
Roles of High Osmolarity Glycerol and Cell Wall Integrity Pathways in Cadmium Toxicity in Saccharomyces cerevisiae


Abstract
:1. Introduction
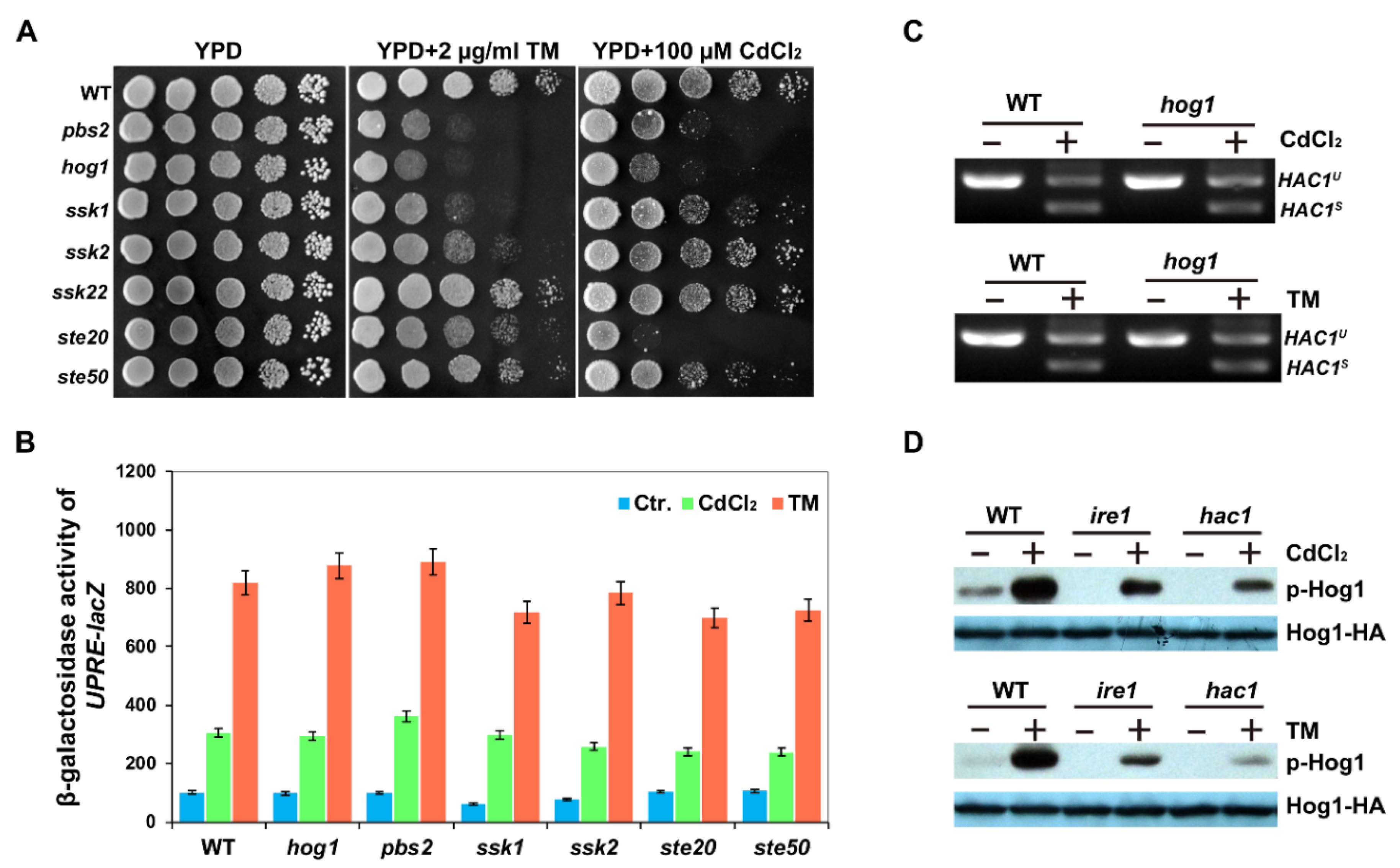
2. Results
2.1. UPR Pathway Is Required for Cadmium-Induced Activation of Hog1
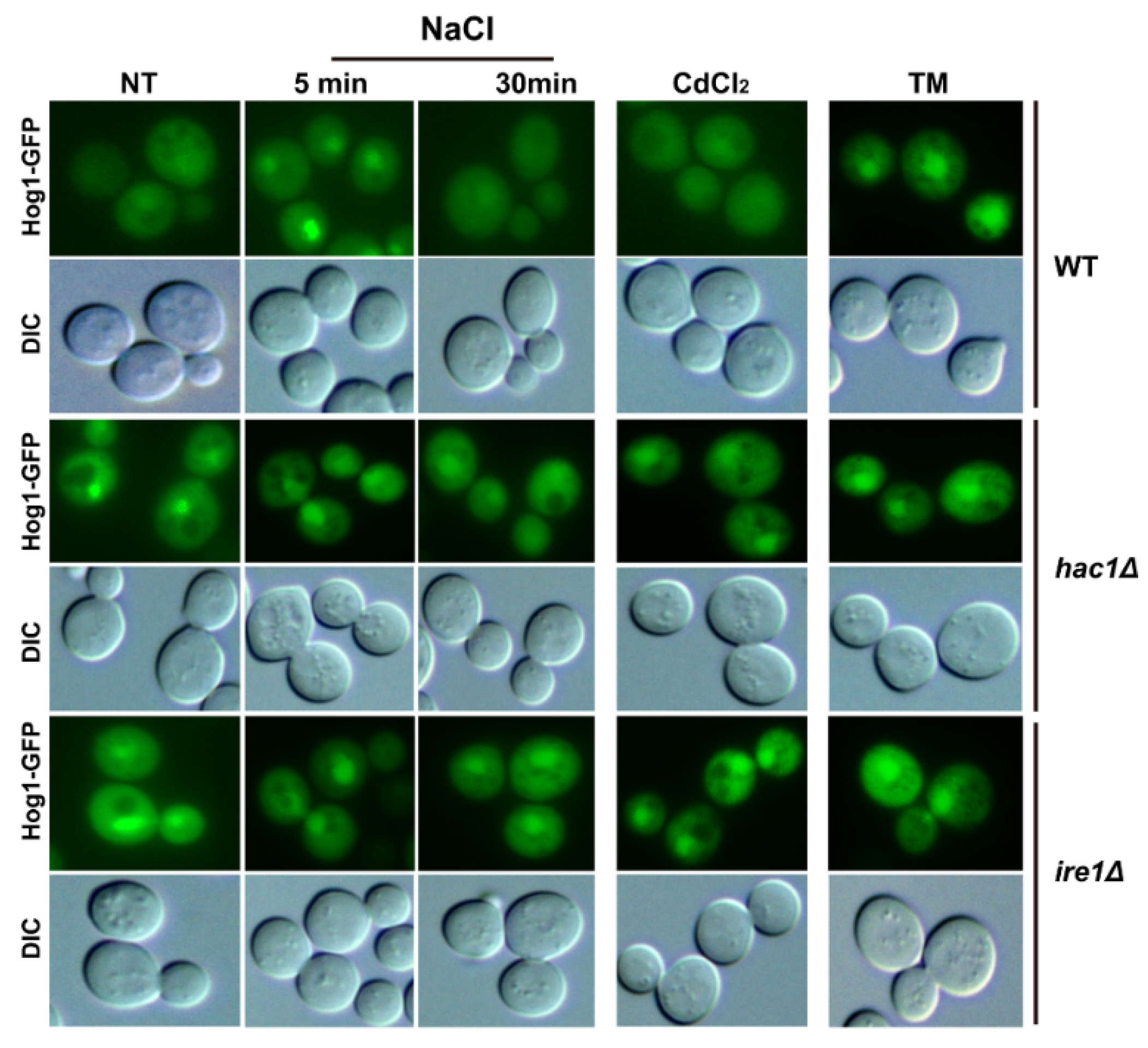
2.2. Ire1 and Hac1 Regulate the Nuclear Localisation of Hog1
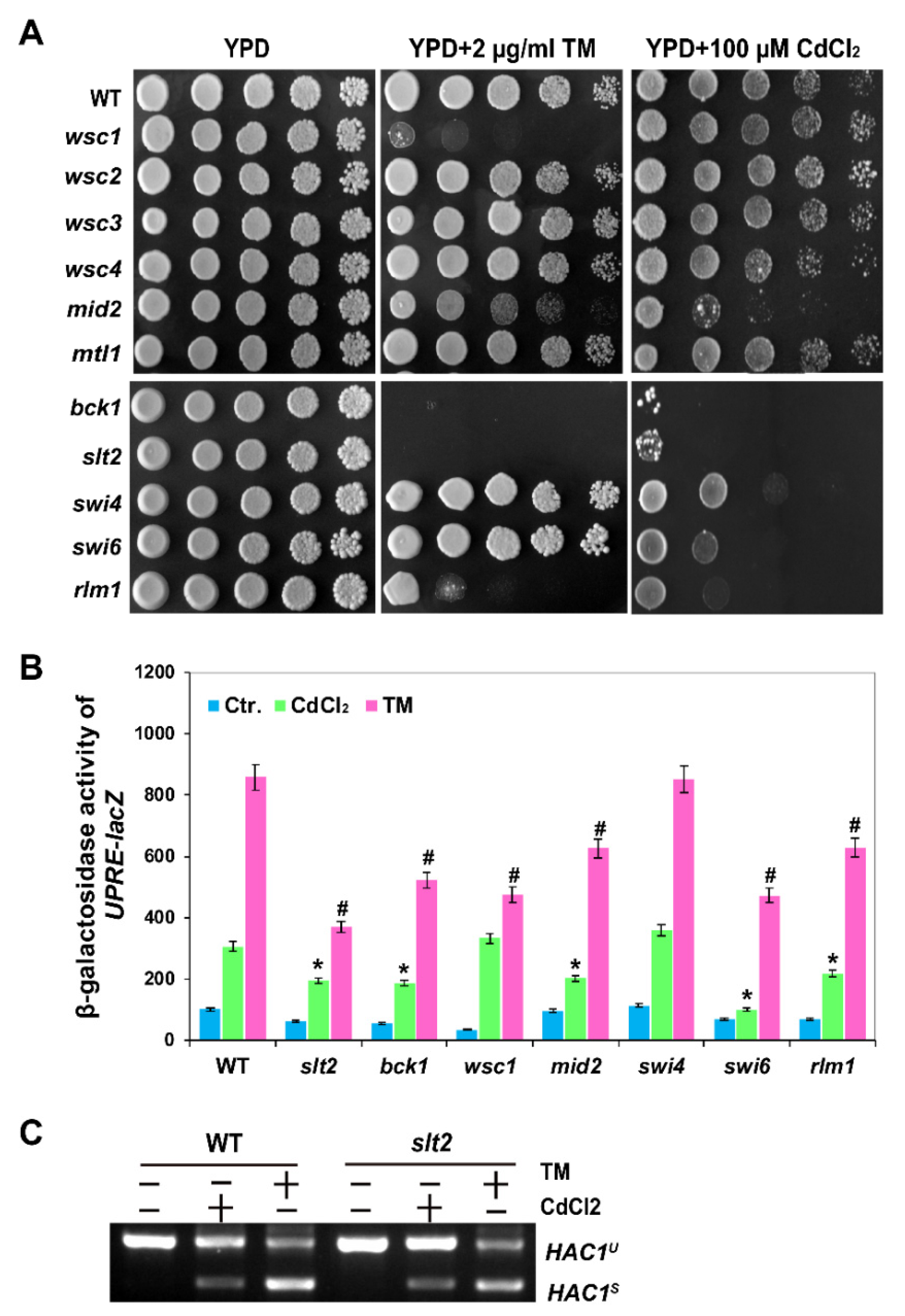
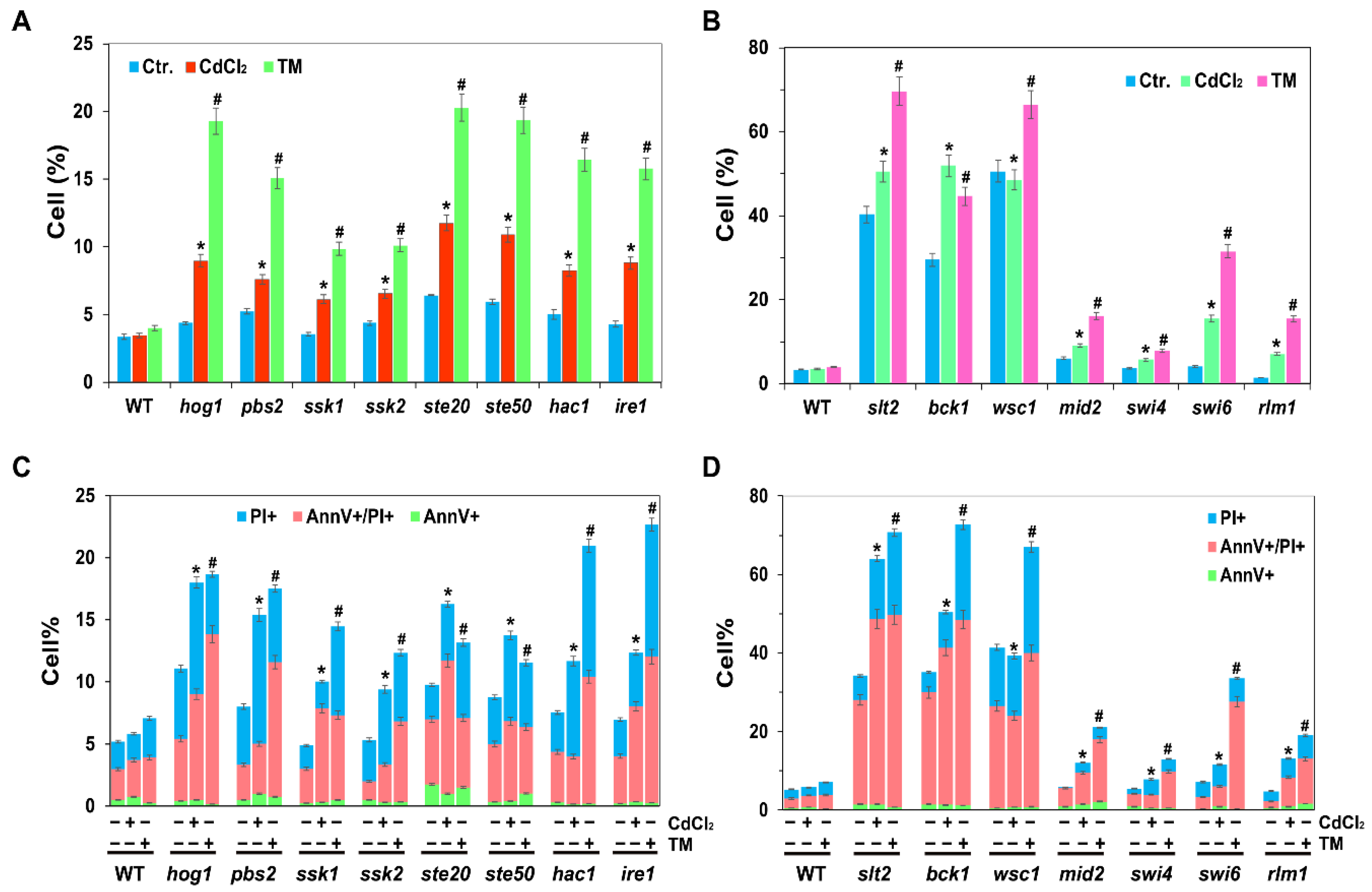
2.3. The MAPK Slt2 Is Required for Both Cadmium- and Tm-Induced UPR Pathways
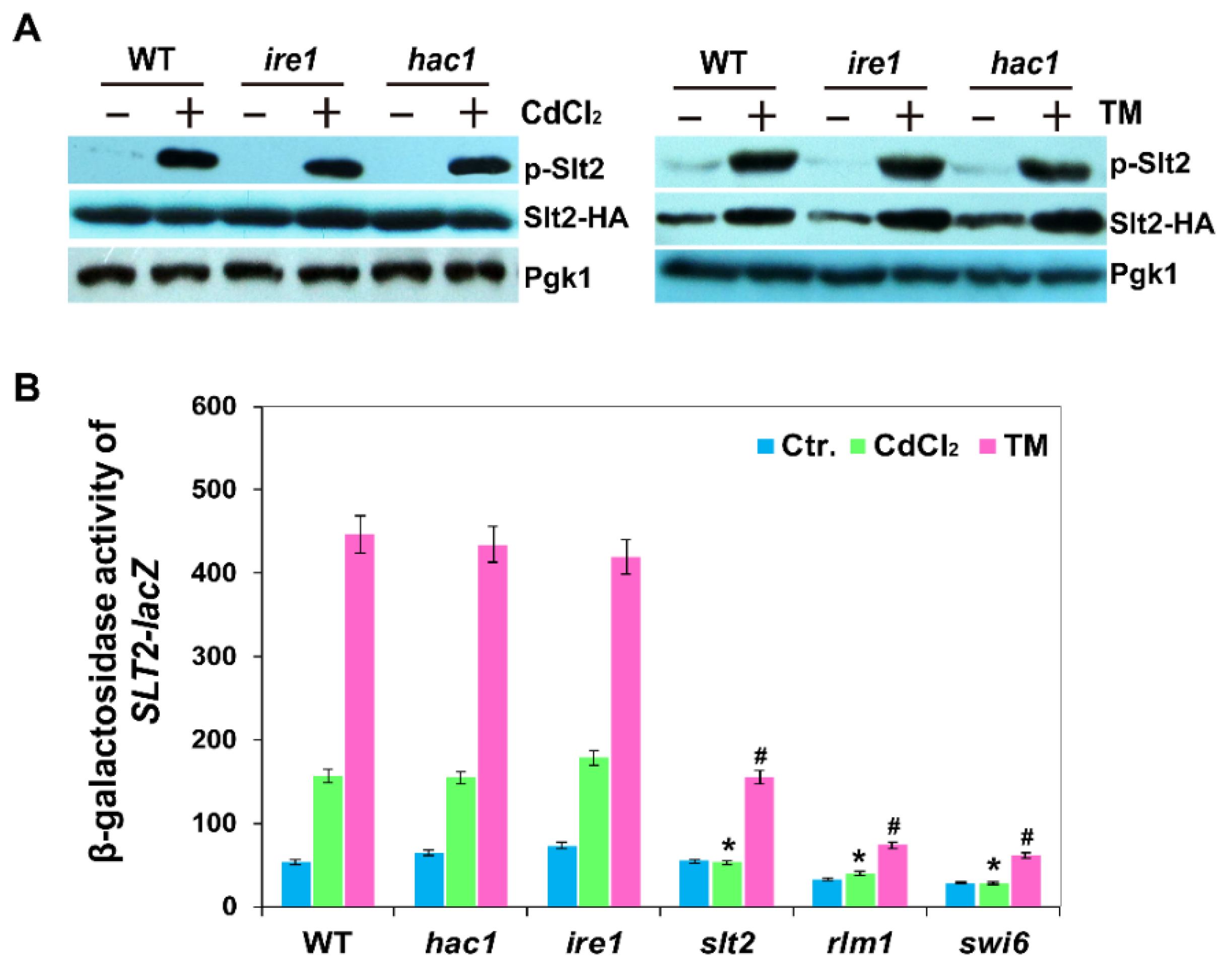
2.4. Phosphorylation and Expression of Slt2 Is Independent of the Ire1-Mediated UPR Pathway
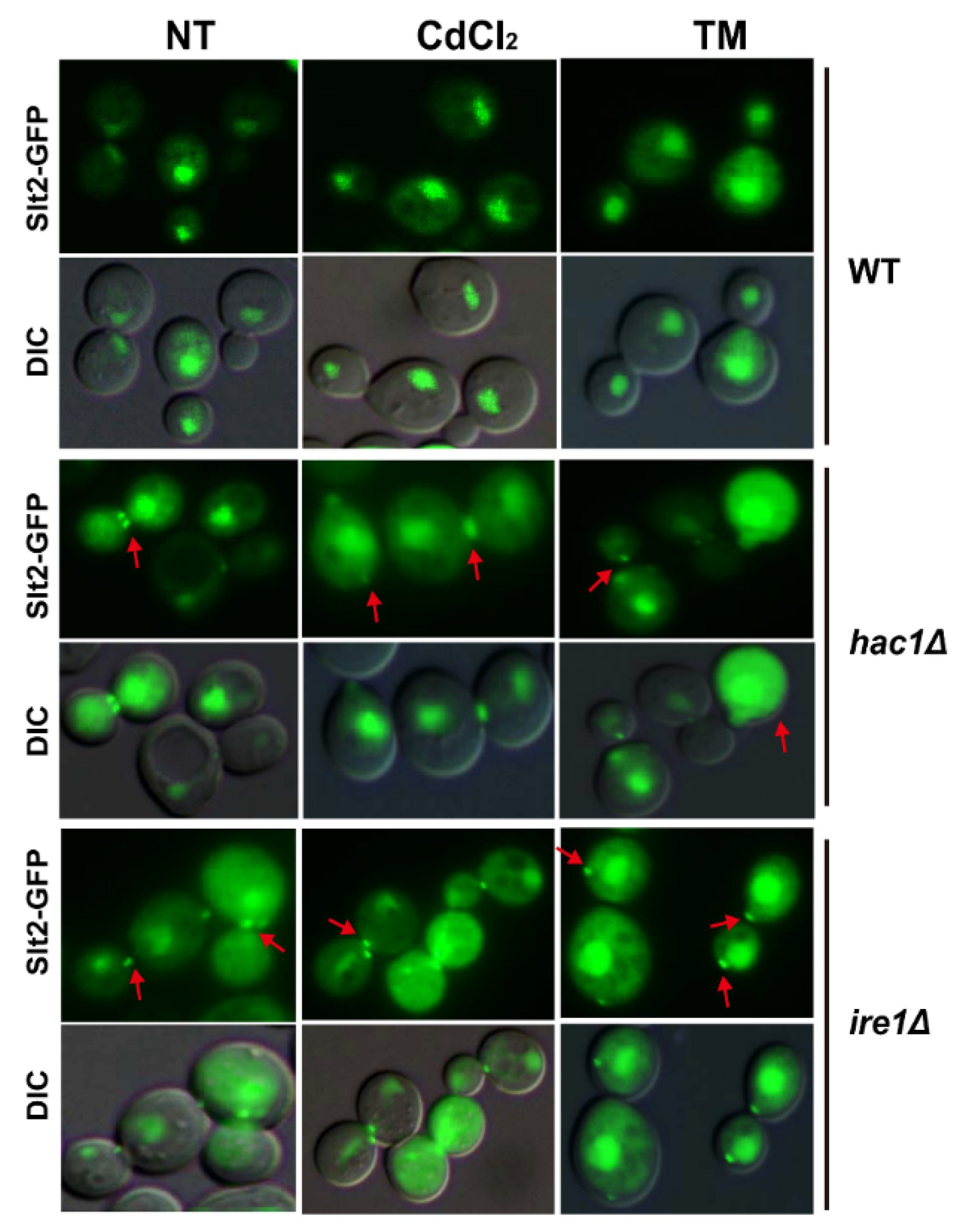
2.5. Defection of UPR Pathway Promotes Slt2 to Localise in Cytosol and Bud Neck
2.6. UPR, HOG and CWI Pathways Are All Required for Cadmium- and Tm-Induced ROS and Cell Death
3. Discussion
4. Materials and Methods
4.1. Yeast Strains and Growth Media
4.2. Phenotype Assay
4.3. DNA Manipulations
4.4. Microscopy Assay
4.5. Intracellular ROS and Cell Death Assay
4.6. β-Galactosidase Activity Assay
4.7. RNA Extraction and RT-PCR Analysis
4.8. Protein Extract Preparation and Western Blotting Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McGregor, D.B.; Baan, R.A.; Partensky, C.; Rice, J.M.; Wilbourn, J.D. Evaluation of the carcinogenic risks to humans associated with surgical implants and other foreign bodies—A report of an IARC Monographs Programme Meeting. International Agency for Research on Cancer. Eur. J. Cancer 2000, 36, 307–313. [Google Scholar] [CrossRef]
- Cox, L.A., Jr. Quantifying potential health impacts of cadmium in cigarettes on smoker risk of lung cancer: A portfolio-of-mechanisms approach. Risk Anal. 2006, 26, 1581–1599. [Google Scholar] [CrossRef] [PubMed]
- Stohs, S.J.; Bagchi, D. Oxidative mechanisms in the toxicity of metal ions. Free Radic Biol. Med. 1995, 18, 321–336. [Google Scholar] [CrossRef] [Green Version]
- Fortuniak, A.; Zadzinski, R.; Bilinski, T.; Bartosz, G. Glutathione depletion in the yeast Saccharomyces cerevisiae. Biochem. Mol. Biol. Int. 1996, 38, 901–910. [Google Scholar] [PubMed]
- Stohs, S.J.; Bagchi, D.; Hassoun, E.; Bagchi, M. Oxidative mechanisms in the toxicity of chromium and cadmium ions. J. Environ. Pathol. Toxicol. Oncol. 2001, 20, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Bertin, G.; Averbeck, D. Cadmium: Cellular effects, modifications of biomolecules, modulation of DNA repair and genotoxic consequences (a review). Biochimie 2006, 88, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Lam, Y.T.; Aung-Htut, M.T.; Lim, Y.L.; Yang, H.; Dawes, I.W. Changes in reactive oxygen species begin early during replicative aging of Saccharomyces cerevisiae cells. Free Radic Biol. Med. 2011, 50, 963–970. [Google Scholar] [CrossRef]
- Nargund, A.M.; Avery, S.V.; Houghton, J.E. Cadmium induces a heterogeneous and caspase-dependent apoptotic response in Saccharomyces cerevisiae. Apoptosis 2008, 13, 811–821. [Google Scholar] [CrossRef]
- Wang, S.H.; Shih, Y.L.; Ko, W.C.; Wei, Y.H.; Shih, C.M. Cadmium-induced autophagy and apoptosis are mediated by a calcium signaling pathway. Cell Mol. Life Sci. 2008, 65, 3640–3652. [Google Scholar] [CrossRef]
- Xu, C.; Bailly-Maitre, B.; Reed, J.C. Endoplasmic reticulum stress: Cell life and death decisions. J. Clin. Investig. 2005, 115, 2656–2664. [Google Scholar] [CrossRef] [Green Version]
- Orrenius, S.; Zhivotovsky, B.; Nicotera, P. Regulation of cell death: The calcium-apoptosis link. Nat. Rev. Mol. Cell Biol. 2003, 4, 552–565. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, M.; Cunningham, K.W. Mitogen-activated protein kinase stimulation of Ca(2+) signaling is required for survival of endoplasmic reticulum stress in yeast. Mol. Biol. Cell 2003, 14, 4296–4305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hampton, R.Y. ER-associated degradation in protein quality control and cellular regulation. Curr. Opin. Cell Biol. 2002, 14, 476–482. [Google Scholar] [CrossRef]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef]
- Cox, J.S.; Walter, P. A novel mechanism for regulating activity of a transcription factor that controls the unfolded protein response. Cell 1996, 87, 391–404. [Google Scholar] [CrossRef] [Green Version]
- Travers, K.J.; Patil, C.K.; Wodicka, L.; Lockhart, D.J.; Weissman, J.S.; Walter, P. Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation. Cell 2000, 101, 249–258. [Google Scholar] [CrossRef] [Green Version]
- Shuda, M.; Kondoh, N.; Imazeki, N.; Tanaka, K.; Okada, T.; Mori, K.; Hada, A.; Arai, M.; Wakatsuki, T.; Matsubara, O.; et al. Activation of the ATF6, XBP1 and grp78 genes in human hepatocellular carcinoma: A possible involvement of the ER stress pathway in hepatocarcinogenesis. J. Hepatol. 2003, 38, 605–614. [Google Scholar] [CrossRef]
- Fernandez, P.M.; Tabbara, S.O.; Jacobs, L.K.; Manning, F.C.; Tsangaris, T.N.; Schwartz, A.M.; Kennedy, K.A.; Patierno, S.R. Overexpression of the glucose-regulated stress gene GRP78 in malignant but not benign human breast lesions. Breast Cancer Res. Treat. 2000, 59, 15–26. [Google Scholar] [CrossRef]
- Maity, S.; Rajkumar, A.; Matai, L.; Bhat, A.; Ghosh, A.; Agam, G.; Kaur, S.; Bhatt, N.R.; Mukhopadhyay, A.; Sengupta, S.; et al. Oxidative homeostasis regulates the response to reductive endoplasmic reticulum stress through translation control. Cell Rep. 2016, 16, 851–865. [Google Scholar] [CrossRef] [Green Version]
- Aragon, T.; van Anken, E.; Pincus, D.; Serafimova, I.M.; Korennykh, A.V.; Rubio, C.A.; Walter, P. Messenger RNA targeting to endoplasmic reticulum stress signalling sites. Nature 2009, 457, 736–740. [Google Scholar] [CrossRef] [Green Version]
- Kimata, Y.; Ishiwata-Kimata, Y.; Ito, T.; Hirata, A.; Suzuki, T.; Oikawa, D.; Takeuchi, M.; Kohno, K. Two regulatory steps of ER-stress sensor Ire1 involving its cluster formation and interaction with unfolded proteins. J. Cell Biol. 2007, 179, 75–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardarin, A.; Chedin, S.; Lagniel, G.; Aude, J.C.; Godat, E.; Catty, P.; Labarre, J. Endoplasmic reticulum is a major target of cadmium toxicity in yeast. Mol. Microbiol. 2010, 76, 1034–1048. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Feldman, D.E.; Deng, C.C.; Brown, J.A.; De Giacomo, A.F.; Gaw, A.F.; Shi, G.Y.; Le, Q.T.; Brown, J.M.; Koong, A.C. Identification of mitogen-activated protein kinase signaling pathways that confer resistance to endoplasmic reticulum stress in Saccharomyces cerevisiae. Mol. Cancer Res. 2005, 3, 669–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilsland, E.; Molin, C.; Swaminathan, S.; Ramne, A.; Sunnerhagen, P. Rck1 and Rck2 MAPKAP kinases and the HOG pathway are required for oxidative stress resistance. Mol. Microbiol. 2004, 53, 1743–1756. [Google Scholar] [CrossRef]
- Xiong, B.; Zhang, L.L.; Xu, H.H.; Yang, Y.; Jiang, L.H. Cadmium induces the activation of cell wall integrity pathway in budding yeast. Chem.-Biol. Interact. 2015, 240, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.H.; Cao, C.L.; Zhang, L.L.; Lin, W.; Xia, J.; Xu, H.H.; Zhang, Y. Cadmium-induced activation of high osmolarity glycerol pathway through its Sln1 branch is dependent on the MAP kinase kinase kinase Ssk2, but not its paralog Ssk22, in budding yeast. FEMS Yeast Res. 2014, 14, 1263–1272. [Google Scholar] [CrossRef]
- Brewster, J.L.; de Valoir, T.; Dwyer, N.D.; Winter, E.; Gustin, M.C. An osmosensing signal transduction pathway in yeast. Science 1993, 259, 1760–1763. [Google Scholar] [CrossRef] [PubMed]
- Bermejo, C.; Rodriguez, E.; Garcia, R.; Rodriguez-Pena, J.M.; de la Concepcion, M.L.R.; Rivas, C.; Arias, P.; Nombela, C.; Posas, F.; Arroyo, J. The sequential activation of the yeast HOG and SLT2 pathways is required for cell survival to cell wall stress. Mol. Biol. Cell 2008, 19, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Garcia, R.; Rodriguez-Pena, J.M.; Bermejo, C.; Nombela, C.; Arroyo, J. The high osmotic response and cell wall integrity pathways cooperate to regulate transcriptional responses to zymolyase-induced cell wall stress in Saccharomyces cerevisiae. J. Biol. Chem. 2009, 284, 10901–10911. [Google Scholar] [CrossRef] [Green Version]
- Bicknell, A.A.; Tourtellotte, J.; Niwa, M. Late phase of the endoplasmic reticulum stress response pathway is regulated by Hog1 MAP kinase. J. Biol. Chem. 2010, 285, 17545–17555. [Google Scholar] [CrossRef] [Green Version]
- Bernales, S.; McDonald, K.L.; Walter, P. Autophagy counterbalances endoplasmic reticulum expansion during the unfolded protein response. PLoS Biol. 2006, 4, e423. [Google Scholar] [CrossRef] [Green Version]
- Yorimitsu, T.; Nair, U.; Yang, Z.; Klionsky, D.J. Endoplasmic reticulum stress triggers autophagy. J. Biol. Chem. 2006, 281, 30299–30304. [Google Scholar] [CrossRef] [Green Version]
- Levin, D.E. Regulation of cell wall biogenesis in Saccharomyces cerevisiae: The cell wall integrity signaling pathway. Genetics 2011, 189, 1145–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonilla, M.; Nastase, K.K.; Cunningham, K.W. Essential role of calcineurin in response to endoplasmic reticulum stress. EMBO J. 2002, 21, 2343–2353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babour, A.; Bicknell, A.A.; Tourtellotte, J.; Niwa, M. A surveillance pathway monitors the fitness of the endoplasmic reticulum to control its inheritance. Cell 2010, 142, 256–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scrimale, T.; Didone, L.; Bentley, K.L.D.M.; Krysan, D.J. The unfolded protein response is induced by the cell wall integrity mitogen-activated protein kinase signaling cascade and is required for cell wall integrity in Saccharomyces cerevisiae. Mol. Biol. Cell 2009, 20, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Levin, D.E. Cell wall integrity signaling in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2005, 69, 262–291. [Google Scholar] [CrossRef] [Green Version]
- Torres-Quiroz, F.; Garcia-Marques, S.; Coria, R.; Randez-Gil, F.; Prieto, J.A. The activity of yeast Hog1 MAPK is required during endoplasmic reticulum stress induced by tunicamycin exposure. J. Biol. Chem. 2010, 285, 20088–20096. [Google Scholar] [CrossRef] [Green Version]
- Proft, M.; Mas, G.; de Nadal, E.; Vendrell, A.; Noriega, N.; Struhl, K.; Posas, F. The stress-activated Hog1 kinase is a selective transcriptional elongation factor for genes responding to osmotic stress. Mol. Cell 2006, 23, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Proft, M.; Pascual-Ahuir, A.; de Nadal, E.; Arino, J.; Serrano, R.; Posas, F. Regulation of the Sko1 transcriptional repressor by the Hog1 MAP kinase in response to osmotic stress. EMBO J. 2001, 20, 1123–1133. [Google Scholar] [CrossRef]
- Pelet, S.; Dechant, R.; Lee, S.S.; van Drogen, F.; Peter, M. An integrated image analysis platform to quantify signal transduction in single cells. Integr. Biol. 2012, 4, 1274–1282. [Google Scholar] [CrossRef] [Green Version]
- Philip, B.; Levin, D.E. Wsc1 and Mid2 are cell surface sensors for cell wall integrity signaling that act through Rom2, a guanine nucleotide exchange factor for Rho1. Mol. Cell. Biol. 2001, 21, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.Y.; Truman, A.W.; Levin, D.E. Yeast mpk1 mitogen-activated protein kinase activates transcription through Swi4/Swi6 by a noncatalytic mechanism that requires upstream signal. Mol. Cell. Biol. 2008, 28, 2579–2589. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Takaesu, G.; Hagiwara, M.; Irie, K.; Matsumoto, K. Characterization of a serum response factor-like protein in Saccharomyces cerevisiae, Rlm1, which has transcriptional activity regulated by the Mpk1 (Slt2) mitogen-activated protein kinase pathway. Mol. Cell. Biol. 1997, 17, 2615–2623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, U.S.; Levin, D.E. Genome-wide analysis of gene expression regulated by the yeast cell wall integrity signalling pathway. Mol. Microbiol. 1999, 34, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Pulido, M.D.; Parrish, A.R. Metal-induced apoptosis: Mechanisms. Mutat. Res. 2003, 533, 227–241. [Google Scholar] [CrossRef]
- Mao, K.; Wang, K.; Zhao, M.; Xu, T.; Klionsky, D.J. Two MAPK-signaling pathways are required for mitophagy in Saccharomyces cerevisiae. J. Cell Biol. 2011, 193, 755–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westfall, P.J.; Ballon, D.R.; Thorner, J. When the stress of your environment makes you go HOG wild. Science 2004, 306, 1511–1512. [Google Scholar] [CrossRef]
- Jarup, L.; Berglund, M.; Elinder, C.G.; Nordberg, G.; Vahter, M. Health effects of cadmium exposure--a review of the literature and a risk estimate. Scand. J. Work Environ. Health 1998, 24 (Suppl. 1), 1–51. [Google Scholar]
- Nair, A.R.; Lee, W.K.; Smeets, K.; Swennen, Q.; Sanchez, A.; Thevenod, F.; Cuypers, A. Glutathione and mitochondria determine acute defense responses and adaptive processes in cadmium-induced oxidative stress and toxicity of the kidney. Arch. Toxicol. 2015, 89, 2273–2289. [Google Scholar] [CrossRef]
- Kitamura, M.; Hiramatsu, N. The oxidative stress: Endoplasmic reticulum stress axis in cadmium toxicity. BioMetals 2010, 23, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Posas, F.; Saito, H. Activation of the yeast SSK2 MAP kinase kinase kinase by the SSK1 two-component response regulator. EMBO J. 1998, 17, 1385–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattison, C.P.; Ota, I.M. Two protein tyrosine phosphatases, Ptp2 and Ptp3, modulate the subcellular localization of the Hog1 MAP kinase in yeast. Genes Dev. 2000, 14, 1229–1235. [Google Scholar] [PubMed]
- Li, X.; Du, Y.R.; Siegel, S.; Ferro-Novick, S.; Novick, P. Activation of the mitogen-activated protein kinase, Slt2p, at bud tips blocks a late stage of endoplasmic reticulum inheritance in Saccharomyces cerevisiae. Mol. Biol. Cell 2010, 21, 1772–1782. [Google Scholar] [CrossRef] [Green Version]
- Winzeler, E.A.; Shoemaker, D.D.; Astromoff, A.; Liang, H.; Anderson, K.; Andre, B.; Bangham, R.; Benito, R.; Boeke, J.D.; Bussey, H.; et al. Functional characterization of the S. cerevisiae genome by gene deletion and parallel analysis. Science 1999, 285, 901–906. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Du, J.; Xiong, B.; Xu, H.; Jiang, L. ESCRT components regulate the expression of the ER/Golgi calcium pump gene PMR1 through the Rim101/Nrg1 pathway in budding yeast. J. Mol. Cell Biol. 2013, 5, 336–344. [Google Scholar] [CrossRef] [Green Version]
- Longtine, M.S.; McKenzie, A., 3rd; Demarini, D.J.; Shah, N.G.; Wach, A.; Brachat, A.; Philippsen, P.; Pringle, J.R. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 1998, 14, 953–961. [Google Scholar] [CrossRef]
- Madeo, F.; Frohlich, E.; Frohlich, K.U. A yeast mutant showing diagnostic markers of early and late apoptosis. J. Cell Biol. 1997, 139, 729–734. [Google Scholar] [CrossRef] [Green Version]
- Ellis, C.D.; Wang, F.; MacDiarmid, C.W.; Clark, S.; Lyons, T.; Eide, D.J. Zinc and the Msc2 zinc transporter protein are required for endoplasmic reticulum function. J. Cell Biol. 2004, 166, 325–335. [Google Scholar] [CrossRef] [Green Version]
- Kohrer, K.; Domdey, H. Preparation of high molecular weight RNA. Methods Enzymol. 1991, 194, 398–405. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Relevant Genotype | Source/Reference |
|---|---|---|
| BY4741 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | [55] |
| pbs2Δ | BY4741 pbs2::kanMX4 | [55] |
| hog1Δ | BY4741 hog1::kanMX4 | [55] |
| ssk1Δ | BY4741 ssk1::kanMX4 | [55] |
| ssk2Δ | BY4741 ssk2::kanMX4 | [55] |
| ssk22Δ | BY4741 ssk22::kanMX4 | [55] |
| ste20Δ | BY4741 ste20::kanMX4 | [55] |
| ste50Δ | BY4741 ste50::kanMX4 | [55] |
| ire1Δ | BY4741 ire1::kanMX4 | [55] |
| hac1Δ | BY4741 hac1::kanMX4 | [55] |
| wsc1Δ | BY4741 wsc1::kanMX4 | [55] |
| wsc2Δ | BY4741 wsc2::kanMX4 | [55] |
| wsc3Δ | BY4741 wsc3::kanMX4 | [55] |
| wsc4Δ | BY4741 wsc4::kanMX4 | [55] |
| mid2Δ | BY4741 mid2::kanMX4 | [55] |
| mtl1Δ | BY4741 mtl1::kanMX4 | [55] |
| bck1Δ | BY4741 bck1::kanMX4 | [55] |
| slt2Δ | BY4741 slt2::kanMX4 | [55] |
| swi4Δ | BY4741 swi4::kanMX4 | [55] |
| swi6Δ | BY4741 swi6::kanMX4 | [55] |
| rlm1Δ | BY4741 rlm1::kanMX4 | [55] |
| BY4741 HOG1-HA | BY4741 HOG1-HA-HIS3MX6 | This study |
| ire1Δ HOG1-HA | BY4741 ire1::kanMX4 HOG1-HA-HIS3MX6 | This study |
| hac1Δ HOG1-HA | BY4741 hac1::kanMX4 HOG1-HA-HIS3MX6 | This study |
| BY4741 HOG1-GFP | BY4741 HOG1-GFP-HIS3MX6 | This study |
| ire1Δ HOG1-GFP | BY4741 ire1::kanMX4 HOG1-GFP-HIS3MX6 | This study |
| hac1Δ HOG1-GFP | BY4741 hac1::kanMX4 HOG1-GFP-HIS3MX6 | This study |
| BY4741 SLT2-HA | BY4741 SLT2-HA-HIS3MX6 | This study |
| ire1Δ SLT2-HA | BY4741 ire1::kanMX4 SLT2-HA-HIS3MX6 | This study |
| hac1Δ SLT2-HA | BY4741 hac1::kanMX4 SLT2-HA-HIS3MX6 | This study |
| BY4741 SLT2-GFP | BY4741 SLT2-GFP-HIS3MX6 | This study |
| ire1Δ SLT2-GFP | BY4741 ire1::kanMX4 SLT2-GFP-HIS3MX6 | This study |
| hac1Δ SLT2-GFP | BY4741 hac1::kanMX4 SLT2-GFP-HIS3MX6 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Li, S.; Wang, J.; Liu, Y.; Deng, Y. Roles of High Osmolarity Glycerol and Cell Wall Integrity Pathways in Cadmium Toxicity in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2021, 22, 6169. https://doi.org/10.3390/ijms22126169
Zhao Y, Li S, Wang J, Liu Y, Deng Y. Roles of High Osmolarity Glycerol and Cell Wall Integrity Pathways in Cadmium Toxicity in Saccharomyces cerevisiae. International Journal of Molecular Sciences. 2021; 22(12):6169. https://doi.org/10.3390/ijms22126169
Chicago/Turabian StyleZhao, Yunying, Shiyun Li, Jing Wang, Yingli Liu, and Yu Deng. 2021. "Roles of High Osmolarity Glycerol and Cell Wall Integrity Pathways in Cadmium Toxicity in Saccharomyces cerevisiae" International Journal of Molecular Sciences 22, no. 12: 6169. https://doi.org/10.3390/ijms22126169