Hisaka, S.; Kato, Y.; Osawa, T. Chemical and immunochemical detection of 8-halogenated deoxyguanosines at early stage inflammation. J. Biol. Chem. 2010, 285, 9282–9291. [Google Scholar] [CrossRef] [Green Version]
- Budovskii, E.I.; Shibaev, V.N.; Spiridonova, S.M.; Kochetkov, N.K. Synthesis of 8-bromoadenosine 5′-(α-D-glucopyranosyl diphosphate). Bull. Acad. Sci. USSR Div. Chem. Sci. 1971, 20, 1183–1185. [Google Scholar] [CrossRef]
- Münzel, M.; Szeibert, C.; Glas, A.F.; Globisch, D.; Carell, T. Discovery and Synthesis of New UV-Induced Intrastrand C(4−8)G and G(8−4)C Photolesions. J. Am. Chem. Soc. 2011, 133, 5186–5189. [Google Scholar] [CrossRef]
- Ikehara, M.; Kaneko, M. Studies of nucleosides and nucleotides—XLI. Tetrahedron 1970, 26, 4251–4259. [Google Scholar] [CrossRef]
- Maity, J.; Stromberg, R. An efficient and facile methodology for bromination of pyrimidine and purine nucleosides with sodium monobromoisocyanurate (SMBI). Molecules 2013, 18, 12740–12750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asahi, T.; Nakamura, Y.; Kato, Y.; Osawa, T. Specific role of taurine in the 8-brominated′-deoxyguanosine formation. Arch. Biochem. Biophys. 2015, 586, 45–50. [Google Scholar] [CrossRef]
- Varizhuk, A.M.; Zatsepin, T.S.; Golovin, A.V.; Belyaev, E.S.; Kostyukevich, Y.I.; Dedkov, V.G.; Shipulin, G.A.; Shpakovski, G.V.; Aralov, A.V. Synthesis of oligonucleotides containing novel G-clamp analogue with C8-tethered group in phenoxazine ring: Implication to qPCR detection of the low-copy Kemerovo virus dsRNA. Bioorgan. Med. Chem. 2017, 25, 3597–3605. [Google Scholar] [CrossRef] [PubMed]
- Gillet, L.C.J.; Schärer, O.D. Preparation of C8-Amine and Acetylamine Adducts of 2′-Deoxyguanosine Suitably Protected for DNA Synthesis. Org. Lett. 2002, 4, 4205–4208. [Google Scholar] [CrossRef] [PubMed]
- Seregin, I.V.; Gevorgyan, V. Direct transition metal-catalyzed functionalization of heteroaromatic compounds. Chem. Soc. Rev. 2007, 36, 1173–1193. [Google Scholar] [CrossRef] [PubMed]
- Čerňa, I.; Pohl, R.; Hocek, M. The first direct C–H arylation of purinenucleosides. Chem. Comm. 2007, 4729–4730. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Jackson, M.J.; Smith, L.A.; Pearce, R.K.; Nakamura, J.; Kase, H.; Kuwana, Y.; Jenner, P. Adenosine A2A antagonist: A novel antiparkinsonian agent that does not provoke dyskinesia in parkinsonian monkeys. Ann. Neurol. 1998, 43, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Gudmundsson, K.S.; Daluge, S.M.; Condreay, L.D.; Johnson, L.C. Synthesis of novel 8-substituted carbocyclic analogs of 2′,3′-dideoxyadenosine with activity against hepatitis B virus. Nucleosides Nucleotides Nucleic Acids 2002, 21, 891–901. [Google Scholar] [CrossRef]
- Carothers, A.M.; Yuan, W.; Hingerty, B.E.; Broyde, S.; Grunberger, D.; Snyderwine, E.G. Mutation and repair induced by the carcinogen 2-(hydroxyamino)-1-methyl-6-phenylimidazo[4,5-b]pyridine (N-OH-PhIP) in the dihydrofolate reductase gene of Chinese hamster ovary cells and conformational modeling of the dG-C8-PhIP adduct in DNA. Chem. Res. Toxicol. 1994, 7, 209–218. [Google Scholar] [CrossRef] [PubMed]
- De Leeuw, W.J.; van Puijenbroek, M.; Merx, R.; Wijnen, J.T.; Bröcker-Vriends, A.H.; Tops, C.; Vasen, H.; Cornelisse, C.J.; Morreau, H. Bias in detection of instability of the (C)8 mononucleotide repeat of MSH6 in tumours from HNPCC patients. Oncogene 2001, 20, 6241–6244. [Google Scholar] [CrossRef]
- Kuska, M.S.; Witham, A.A.; Sproviero, M.; Manderville, R.A.; Majdi Yazdi, M.; Sharma, P.; Wetmore, S.D. Structural influence of C8-phenoxy-guanine in the NarI recognition DNA sequence. Chem. Res. Toxicol. 2013, 26, 1397–1408. [Google Scholar] [CrossRef] [PubMed]
- Agrofoglio, L.A.; Gillaizeau, I.; Saito, Y. Palladium-assisted routes to nucleosides. Chem. Rev. 2003, 103, 1875–1916. [Google Scholar] [CrossRef] [PubMed]
- Western, E.C.; Shaughnessy, K.H. Inhibitory Effects of the Guanine Moiety on Suzuki Couplings of Unprotected Halonucleosides in Aqueous Media. J. Org. Chem. 2005, 70, 6378–6388. [Google Scholar] [CrossRef]
- Vaňková, B.; Krchňák, V.; Soural, M.; Hlaváč, J. Direct C–H Arylation of Purine on Solid Phase and Its Use for Chemical Libraries Synthesis. ACS Comb. Sci. 2011, 13, 496–500. [Google Scholar] [CrossRef]
- Abdoli, M.; Mirjafary, Z.; Saeidian, H.; Kakanejadifard, A. New developments in direct functionalization of C–H and N–H bonds of purine bases via metal catalyzed cross-coupling reactions. RSC Adv. 2015, 5, 44371–44389. [Google Scholar] [CrossRef]
- Omumi, A.; Beach, D.G.; Baker, M.; Gabryelski, W.; Manderville, R.A. Postsynthetic Guanine Arylation of DNA by Suzuki−Miyaura Cross-Coupling. J. Am. Chem. Soc. 2011, 133, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, P.H.N.; Furtado, C.; Repolês, B.M.; Ribeiro, G.A.; Mendes, I.C.; Peloso, E.F.; Gadelha, F.R.; Macedo, A.M.; Franco, G.R.; Pena, S.D.J.; et al. Oxidative Stress and DNA Lesions: The Role of 8-Oxoguanine Lesions in Trypanosoma cruzi Cell Viability. PLoS Neglected Trop. Dis. 2013, 7, e2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tchou, J.; Bodepudi, V.; Shibutani, S.; Antoshechkin, I.; Miller, J.; Grollman, A.P.; Johnson, F. Substrate specificity of Fpg protein. Recognition and cleavage of oxidatively damaged DNA. J. Biol. Chem. 1994, 269, 15318–15324. [Google Scholar] [CrossRef]
- Dai, Q.; Xu, D.; Lim, K.; Harvey, R.G. Efficient Syntheses of C8-Aryl Adducts of Adenine and Guanine Formed by Reaction of Radical Cation Metabolites of Carcinogenic Polycyclic Aromatic Hydrocarbons with DNA. J. Org. Chem. 2007, 72, 4856–4863. [Google Scholar] [CrossRef]
- Vongsutilers, V.; Daft, J.R.; Shaughnessy, K.H.; Gannett, P.M. A general synthesis of C8-arylpurine phosphoramidites. Molecules 2009, 14, 3339–3352. [Google Scholar] [CrossRef] [Green Version]
- Storr, T.E.; Firth, A.G.; Wilson, K.; Darley, K.; Baumann, C.G.; Fairlamb, I.J.S. Site-selective direct arylation of unprotected adenine nucleosides mediated by palladium and copper: Insights into the reaction mechanism. Tetrahedron 2008, 64, 6125–6137. [Google Scholar] [CrossRef]
- Tu, C.; Keane, C.; Eaton, B.E. Palladium Catalysis in the Synthesis of 8-Position modified Adenosine, 2′-Deoxyadenosine and Guanosine. Nucleosides Nucleotides Nucleic Acids 1995, 14, 1631–1638. [Google Scholar] [CrossRef]
- Xu, Y.; Ikeda, R.; Sugiyama, H. 8-Methylguanosine: A powerful Z-DNA stabilizer. J. Am. Chem. Soc. 2003, 125, 13519–13524. [Google Scholar] [CrossRef]
- Bhanjadeo, M.M.; Subudhi, U. Praseodymium promotes B–Z transition in self-assembled DNA nanostructures. RSC Adv. 2019, 9, 4616–4620. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Zhao, A.; Ren, J.; Qu, X. Lighting up left-handed Z-DNA: Photoluminescent carbon dots induce DNA B to Z transition and perform DNA logic operations. Nucleic Acids Res. 2013, 41, 7987–7996. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Tian, T.; Yu, J.; Weng, X.; Liu, Y.; Zhou, X. Formation of sequence-independent Z-DNA induced by a ruthenium complex at low salt concentrations. Angew. Chem. Int. Ed. Engl. 2011, 50, 11962–11967. [Google Scholar] [CrossRef]
- Kim, S.H.; Lim, S.H.; Lee, A.R.; Kwon, D.H.; Song, H.K.; Lee, J.H.; Cho, M.; Johner, A.; Lee, N.K.; Hong, S.C. Unveiling the pathway to Z-DNA in the protein-induced B-Z transition. Nucleic Acids Res. 2018, 46, 4129–4137. [Google Scholar] [CrossRef] [Green Version]
- Herbert, A.; Schade, M.; Lowenhaupt, K.; Alfken, J.; Schwartz, T.; Shlyakhtenko, L.S.; Lyubchenko, Y.L.; Rich, A. The Zα domain from human ADAR1 binds to the Z-DNA conformer of many different sequences. Nucleic Acids Res. 1998, 26, 3486–3493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiyama, H.; Kawai, K.; Matsunaga, A.; Fujimoto, K.; Saito, I.; Robinson, H.; Wang, A.H. Synthesis, structure and thermodynamic properties of 8-methylguanine-containing oligonucleotides: Z-DNA under physiological salt conditions. Nucleic Acids Res. 1996, 24, 1272–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramaniyam, T.; Ishizuka, T.; Xu, Y. Stability and properties of Z-DNA containing artificial nucleobase 2′-O-methyl-8-methyl guanosine. Bioorgan. Med. Chem. 2019, 27, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniyam, T.; Ishizuka, T.; Xiao, C.D.; Bao, H.L.; Xu, Y. Observation of Z-DNA Structure via the Synthesis of Oligonucleotide DNA Containing 8-Trifluoromethyl-2-Deoxyguanosine. Molecules 2018, 23, 2572. [Google Scholar] [CrossRef] [Green Version]
- Bao, H.-L.; Masuzawa, T.; Oyoshi, T.; Xu, Y. Oligonucleotides DNA containing 8-trifluoromethyl-2′-deoxyguanosine for observing Z-DNA structure. Nucleic Acids Res. 2020, 48, 7041–7051. [Google Scholar] [CrossRef]
- Train, B.C.; Bilgesü, S.A.; Despeaux, E.C.; Vongsutilers, V.; Gannett, P.M. Single C8-Arylguanine modifications render oligonucleotides in the Z-DNA conformation under physiological conditions. Chem. Res. Toxicol. 2014, 27, 1176–1186. [Google Scholar] [CrossRef]
- Chen, F.Y.; Park, S.; Otomo, H.; Sakashita, S.; Sugiyama, H. Investigation of B-Z transitions with DNA oligonucleotides containing 8-methylguanine. Artif. DNA PNA XNA 2014, 5, e28226. [Google Scholar] [CrossRef] [Green Version]
- Moller, A.; Nordheim, A.; Kozlowski, S.A.; Patel, D.; Rich, A. Bromination stabilizes poly(dG-dC) in the Z-DNA form under low-salt conditions. Biochemistry 1984, 23, 54–62. [Google Scholar] [CrossRef]
- Lafer, E.M.; Moller, A.; Nordheim, A.; Stollar, B.D.; Rich, A. Antibodies specific for left-handed Z-DNA. Proc. Natl. Acad. Sci. USA 1981, 78, 3546–3550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, W.S.; Hardin, C.C.; Tinoco, I., Jr.; Rao, S.N.; Pearlman, D.A.; Kollman, P.A. Effects of nucleotide bromination on the stabilities of Z-RNA and Z-DNA: A molecular mechanics/thermodynamic perturbation study. Biopolymers 1989, 28, 1939–1957. [Google Scholar] [CrossRef]
- Suárez-Marina, I.; Abul-Haija, Y.M.; Turk-MacLeod, R.; Gromski, P.S.; Cooper, G.J.T.; Olivé, A.O.; Colón-Santos, S.; Cronin, L. Integrated synthesis of nucleotide and nucleosides influenced by amino acids. Commun. Chem. 2019, 2, 28. [Google Scholar] [CrossRef]
- Rayala, R.; Wnuk, S.F. Bromination at C-5 of Pyrimidine and C-8 of Purine Nucleosides with 1,3-Dibromo-5,5-dimethylhydantoin. Tetrahedron Lett. 2012, 53, 3333–3336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, R.; Ellis, S.; Hingerty, B.E.; Broyde, S. Effect of ring size on conformations of aromatic amine-DNA adducts: The aniline-C8 guanine adduct resides in the B-DNA major groove. Chem. Res. Toxicol. 1998, 11, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Turesky, R.J.; Markovic, J. DNA adduct formation of the food carcinogen 2-amino-3-methylimidazo[4,5- f]quinoline at the C-8 and N2 atoms of guanine. Chem. Res. Toxicol. 1994, 7, 752–761. [Google Scholar] [CrossRef]
- Van Houte, L.P.; Westra, J.G.; Retèl, J.; van Grondelle, R. A spectroscopic study of the conformation of poly d(G-C).poly d(G-C) modified with the carcinogenic 2-aminofluorene. Carcinogenesis 1988, 9, 1017–1027. [Google Scholar] [CrossRef]
- Galiègue-Zouitina, S.; Bailleul, B.; Loucheux-Lefebvre, M.H. Guanyl-C8-arylamination of DNA by the ultimate carcinogen of 4-nitroquinoline-1-oxide: A spectrophotometric titration. Anal. Biochem. 1984, 138, 454–457. [Google Scholar] [CrossRef]
- Gu, Z.; Gorin, A.; Hingerty, B.E.; Broyde, S.; Patel, D.J. Solution structures of aminofluorene [AF]-stacked conformers of the syn [AF]-C8-dG adduct positioned opposite dC or dA at a template-primer junction. Biochemistry 1999, 38, 10855–10870. [Google Scholar] [CrossRef]
- Sharma, P.; Manderville, R.A.; Wetmore, S.D. Modeling the Conformational Preference of the Carbon-Bonded Covalent Adduct Formed upon Exposure of 2′-Deoxyguanosine to Ochratoxin, A. Chem. Res. Toxicol. 2013, 26, 803–816. [Google Scholar] [CrossRef]
- Ebert, C.; Simon, N.; Schneider, S.; Carell, T. Structural Insights into the Recognition of N(2) -Aryl- and C8-Aryl DNA Lesions by the Repair Protein XPA/Rad14. Chembiochem 2017, 18, 1379–1382. [Google Scholar] [CrossRef]
- Phan, A.T.; Kuryavyi, V.; Luu, K.N.; Patel, D.J. Structure of two intramolecular G-quadruplexes formed by natural human telomere sequences in K+ solution. Nucleic Acids Res. 2007, 35, 6517–6525. [Google Scholar] [CrossRef] [Green Version]
- Skolakova, P.; Bednarova, K.; Vorlickova, M.; Sagi, J. Quadruplexes of human telomere dG(3)(TTAG(3))(3) sequences containing guanine abasic sites. Biochem. Biophys. Res. Commun. 2010, 399, 203–208. [Google Scholar] [CrossRef]
- Smith, F.W.; Feigon, J. Strand orientation in the DNA quadruplex formed from the Oxytricha telomere repeat oligonucleotide d(G4T4G4) in solution. Biochemistry 1993, 32, 8682–8692. [Google Scholar] [CrossRef]
- Tomasko, M.; Vorlickova, M.; Sagi, J. Substitution of adenine for guanine in the quadruplex-forming human telomere DNA sequence G(3)(T(2)AG(3))(3). Biochimie 2009, 91, 171–179. [Google Scholar] [CrossRef]
- Xu, Y.; Noguchi, Y.; Sugiyama, H. The new models of the human telomere d[AGGG(TTAGGG)3] in K+ solution. Bioorgan. Med. Chem. 2006, 14, 5584–5591. [Google Scholar] [CrossRef] [PubMed]
- Lech, C.J.; Cheow Lim, J.K.; Wen Lim, J.M.; Amrane, S.; Heddi, B.; Phan, A.T. Effects of site-specific guanine C8-modifications on an intramolecular DNA G-quadruplex. Biophys. J. 2011, 101, 1987–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagi, J. G-quadruplexes incorporating modified constituents: A review. J. Biomol. Struct. Dyn. 2014, 32, 477–511. [Google Scholar] [CrossRef] [PubMed]
- Sannohe, Y.; Sato, K.; Matsugami, A.; Shinohara, K.; Mashimo, T.; Katahira, M.; Sugiyama, H. Orientation of ends of G-quadruplex structure investigated with end-extended oligonucleotides. Nucleic. Acids. Symp. Ser. 2008, 171–172. [Google Scholar] [CrossRef] [Green Version]
- Virgilio, A.; Esposito, V.; Randazzo, A.; Mayol, L.; Galeone, A. Effects of 8-methyl-2′-deoxyadenosine incorporation into quadruplex forming oligodeoxyribonucleotides. Bioorgan. Med. Chem. 2005, 13, 1037–1044. [Google Scholar] [CrossRef]
- Virgilio, A.; Esposito, V.; Randazzo, A.; Mayol, L.; Galeone, A. 8-methyl-2′-deoxyguanosine incorporation into parallel DNA quadruplex structures. Nucleic Acids Res. 2005, 33, 6188–6195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Sugiyama, H. Formation of the G-quadruplex and i-motif structures in retinoblastoma susceptibility genes (Rb). Nucleic Acids Res. 2006, 34, 949–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumas, A.; Luedtke, N.W. Cation-Mediated Energy Transfer in G-Quadruplexes Revealed by an Internal Fluorescent Probe. J. Am. Chem. Soc. 2010, 132, 18004–18007. [Google Scholar] [CrossRef] [PubMed]
- Esposito, V.; Randazzo, A.; Piccialli, G.; Petraccone, L.; Giancola, C.; Mayol, L. Effects of an 8-bromodeoxyguanosine incorporation on the parallel quadruplex structure [d(TGGGT)]4. Org. Biomol. Chem. 2004, 2, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Dias, E.; Battiste, J.L.; Williamson, J.R. Chemical Probe for Glycosidic Conformation in Telomeric DNAs. J. Am. Chem. Soc. 1994, 116, 4479–4480. [Google Scholar] [CrossRef]
- Mashimo, T.; Sannohe, Y.; Yagi, H.; Sugiyama, H. Folding pathways of hybrid-1 and hybrid-2 G-quadruplex structures. Nucleic Acids Symp. Ser. 2008, 409–410. [Google Scholar] [CrossRef] [Green Version]
- Matsugami, A.; Xu, Y.; Noguchi, Y.; Sugiyama, H.; Katahira, M. Structure of a human telomeric DNA sequence stabilized by 8-bromoguanosine substitutions, as determined by NMR in a K+ solution. FEBS J. 2007, 274, 3545–3556. [Google Scholar] [CrossRef]
- Matsugami, A.; Tsuchibayashi, H.; Xu, Y.; Noguchi, Y.; Sugiyama, H.; Katahira, M. The new models of the human telomere DNA in K+ solution revealed by NMR analysis assisted by the incorporation of 8-bromoguanines. Nucleic Acids Symp. Ser. 2006, 45–46. [Google Scholar] [CrossRef] [Green Version]
- Gubala, V.; Betancourt, J.E.; Rivera, J.M. Expanding the Hoogsteen Edge of 2′-Deoxyguanosine: Consequences for G-Quadruplex Formation. Org. Lett. 2004, 6, 4735–4738. [Google Scholar] [CrossRef] [Green Version]
- Szalai, V.A.; Singer, M.J.; Thorp, H.H. Site-specific probing of oxidative reactivity and telomerase function using 7,8-dihydro-8-oxoguanine in telomeric DNA. J. Am. Chem. Soc. 2002, 124, 1625–1631. [Google Scholar] [CrossRef]
- López de la Osa, J.; González, C.; Gargallo, R.; Rueda, M.; Cubero, E.; Orozco, M.; Aviñó, A.; Eritja, R. Destabilization of quadruplex DNA by 8-aminoguanine. Chembiochem 2006, 7, 46–48. [Google Scholar] [CrossRef]
- Dumas, A.; Luedtke, N.W. Highly fluorescent guanosine mimics for folding and energy transfer studies. Nucleic Acids Res. 2011, 39, 6825–6834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Gaied, N.; Glasser, N.; Ramalanjaona, N.; Beltz, H.; Wolff, P.; Marquet, R.; Burger, A.; Mély, Y. 8-vinyl-deoxyadenosine, an alternative fluorescent nucleoside analog to 2′-deoxyribosyl-2-aminopurine with improved properties. Nucleic Acids Res. 2005, 33, 1031–1039. [Google Scholar] [CrossRef] [Green Version]
- Ogasawara, S.; Maeda, M. Photo-controllable aptamer. Nucleic Acids Symp. Ser. 2009, 195–196. [Google Scholar] [CrossRef]
- Aschenbrenner, D.; Baumann, F.; Milles, L.F.; Pippig, D.A.; Gaub, H.E. C-5 Propynyl Modifications Enhance the Mechanical Stability of DNA. ChemPhysChem 2015, 16, 2085–2090. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Pastor, W.A.; Shen, Y.; Tahiliani, M.; Liu, D.R.; Rao, A. The Behaviour of 5-Hydroxymethylcytosine in Bisulfite Sequencing. PLoS ONE 2010, 5, e8888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, D.J. 2.13—Pyrimidines and their Benzo Derivatives. In Comprehensive Heterocyclic Chemistry; Katritzky, A.R., Rees, C.W., Eds.; Pergamon: Oxford, UK, 1984; pp. 57–155. [Google Scholar]
- Hayakawa, H.; Tanaka, H.; Obi, K.; Itoh, M.; Miyasaka, T. A simple and general entry to 5-substituted uridines based on the regioselective lithiation controlled by a protecting group in the sugar moiety. Tetrahedron Lett. 1987, 28, 87–90. [Google Scholar] [CrossRef]
- Tahiliani, M.; Koh, K.P.; Shen, Y.H.; Pastor, W.A.; Bandukwala, H.; Brudno, Y.; Agarwal, S.; Iyer, L.M.; Liu, D.R.; Aravind, L.; et al. Conversion of 5-Methylcytosine to 5-Hydroxymethylcytosine in Mammalian DNA by MLL Partner TET1. Science 2009, 324, 930–935. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.; Chan, M.F.; Tomigahara, Y.; Tsai, Y.C.; Gonzales, F.A.; Li, E.; Laird, P.W.; Jones, P.A. Cooperativity between DNA methyltransferases in the maintenance methylation of repetitive elements. Mol. Cell. Biol. 2002, 22, 480–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagase, H.; Ghosh, S. Epigenetics: Differential DNA methylation in mammalian somatic tissues. FEBS J. 2008, 275, 1617–1623. [Google Scholar] [CrossRef]
- Panning, B.; Jaenisch, R. RNA and the Epigenetic Regulation of X Chromosome Inactivation. Cell 1998, 93, 305–308. [Google Scholar] [CrossRef] [Green Version]
- Amort, T.; Rieder, D.; Wille, A.; Khokhlova-Cubberley, D.; Riml, C.; Trixl, L.; Jia, X.-Y.; Micura, R.; Lusser, A. Distinct 5-methylcytosine profiles in poly(A) RNA from mouse embryonic stem cells and brain. Genome Biol. 2017, 18, 1. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, K.; Kishi, S.; Sakamoto, T. Geometric effect on the photocrosslinking reaction between 3-cyanovinylcarbazole nucleoside and pyrimidine base in DNA/RNA heteroduplex. Photochem. Photobiol. 2013, 89, 1095–1099. [Google Scholar] [CrossRef]
- O’Brown, Z.K.; Boulias, K.; Wang, J.; Wang, S.Y.; O’Brown, N.M.; Hao, Z.; Shibuya, H.; Fady, P.E.; Shi, Y.; He, C.; et al. Sources of artifact in measurements of 6mA and 4mC abundance in eukaryotic genomic DNA. BMC Genom. 2019, 20, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, V.; Chitranshi, N.; Agarwal, A.K. Significance and biological importance of pyrimidine in the microbial world. Int. J. Med. Chem. 2014, 2014, 202784. [Google Scholar] [CrossRef] [Green Version]
- Wescoe, Z.L.; Schreiber, J.; Akeson, M. Nanopores Discriminate among Five C5-Cytosine Variants in DNA. J. Am. Chem. Soc. 2014, 136, 16582–16587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergstrom, D.E.; Ogawa, M.K. C-5 substituted pyrimidine nucleosides. 2. Synthesis via olefin coupling to organopalladium intermediates derived from uridine and 2’-deoxyuridine. J. Am. Chem. Soc. 1978, 100, 8106–8112. [Google Scholar] [CrossRef]
- Bradshaw, T.K.; Hutchinson, D.W. 5-Substituted pyrimidine nucleosides and nucleotides. Chem. Soc. Rev. 1977, 6, 43–62. [Google Scholar] [CrossRef]
- Bhat, K.S.; Rao, A.S. Synthesis of Uracil, 6-Methyluracil and Some Dihydrouracils. Org. Prep. Proced. Int. 2009, 15, 303–313. [Google Scholar] [CrossRef]
- Feng, Z.; Hu, W.; Rom, W.N.; Beland, F.A.; Tang, M.S. N-hydroxy-4-aminobiphenyl-DNA binding in human p53 gene: Sequence preference and the effect of C5 cytosine methylation. Biochemistry 2002, 41, 6414–6421. [Google Scholar] [CrossRef]
- Patton, W.; Bacon, V.; Duffield, A.M.; Halpern, B.; Hoyano, Y.; Pereira, W.; Lederberg, J. Chlorination studies. I. The reaction of aqueous hypochlorous acid with cytosine. Biochem. Biophys. Res. Commun. 1972, 48, 880–884. [Google Scholar] [CrossRef]
- Su, D.G.; Taylor, J.S.; Gross, M.L. A new photoproduct of 5-methylcytosine and adenine characterized by high-performance liquid chromatography and mass spectrometry. Chem. Res. Toxicol. 2010, 23, 474–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Ren, L.; Wang, J.; Zheng, G.; Tang, P. Two-step efficient synthesis of 5-methyluridine via two thermostable nucleoside phosphorylase from Aeropyrum pernix. Bioorgan. Med. Chem. Lett. 2012, 22, 2102–2104. [Google Scholar] [CrossRef] [PubMed]
- Sasnauskas, G.; Zagorskaite, E.; Kauneckaite, K.; Tamulaitiene, G.; Siksnys, V. Structure-guided sequence specificity engineering of the modification-dependent restriction endonuclease LpnPI. Nucleic Acids Res. 2015, 43, 6144–6155. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Dyatkina, N.; Prhavc, M.; Williams, C.; Serebryany, V.; Hu, Y.; Huang, Y.; Wan, J.; Wu, X.; Deval, J.; et al. Synthesis and Anti-HCV Activities of 4′-Fluoro-2′-Substituted Uridine Triphosphates and Nucleotide Prodrugs: Discovery of 4′-Fluoro-2′- C-methyluridine 5′-Phosphoramidate Prodrug (AL-335) for the Treatment of Hepatitis C Infection. J. Med. Chem. 2019, 62, 4555–4570. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.K.; Sharma, R.K.; Singh, S.K. Antisense oligonucleotides: Modifications and clinical trials. MedChemComm 2014, 5, 1454–1471. [Google Scholar] [CrossRef]
- Østergaard, M.E.; Kumar, P.; Baral, B.; Guenther, D.C.; Anderson, B.A.; Ytreberg, F.M.; Deobald, L.; Paszczynski, A.J.; Sharma, P.K.; Hrdlicka, P.J. C5-functionalized DNA, LNA, and α-L-LNA: Positional control of polarity-sensitive fluorophores leads to improved SNP-typing. Chemistry 2011, 17, 3157–3165. [Google Scholar] [CrossRef]
- Wolk, S.K.; Mayfield, W.S.; Gelinas, A.D.; Astling, D.; Guillot, J.; Brody, E.N.; Janjic, N.; Gold, L. Modified nucleotides may have enhanced early RNA catalysis. Proc. Natl. Acad. Sci. USA 2020, 117, 8236–8242. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Gianni, L.; Kinsella, T.J.; Klecker, R.W., Jr.; Jenkins, J.; Rowland, J.; Glatstein, E.; Mitchell, J.B.; Collins, J.; Myers, C. Pharmacological evaluation of intravenous delivery of 5-bromodeoxyuridine to patients with brain tumors. Cancer Res. 1984, 44, 1702–1705. [Google Scholar] [PubMed]
- Kofoed, R.H.; Betzer, C.; Lykke-Andersen, S.; Molska, E.; Jensen, P.H. Investigation of RNA Synthesis Using 5-Bromouridine Labelling and Immunoprecipitation. JoVE 2018, e57056. [Google Scholar] [CrossRef] [Green Version]
- Ochoa, S.; Milam, V.T. Modified Nucleic Acids: Expanding the Capabilities of Functional Oligonucleotides. Molecules 2020, 25, 4659. [Google Scholar] [CrossRef]
- Held, H.A.; Roychowdhury, A.; Benner, S.A. C-5 modified nucleosides: Direct insertion of alkynyl-thio functionality in pyrimidines. Nucleosides Nucleotides Nucleic Acids 2003, 22, 391–404. [Google Scholar] [CrossRef]
- Yang, Q.; Wei, T.; He, Y.; Liang, Y.; Zhang, Z.-T. Direct Arylation of 5-Iodouracil and 5-Iodouridine with Heteroarenes and BenzeneviaPhotochemical Reaction. Helv. Chim. Acta 2015, 98, 953–960. [Google Scholar] [CrossRef]
- Hwang, C.H.; Park, J.S.; Won, J.H.; Kim, J.N.; Ryu, E.K. The oxidative iodination of pyrimidine bases and their nucleosides using iodine/dimethylformamide/ m-chloroperbenzoic acid. Arch. Pharmacal Res. 1992, 15, 69–72. [Google Scholar] [CrossRef]
- Itahara, T.; Ide, N. Myeloperoxidase Catalyzed Bromination of Nucleic Acid Bases and Related Compounds. Bull. Chem. Soc. Jpn. 1989, 62, 3750–3751. [Google Scholar] [CrossRef] [Green Version]
- Ito, Y.; Masaki, Y.; Kanamori, T.; Ohkubo, A.; Seio, K.; Sekine, M. Synthesis of 5-[3-(2-aminopyrimidin-4-yl)aminopropyn-1-yl]uracil derivative that recognizes Ade-Thy base pairs in double-stranded DNA. Bioorgan. Med. Chem. Lett. 2016, 26, 194–196. [Google Scholar] [CrossRef]
- Ashwell, M.; Jones, A.S.; Kumar, A.; Sayers, J.R.; Walker, R.T.; Sakuma, T.; De Clercq, E. The synthesis and antiviral properties of ()-5-(2-bromovinyl)-2’-deoxyuridine-related compounds. Tetrahedron 1987, 43, 4601–4608. [Google Scholar] [CrossRef]
- Liang, Y.; Wnuk, S.F. Modification of purine and pyrimidine nucleosides by direct C-H bond activation. Molecules 2015, 20, 4874–4901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Gloudeman, J.; Wnuk, S.F. Palladium-Catalyzed Direct Arylation of 5-Halouracils and 5-Halouracil Nucleosides with Arenes and Heteroarenes Promoted by TBAF. J. Org. Chem. 2014, 79, 4094–4103. [Google Scholar] [CrossRef] [PubMed]
- Ruth, J.L.; Bergstrom, D.E. C-5 substituted pyrimidine nucleosides. 1. Synthesis of C-5 allyl, propyl, and propenyl uracil and cytosine nucleosides via organopalladium intermediates. J. Org. Chem. 1978, 43, 2870–2876. [Google Scholar] [CrossRef]
- Robins, M.J.; Barr, P.J. Nucleic acid related compounds. 39. Efficient conversion of 5-iodo to 5-alkynyl and derived 5-substituted uracil bases and nucleosides. J. Org. Chem. 1983, 48, 1854–1862. [Google Scholar] [CrossRef]
- Western, E.C.; Daft, J.R.; Johnson, E.M.; Gannett, P.M.; Shaughnessy, K.H. Efficient One-Step Suzuki Arylation of Unprotected Halonucleosides, Using Water-Soluble Palladium Catalysts. J. Org. Chem. 2003, 68, 6767–6774. [Google Scholar] [CrossRef]
- Fresneau, N.; Hiebel, M.-A.; Agrofoglio, L.A.; Berteina-Raboin, S. Efficient Synthesis of Unprotected C-5-Aryl/Heteroaryl-2’-deoxyuridine via a Suzuki-Miyaura Reaction in Aqueous Media. Molecules 2012, 17, 14409–14417. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, A.J.; Terhorst, T.J.; Matteucci, M.D.; Froehler, B.C. 5-Heteroaryl-2′-deoxyuridine Analogs. Synthesis and Incorporation into High-Affinity Oligonucleotides. J. Am. Chem. Soc. 1994, 116, 5540–5544. [Google Scholar] [CrossRef]
- Peyron, C.; Benhida, R.; Bories, C.; Loiseau, P.M. Synthesis and in vitro antileishmanial activity of 5-substituted-2’-deoxyuridine derivatives. Bioorgan. Chem. 2005, 33, 439–447. [Google Scholar] [CrossRef]
- Tian, M.; Yu, M.; Shi, T.; Hu, J.; Li, S.; Xu, J.; Chen, N.; Du, H. Silver-Catalyzed Direct C6-H Arylation of Purines and Purine Nucleosides with Arylboronic Acids. Eur. J. Org. Chem. 2017, 2017, 3415–3420. [Google Scholar] [CrossRef]
- Gayakhe, V.; Sanghvi, Y.S.; Fairlamb, I.J.; Kapdi, A.R. Catalytic C-H bond functionalisation of purine and pyrimidine nucleosides: A synthetic and mechanistic perspective. Chem. Commun. 2015, 51, 11944–11960. [Google Scholar] [CrossRef] [PubMed]
- Čerňová, M.; Čerňa, I.; Pohl, R.; Hocek, M. Regioselective Direct C–H Arylations of Protected Uracils. Synthesis of 5- and 6-Aryluracil Bases. J. Org. Chem. 2011, 76, 5309–5319. [Google Scholar] [CrossRef] [PubMed]
- Sahnoun, S.; Messaoudi, S.; Peyrat, J.F.; Brion, J.D.; Alami, M. Microwave-assisted Pd(OH)(2)-catalyzed direct C-H arylation of free-(NH2) adenines with aryl halides. Tetrahedron Lett. 2008, 49, 7279–7283. [Google Scholar] [CrossRef]
- Mori, Y.; Seki, M. Pd(OH)2/C (Pearlman’s Catalyst): A Highly Active Catalyst for Fukuyama, Sonogashira, and Suzuki Coupling Reactions. J. Org. Chem. 2003, 68, 1571–1574. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.Y.; Chen, L.; Doucet, H. Phosphine-Free Palladium-Catalyzed Direct Arylation of Imidazo[1,2-a]pyridines with Aryl Bromides at Low Catalyst Loading. J. Org. Chem. 2012, 77, 4473–4478. [Google Scholar] [CrossRef]
- Fuertes, M.A.; Cepeda, V.; Alonso, C.; Perez, J.M. Molecular mechanisms for the B-Z transition in the example of poly[d(G-C) x d(G-C)] polymers. A critical review. Chem. Rev. 2006, 106, 2045–2064. [Google Scholar] [CrossRef] [PubMed]
- Behe, M.; Felsenfeld, G. Effects of methylation on a synthetic polynucleotide: The B–Z transition in poly(dG-m5dC).poly(dG-m5dC). Proc. Natl. Acad. Sci. USA 1981, 78, 1619–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheardy, R.D.; Levine, N.; Marotta, S.; Suh, D.; Chaires, J.B. A thermodynamic investigation of the melting of B-Z junction forming DNA oligomers. Biochemistry 1994, 33, 1385–1391. [Google Scholar] [CrossRef]
- Taboury, J.A.; Adam, S.; Taillandier, E.; Neumann, J.M.; Tran-Dinh, S.; Huynh-Dinh, T.; Langlois d’Estaintot, B.; Conti, M.; Igolen, J. The B–Z transition in two synthetic oligonucleotides: D(C-2-amino-ACGTG) and d(m5CGCAm5CGTGCG) studied by IR, NMR and CD spectroscopies. Nucleic Acids Res. 1984, 12, 6291–6305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rich, A.; Nordheim, A.; Wang, A.H. The chemistry and biology of left-handed Z-DNA. Annu. Rev. Biochem. 1984, 53, 791–846. [Google Scholar] [CrossRef]
- Herbert, A. ALU non-B-DNA conformations, flipons, binary codes and evolution. R. Soc. Open Sci. 2020, 7, 200222. [Google Scholar] [CrossRef]
- Vongsutilers, V.; Shinohara, Y.; Kawai, G. Epigenetic TET-Catalyzed Oxidative Products of 5-Methylcytosine Impede Z-DNA Formation of CG Decamers. ACS Omega 2020, 5, 8056–8064. [Google Scholar] [CrossRef]
- Wang, S.; Long, Y.; Wang, J.; Ge, Y.; Guo, P.; Liu, Y.; Tian, T.; Zhou, X. Systematic investigations of different cytosine modifications on CpG dinucleotide sequences: The effects on the B-Z transition. J. Am. Chem. Soc. 2014, 136, 56–59. [Google Scholar] [CrossRef]
- Wang, J.; Wang, S.; Zhong, C.; Tian, T.; Zhou, X. Novel insights into a major DNA oxidative lesion: Its effects on Z-DNA stabilization. Org. Biomol. Chem. 2015, 13, 8996–8999. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Low, J.J.A.; Woon, E.C.Y. A general strategy exploiting m5C duplex-remodelling effect for selective detection of RNA and DNA m5C methyltransferase activity in cells. Nucleic Acids Res. 2020, 48, e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagawa, T.F.; Howell, M.L.; Tseng, K.; Ho, P.S. Effects of base substituents on the hydration of B- and Z-DNA: Correlations to the B- to Z-DNA transition. Nucleic Acids Res. 1993, 21, 5978–5986. [Google Scholar] [CrossRef] [Green Version]
- Fujii, S.; Wang, A.H.; van der Marel, G.; van Boom, J.H.; Rich, A. Molecular structure of (m5 dC-dG)3: The role of the methyl group on 5-methyl cytosine in stabilizing Z-DNA. Nucleic Acids Res. 1982, 10, 7879–7892. [Google Scholar] [CrossRef] [Green Version]
- Popenda, M.; Milecki, J.; Adamiak, R.W. High salt solution structure of a left-handed RNA double helix. Nucleic Acids Res. 2004, 32, 4044–4054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temiz, N.A.; Donohue, D.E.; Bacolla, A.; Luke, B.T.; Collins, J.R. The role of methylation in the intrinsic dynamics of B- and Z-DNA. PLoS ONE 2012, 7, e35558. [Google Scholar] [CrossRef]
- Belmont, P.; Constant, J.F.; Demeunynck, M. Nucleic acid conformation diversity: From structure to function and regulation. Chem. Soc. Rev. 2001, 30, 70–81. [Google Scholar] [CrossRef]
- Herbert, A.G.; Rich, A. A method to identify and characterize Z-DNA binding proteins using a linear oligodeoxynucleotide. Nucleic Acids Res. 1993, 21, 2669–2672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gislason, K.; Sigurdsson, S.T. Synthesis of a 5’-6-Locked, 1,10-Phenanthroline-Containing Nucleoside and Its Incorporation into DNA. Eur. J. Org. Chem. 2010, 2010, 4713–4718. [Google Scholar] [CrossRef]
- Hardin, C.C.; Zarling, D.A.; Puglisi, J.D.; Trulson, M.O.; Davis, P.W.; Tinoco, I., Jr. Stabilization of Z-RNA by chemical bromination and its recognition by anti-Z-DNA antibodies. Biochemistry 1987, 26, 5191–5199. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Sharma, S.; Chowdhury, S. Non-duplex G-Quadruplex Structures Emerge as Mediators of Epigenetic Modifications. Trends Genet. 2019, 35, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Hou, J.Q.; Xiang, H.D.; Yan, Y.Y.; Gu, Y.C.; Tan, J.H.; Li, D.; Gu, L.Q.; Ou, T.M.; Huang, Z.S. Stabilization of G-quadruplex DNA by C-5-methyl-cytosine in bcl-2 promoter: Implications for epigenetic regulation. Biochem. Biophys. Res. Commun. 2013, 433, 368–373. [Google Scholar] [CrossRef]
- Hardin, C.C.; Corregan, M.; Brown, B.A., 2nd; Frederick, L.N. Cytosine-cytosine+ base pairing stabilizes DNA quadruplexes and cytosine methylation greatly enhances the effect. Biochemistry 1993, 32, 5870–5880. [Google Scholar] [CrossRef]
- Xu, Y.; Sugiyama, H. Highly Efficient Photochemical 2′-Deoxyribonolactone Formation at the Diagonal Loop of a 5-Iodouracil-Containing Antiparallel G-Quartet. J. Am. Chem. Soc. 2004, 126, 6274–6279. [Google Scholar] [CrossRef]
- Li, Y.; Sugiyama, H. Photoreactivity of the linker region of two consecutive G-quadruplexes formed by human telomeric DNA. Chem. Comm. 2015, 51, 8861–8864. [Google Scholar] [CrossRef] [Green Version]
- Heinen, L.; Walther, A. Temporal control of i-motif switch lifetimes for autonomous operation of transient DNA nanostructures. Chem. Sci. 2017, 8, 4100–4107. [Google Scholar] [CrossRef] [Green Version]
- Gueron, M.; Leroy, J.L. The i-motif in nucleic acids. Curr. Opin. Struct. Biol. 2000, 10, 326–331. [Google Scholar] [CrossRef]
- Bielecka, P.; Dembska, A.; Juskowiak, B. Monitoring of pH Using an i-Motif-Forming Sequence Containing a Fluorescent Cytosine Analogue, tC. Molecules 2019, 24, 952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhavsar-Jog, Y.P.; Van Dornshuld, E.; Brooks, T.A.; Tschumper, G.S.; Wadkins, R.M. Epigenetic modification, dehydration, and molecular crowding effects on the thermodynamics of i-motif structure formation from C-rich DNA. Biochemistry 2014, 53, 1586–1594. [Google Scholar] [CrossRef] [PubMed]
- Abou Assi, H.; Garavís, M.; González, C.; Damha, M.J. i-Motif DNA: Structural features and significance to cell biology. Nucleic Acids Res. 2018, 46, 8038–8056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, E.P.; Lamparska, K.; Smith, S.S.; Waller, Z.A.E. Substitution of Cytosine with Guanylurea Decreases the Stability of i-Motif DNA. Biochemistry 2017, 56, 4879–4883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lannes, L.; Halder, S.; Krishnan, Y.; Schwalbe, H. Tuning the pH Response of i-Motif DNA Oligonucleotides. Chembiochem 2015, 16, 1647–1656. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.P.; Abdelhamid, M.A.S.; Ehiabor, M.O.; Grigg, M.C.; Irving, K.; Smith, N.M.; Waller, Z.A.E. Epigenetic modification of cytosines fine tunes the stability of i-motif DNA. Nucleic Acids Res. 2020, 48, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Devi, G.; Shao, F. Regulation of telomeric i-motif stability by 5-methylcytosine and 5-hydroxymethylcytosine modification. Org. Biomol. Chem. 2015, 13, 5646–5651. [Google Scholar] [CrossRef] [PubMed]
- Benabou, S.; Aviñó, A.; Eritja, R.; González, C.; Gargallo, R. Fundamental aspects of the nucleic acid i-motif structures. RSC Adv. 2014, 4, 26956–26980. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Leroy, J.L.; Gueron, M. An intramolecular i-motif: The solution structure and base-pair opening kinetics of d(5mCCT3CCT3ACCT3CC). J. Mol. Biol. 1998, 278, 949–965. [Google Scholar] [CrossRef]
- Phan, A.T.; Gueron, M.; Leroy, J.L. The solution structure and internal motions of a fragment of the cytidine-rich strand of the human telomere. J. Mol. Biol. 2000, 299, 123–144. [Google Scholar] [CrossRef]
- Kendrick, S.; Akiyama, Y.; Hecht, S.M.; Hurley, L.H. The i-motif in the bcl-2 P1 promoter forms an unexpectedly stable structure with a unique 8:5:7 loop folding pattern. J. Am. Chem. Soc. 2009, 131, 17667–17676. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Rodgers, M.T. Base-Pairing Energies of Protonated Nucleoside Base Pairs of dCyd and m(5)dCyd: Implications for the Stability of DNA i-Motif Conformations. J. Am. Soc. Mass Spectrom. 2015, 26, 1394–1403. [Google Scholar] [CrossRef] [Green Version]
- Assi, H.A.; Harkness, R.W.; Martin-Pintado, N.; Wilds, C.J.; Campos-Olivas, R.; Mittermaier, A.K.; Gonzalez, C.; Damha, M.J. Stabilization of i-motif structures by 2’-beta-fluorination of DNA. Nucleic Acids Res. 2016, 44, 4998–5009. [Google Scholar] [CrossRef] [Green Version]
- Osborne, S.D.; Powers, V.E.; Rusling, D.A.; Lack, O.; Fox, K.R.; Brown, T. Selectivity and affinity of triplex-forming oligonucleotides containing 2’-aminoethoxy-5-(3-aminoprop-1-ynyl)uridine for recognizing AT base pairs in duplex DNA. Nucleic Acids Res. 2004, 32, 4439–4447. [Google Scholar] [CrossRef]
- Frank-Kamenetskii, M.D.; Mirkin, S.M. Triplex DNA structures. Annu. Rev. Biochem. 1995, 64, 65–95. [Google Scholar] [CrossRef]
- Kumar, P.; Sharma, P.K.; Hansen, J.; Jedinak, L.; Reslow-Jacobsen, C.; Hornum, M.; Nielsen, P. Three new double-headed nucleotides with additional nucleobases connected to C-5 of pyrimidines; synthesis, duplex and triplex studies. Bioorgan. Med. Chem. 2016, 24, 742–749. [Google Scholar] [CrossRef]
- Brazier, J.A.; Shibata, T.; Townsley, J.; Taylor, B.F.; Frary, E.; Williams, N.H.; Williams, D.M. Amino-functionalized DNA: The properties of C5-amino-alkyl substituted 2’-deoxyuridines and their application in DNA triplex formation. Nucleic Acids Res. 2005, 33, 1362–1371. [Google Scholar] [CrossRef] [Green Version]
- Kanamori, T.; Masaki, Y.; Mizuta, M.; Tsunoda, H.; Ohkubo, A.; Sekine, M.; Seio, K. DNA duplexes and triplex-forming oligodeoxynucleotides incorporating modified nucleosides forming stable and selective triplexes. Org. Biomol. Chem. 2012, 10, 1007–1013. [Google Scholar] [CrossRef]
- Rusling, D.A.; Peng, G.; Srinivasan, N.; Fox, K.R.; Brown, T. DNA triplex formation with 5-dimethylaminopropargyl deoxyuridine. Nucleic Acids Res. 2009, 37, 1288–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sau, S.P.; Kumar, P.; Anderson, B.A.; Ostergaard, M.E.; Deobald, L.; Paszczynski, A.; Sharma, P.K.; Hrdlicka, P.J. Optimized DNA-targeting using triplex forming C5-alkynyl functionalized LNA. Chem. Commun. 2009, 6756–6758. [Google Scholar] [CrossRef]
- Devi, G.; Zhou, Y.; Zhong, Z.; Toh, D.F.; Chen, G. RNA triplexes: From structural principles to biological and biotech applications. Wiley Interdiscip. Rev. RNA 2015, 6, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Duca, M.; Vekhoff, P.; Oussedik, K.; Halby, L.; Arimondo, P.B. The triple helix: 50 years later, the outcome. Nucleic Acids Res. 2008, 36, 5123–5138. [Google Scholar] [CrossRef] [PubMed]
- McKinney, J.A.; Wang, G.; Mukherjee, A.; Christensen, L.; Subramanian, S.H.S.; Zhao, J.; Vasquez, K.M. Distinct DNA repair pathways cause genomic instability at alternative DNA structures. Nat. Commun. 2020, 11, 236. [Google Scholar] [CrossRef] [Green Version]
- Sipa, K.; Sochacka, E.; Kazmierczak-Baranska, J.; Maszewska, M.; Janicka, M.; Nowak, G.; Nawrot, B. Effect of base modifications on structure, thermodynamic stability, and gene silencing activity of short interfering RNA. RNA 2007, 13, 1301–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]









| Substrate | Entry | Base/Ligand and Solvent | Temp (°C) | Aryl | Yield (%) | Reference |
|---|---|---|---|---|---|---|
| 1 | 1 | NaCO3/TPPTS DMSO | 90 °C |  | 91% | [75] |
| 2 | 2 | NaCO3/TPPTS H2O | 80 °C |  | 85% | [76] |
| 3 | NaCO3/TPPTS H2O | 80 °C | 67% | |||
| 4 | NaCO3/TPPTS H2O | 80 °C | 83% | |||
| 5 | NaCO3/TPPTS H2O | 80 °C | 83% | |||
| 6 | NaCO3/TPPTS H2O | 80 °C | 72% | |||
| 3 | 7 | CuI/piperidine DMF | 150 °C |  | 55% | [62] |
| 8 | CuI/piperidine DMF | 150 °C | 68% | |||
| 9 | CuI/piperidine DMF | 150 °C | 55% | |||
| 10 | Cs2CO3/CuI DMF | 120 °C | 66% | [77] | ||
| 4 | 11 | CuI/PPh3 THF | 90 °C |  | 95% | [78] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balasubramaniyam, T.; Oh, K.-I.; Jin, H.-S.; Ahn, H.-B.; Kim, B.-S.; Lee, J.-H. Non-Canonical Helical Structure of Nucleic Acids Containing Base-Modified Nucleotides. Int. J. Mol. Sci. 2021, 22, 9552. https://doi.org/10.3390/ijms22179552
Balasubramaniyam T, Oh K-I, Jin H-S, Ahn H-B, Kim B-S, Lee J-H. Non-Canonical Helical Structure of Nucleic Acids Containing Base-Modified Nucleotides. International Journal of Molecular Sciences. 2021; 22(17):9552. https://doi.org/10.3390/ijms22179552
Chicago/Turabian StyleBalasubramaniyam, Thananjeyan, Kwnag-Im Oh, Ho-Seong Jin, Hye-Bin Ahn, Byeong-Seon Kim, and Joon-Hwa Lee. 2021. "Non-Canonical Helical Structure of Nucleic Acids Containing Base-Modified Nucleotides" International Journal of Molecular Sciences 22, no. 17: 9552. https://doi.org/10.3390/ijms22179552